
Abstract
Age estimates were made on otoliths of 32 young-of-the-year (YOY) Barracuda, Sphyraena viridensis Cuvier (1829), captured in the south-western Mediterranean Sea. Increments were observed in Barracuda sagittae sectioned in a transverse plane and viewed with light microscopy. Increment counts were made for age estimation. Estimated ages ranged from 27 to 299 days of fish ranging in size from 15.9 to 38.9 cm total length (TL). The observed mean growth rate is 3.487 mm.day−1. Decreasing growth performance is evidenced according to age: 3.822 mm.day−1 between 159 and 302 mm (27–139 days), 1.596 mm.day−1 between 352 and 389 mm (172–299 days).
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Otoliths naturally log data that record and store information in their microstructure and chemistry at different spatio-temporal scales related to growth and habitat use by fish (Kalish 1989; Campana 1999; Yamashita et al. 2000; Berg et al. 2005). Because of the information they contain, otoliths are unique and the single most important biological structure in the teleost fish body (Berg et al. 2005). The discovery of daily increments has made the interpretation of otolith microstructure a major tool for investigating the early life history of fishes (Pannela 1971). The presence of daily increments has now been verified for a wide variety of species (Jones 1986).
Family Sphyraenidae (Barracudas) contains one genus Sphyraena which includes twenty-one species (Nelson 2006). They are pelagic to demersal fishes, single or in schools. Four Sphyraenidae species occur in the Mediterranean Sea, two of which, Sphyraena chrysotaenia (Klunzinqer, 1884) and S. flavicauda (Ruppell, 1838) are of Red Sea origin (Golani et al. 2006). Sphyraena viridensis (Cuvier, 1829) is an Atlanto-Mediterranean species found from Iceland to Morocco and Cape Verde, Madeira Island, the Canaries and the Azores (Ben-Tuvia 1986; Froese and Pauly 2014). However, its exact distribution and abundance is unknown (Kara and Bourehail 2003). Golani et al. (2006) noted that it occurs mainly in the southern Mediterranean. Milana et al. (2014) reported that S. viridensis is a native Mediterranean species, with an occurrence no longer rare and suggested as having a north ward distributional expansion. In many reports, S. sphyraena (Linnaeus, 1758) and S. viridensis are considered as the juvenile and adult of the same species S. sphyraena (Relini and Orsi-Relini 1997; Vacchi et al. 1999; Golani et al. 2006). Pastore (2009) describe a new species S. intermedia and believe that it is not hybrid between S. sphyraena and S. viridensis. Considering otolith shape, Bourehail et al. (2015) distinguish the two species S. sphyraena and S. viridensis.
In Mediterranean, Sphyraenidae, all species combined, have an average annual production of 1000 T in 2014 (FAO 2016). In western Mediterranean, S. viridensis is becoming more and more present in fisheries (Azzurro et al. 2011) and has become very common on the Algerian coasts since the early 1990s where it is highly appreciated by consumers.
Despite its ecological interest as indicator of Mediterranean warming (http://www.ciesm.org/marine/programs/tropicalization.htm) and its economic importance, information on the biology and dynamics of S. viridensis is extremely limited. Only Allam et al. (2004) studied its growth in Egyptian waters. Villegas-Hernandez et al. (2014) compared its reproduction with those of S. sphyraena in the north-west Mediterranean; Barreiros et al. (2002) analysed its diet in the Atlantic.
This study is the first attempt to estimate age and growth of young S. viridensis. In this purpose, we used the information recorded in the otoliths. Analysis of daily growth increments in otoliths from juvenile specimens can provide valuable information about the early growth stages. The ability to accurately determine the age and growth rate is an essential feature of population dynamics and stock assessment models for fisheries.
Materials and Methods
Study Area and Sampling
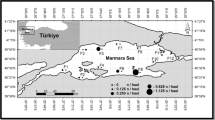
Sagittal otoliths used in this study were obtained from samples of 32 specimens, caught in the Gulf of Annaba in the south-western Mediterranean Sea (36°54’N, 7°45′E) (Fig. 1), where temperature ranged from 14 to 25.5 C° and salinity from 35.1 to 37.5 (Frehi et al. 2007). The juvenile Barracuda Sphyraena viridensis, ranging from 159 to 389 mm in total length (17.06 to 228.44 g) were collected between July and October 2009 from commercial trawlers and from the artisanal fishery.

Location of studied area in the South-Western Mediterranean Sea
Laboratory Procedures
Total weight (TW) in g, total length (TL) in mm, and date of capture were recorded for each fish. Sagittal otoliths (left and right) were removed, cleaned of adhering tissue, dried and stored in clean Eppendorf tube. The following measures of each otolith were taken with a calliper to the nearest mm: length (Lo), width (lo) and the thickness (To). Their weight (Wo) is committed to 0.001 g. From each pair, one otolith was randomly selected and mounted on a glass slide with thermoplastic Crystalbond resin. Each otolith was sectioned and ground with a series of abrasive papers of decreasing roughness from 12 to 9 to 3 mm and polished with 0.3 mm alumina paste on a polishing cloth until daily rings were discernible from the centre to the edge (Miller and Storck 1982; Secor et al. 1991; Jones 1992; Hayes 1995). The process was checked frequently under a light microscope adapted for video viewing to avoid over-polishing the centre. Counts of rings in each otolith were blind-read; the information about fish length and date of capture was withheld from the reader. All otoliths were read twice by two readers and final age estimates achieved when the differences, obtained from the two readings, were less than 5 days. A system microscope with objective lenses with nominal magnifications of 20×, 40× and 100× was used for the daily ring counts. The daily rings were counted from the nucleus to the dorso-posterior axis, however the readings were not always countable in a linear axis. Age was determined by counting the number of increments from the nucleus to the outer edge of the otolith.
The spawning season was determined by analysing the temporal evolution in the relative frequency of maturity stages and the gonado-somatic index GSI = (GW/TW) × 100. Based on juvenile frequency distribution and the GSI, the date of spawning was determined. Hatch date distributions were back-calculated by subtracting the estimated age (days) from the date of capture.
Data Analysis
The relationship between different otolith measures (length Lo, width lo, thickness To and weight Wo) and somatic parameters (TL, TW) was investigated by linear regression. Paired t-tests were used to determine if there were different daily age counts between left and right otoliths. No significant differences in increment counts were found between left and right otoliths (paired t-test, N = 32, P ˃ 0.05), so for further daily age estimations only one otolith was randomly selected. The precision of the daily ring counts was calculated using the average percentage error (APE) of Beamish and Fournier (1981) and coefficient of variation (CV) (Chang 1982; Campana 2001). An age–length key was generated to convert larger length samples from the population into ages. Observed daily growth rate was calculated by dividing the fish length by its age in days. Individual growth rate differences were researched by K-means analysis (MacQueen 1967). Theoretical daily growth rate (mm.d−1) was determined from the slopes of a linear regression based on the following equation:
where, TL is the total fish length (mm); a is the fish size at age 0; b is the somatic growth rate. Statistical analyses were performed with Minitab 18 software package.
Results
Otoliths of YOY. S. viridensis are lancelolated and thin. The inner face is convex and the outer face is flat. The dorsal margin is sinuate. The posterior margin is rounded ventrally. The sulcus is ostial and the cauda had a poorly defined tip. Length of YOY S. viridensis was strongly correlated with their otolith width (r2 = 0.872), length (r2 = 0.898), thickness (r2 = 0.742) and weight (r2 = 0.894) (Table 1). The Age-weight relationship is Wt (g) = 32.29exp0.007x (r2 = 0.833) (Fig. 2).

Age–weight relationship estimated for young of the year Sphyraena viridensis from Algerian eastern coasts
Sectioning and polishing of the sagittae enhanced the light microscopy images, particularly in the area near the core (Fig. 3). The primordium, thought to be the original point of growth, was clearly visible as a dark spot in the center of the core. In general, patterns of formation of micro-increments of YOY S. viridensis ˂ 40 cm TL were very distinctive, showing two micro-structural zones (I and II) (Fig. 3a.). The first one formed immediately after a diffuse area surrounding a central primordium and extended to ~20 days (Fig. 3b.). The increment width in Zone I, showing a sequential increase, a regular appearance and almost a complete absence of subdaily microincrements. Zone II is characterised by the presence of wider micro-increments (Fig. 3c.) where the formation pattern is more irregular, with the presence of growth discontinuities.

Photographs showing microstructural features of sagittae from a juvenile Sphyraena viridensis (250 mm TL, 72 days old) from Algerian eastern coasts
The estimated age obtained from the increment counts of the otoliths ranged between 27 d (15.9 mm TL) and 299 d (389 mm TL). The daily growth increments consisted of an accretion zone and discontinuous zone and were clearly and easily distinguished along a counting path from the core to the ventral edge. Precision of daily ring counts estimates was calculated as APE 1.5% and CV 1.86%. An APE and CV values which are below 10% comprise an acceptable ageing accuracy between ageing estimates.
Individual observed average daily growth rate varied between 1.301 ± 1.28 mm and 6.142 ± 1.28 mm (mean = 3.487 ± 1.28 mm). Two phases of different growth rate are identified: 3.822 mm.day−1 from 159 mm (27 d) to 302 mm (299 d) and 1.596 mm.day−1 from 352 (172 d) to 389 mm (299 d) (Fig. 4).

Age–length relationship estimated for young of the year Sphyraena viridensis from Algerian eastern coasts
Estimated spawning dates, based upon the otolith analysis, were from April 11 to August 03. The half of the sampled specimens (50%) spawned in July (Fig. 5).

Estimated date of spawning of S. viridensis from eastern Algeria, obtained from individual monthly length frequency, GSI and sampling time
Discussion
Otolith morphology of YOY. S. viridensis is not different from those of adults (Bourehail et al. 2015). Statistically significant relationships exist between otoliths measurement and fish length. Therefore, it is possible that otoliths morphometric measurements (Lo, lo, and Eo) could provide a good alternative way to age young S. viridensis.
Our observations of the sagittae clearly recognized fine increments resembling those found in otoliths from some adults (Bourehail et al. 2015). They were also similar to validated daily increments described for other species with similar growth pattern as the great barracuda S. barracuda (Kadison et al. 2010) and Swordfish Xiphias gladius (Megalofonou et al. 1987; Akyol and Ceyhan 2013).
A distinctive feature of this species was the occurrence of two microstructural zones in otoliths: a first zone where primary increments were more homogeneous and easy to identify, in contrast with a second zone where ring’s alignment and other perturbations predominated. Microstructural zones in sagittal otoliths of YOY have been reported in several teleost fish (Wellington and Victor 1989; Beldade et al. 2007; Sponaugle et al. 2010; Garcia-Seoane et al. 2015), particularly in demersal and reef-associated fish (Victor 1986; Wilson and McCormick 1997; Sponaugle et al. 2010). These features have been linked roughly with the larval stage, the transition from larval to juvenile stages or habitat transitions, and later juvenile stages where juveniles occur in nursery areas or deeper habitats (Jenkins and May 1994; Raventos and Macpherson 2001; Plaza et al. 2001; Mansur et al. 2014). Conversely, abrupt changes in otolith microstructure seem to be less common in sagittal otoliths of YOY of pelagic fish where habitat transition is less marked (Cerna and Plaza 2016). However, subdaily rings, double microincrements, discontinuities and other perturbations have been reported in some areas of sagittal otoliths in recent studies in other pelagic fish as the great barracuda S. barracuda (Kadison et al. 2010) and Sword fish Xiphias gladius (Akyol and Ceyhan 2013) similar to the microstructural features observed in the transition zone in this study.
The succession of spawning date (May – August; Bourehail et al. 2010) and the apparition of the smaller individuals in September 15th showed by fish length frequency distribution (Bourehail, unpub. data), support the daily deposition of rings on the otoliths of 0+S. viridensis. This is corroborated by the annual growth of this species who attains 445 mm at 1 year old (Bourehail et al. 2010).
Observed mean daily growth increment is 3.486 mm.day−1 during the ten first months of fish life. Two growth stages are recognized. During the first two months, the growth is better (3.822 mm.day−1) than the rest of the year (1.596 mm.day−1). The upward trend in the first stage suggests a period of accelerated growth, because larvae are able to feed independently. Importantly, during this period there is continuous development of morphological and sensory aspects of larvae, which increases their motility and ability to capture prey. The theoretical daily growth increment (0,67 mm.d−1) is very low compared to the observed value and is equal to the calculated value in the congeneric species S. barracuda inhabiting the North Atlantic Ocean (D’Alessandro et al. 2013). In swordfish Xiphias gladius, a species with the same annual growth pattern, Arata (1954) also estimated theoretical larvae daily growth at 0.60 mm. Larvae of Sphyraenidae are known to have some of the highest rates of growth and mortality of any marine fish studied (D’Alessandro et al. 2013). This fast growth is fueled by early digestive system development and consumption of larval fishes, which are of higher energy content and nutritional value than crustaceans (reviewed in Tanaka et al. 1996; Govoni et al. 2003).
The results of this study indicate that daily growth increments can be used to estimate the age of young-of-the year S.viridensis and help clarify some aspects of juvenile growth of this species. Further studies on otoliths from both smaller and larger individuals, can provide the information necessary to form a better understanding of the growth of juvenile Barracudas.
References
Akyol O, Ceyhan T (2013) Age and growth of swordfish (Xiphias gladius L.) in the Aegean Sea. Turk J Zool 37:59–64
Allam MS, Faltas SN, Ragheb E (2004) Age and growth of barracudas in the egyptian Mediterranean waters. Egypt J Aquat Res 30(2):281–289
Arata GF (1954) A contribution to the life history of the swordfish, Xiphias gladius Linnaeus, from the South Atlantic coast of the United States and the Gulf of Mexico. Bull Mar Sci Gulf Caribb 4(3):183–243
Azzurro E, Moschella P, Maynou F (2011) Tracking Signals of change in Mediterranean fish diversity based on local ecological knowledge. PLoS One 6(9):e24885. https://doi.org/10.1371/journal.pone.0024885
Barreiros JP, Santos RS, de Borba AE (2002) Food habits, schooling and predatory behaviour of the yellowmouth barracuda, Sphyraena viridensis (Perciformes: Sphyraenidae) in the Azores. Cybium 26(2):83–88
Beamish RJ, Fournier DA (1981) A method for comparing the precision of a set of age determination. Can J Fish Aquat Sci 38:982–983
Beldade R, Pedro T, Goncalves J (2007) Pelagic larval duration of 10 temperate cryptobenthic fishes. J Fish Biol 71:376–382. https://doi.org/10.1111/J.1095-8649.2007.01491.X
Ben-Tuvia A (1986) Sphyraenidae. In: Whitehead PJP, Bauchot M-L, Hureu J-C, Nielsen J, Tortonose E (eds) Fishes of the North-Eastern Atlantic and the Mediterranean, Vol. III. UNESCO, Paris, pp 1194–1196
Berg E, Sarvas TH, Harbitz A, Fevolden SE, Salberg AB (2005) Accuracy and precision in stock of north-East Arctic and Norwegian coastal cod by otoliths – comparing readings, image analyses and a genetic method. Mar Freshw Res 56:753–762. https://doi.org/10.1071/MF04172
Bourehail N, Lecomte-Finiger R, Kara MH (2010) Age, croissance et reproduction du barracuda Sphyraena viridensis (Sphyraenidae) des côtes de l’est Algérien. Rapport de Communication Internationale de la Mer Méditerranée CIESM 39:459
Bourehail N, Morat F, Lecompte-Finiger R, Kara MH (2015) Using otolith shape analysis to distinguish barracudas Sphyraena sphyraena and Sphyraena viridensis from the Algerian coast. Cybium 39(4):271–278
Campana SE (2001) Accuracy, precision and quality control in age determination, including a review of the use and abuse of age validation methods. J Fish Biol 59(2):197–242. https://doi.org/10.1111/J.10958649.2001.TB00127.X
Campana SE (1999) Chemistry and composition of fish otoliths: pathways, mechanisms and applications. Mar Ecol Prog Ser 188:263–297
Cerna F, Plaza G (2016) Daily growth patterns of juveniles and adults of the Peruvian anchovy (Engraulis ringens) in northern Chile. Mar Freshw Res 67:899–912. https://doi.org/10.1071/MF15032
Chang WYB (1982) A statistical method for evaluating the reproducibility of age determination. Can J Fish Aquat Sci 39(8):1208–1210
D’Alessandro EK, Sponaugle S, Cowen RK (2013) Selective mortality during the larval and juvenile stages of snappers (Lutjanidae) and great barracuda Sphyraena barracuda. Mar Ecol Prog Ser 474:227–242. https://doi.org/10.3354/meps10114
FAO (2016) FAO fisheries and aquaculture department. Universal Software for Fishery Statistical Time Series. FAO, Rome, Statistics and Information Service FishStatJ
Frehi H, Couté A, Mascarell G, Perrette-Gallet C, Ayada M, Kara MH (2007) Dinoflagellés toxiques et/ou responsables de blooms dans la baie d’Annba (Algérie). C R Biol 330:615–628
Garcia-Seoane E, Meneses ACI, Silva B (2015) Microstructure of the otoliths of the glacier lanternfish, Benthosema glaciale. Mar Freshw Res 66:70–77. https://doi.org/10.1071/MF13211
Golani DB, Ozturk B, Basusta N (2006) The fishes of eastern Mediterranean. Turkish marine Research Foundation, publication no. 24, Istanbul, Turkey
Govoni JJ, Laban EH, Hare JA (2003) The early life history of sword fish (Xiphias gladius) in the western North Atlantic. Fish Bull 101:778–789
Hayes JW (1995) Importance of stream versus early lake rearing for rainbow trout fry in Lake Alexandrina, South Island, New Zealand, determined from otolith daily growth patterns. N. Z. J. Mar Freshw Res 29:409–420
Jenkins GP, May HMA (1994) Variation in settlement and larval duration of King George whiting, Sillaginodes punctata (Sillaginidae), in swan bay, Victoria, Australia. Bull Mar Sci 54:281–296
Jones C (1986) Determining age of larval fish with the otolith increment technique. Fish Bull 84:91–104
Jones CM (1992) Development and application of the otolith increment technique. In: Stevenson DK, Campana SE (eds) Otolith microstructure examination and analysis, Ottawa: can spec Publ fish Aquat Sci, vol 117, pp 1–11
Kalish JM (1989) Otolith microchemistry: validation of the effects of physiology, age and environment on otolith composition. J Exp Mar Biol Ecol 132:151–178
Kara MH, Bourehail N (2003) Présence du Barracuda, Sphyraena viridensis (Sphyraenidés), sur les côtes de l’est algérien. Cybium 27:59–60
Kadison E, D’Alessandro EK, Davis GO, Hood PB (2010) Age, growth, and reproductive patterns of the great barracuda, Sphyraena barracuda, from waters of the Florida keys. Bull Mar Sci 86:773–784
MacQueen JB (1967) Some methods for classification and analysis of multivariate observations, proceedings of 5-th Berkeley symposium on mathematical statistics and probability. Berkeley, University of California Press 1:281–297
Mansur L, Plaza G, Landaeta MF, Ojeda FP (2014) Planktonic duration in fourteen species of intertidal rockyfishes from the southeastern Pacific Ocean. Mar Freshw Res 65:901–909. https://doi.org/10.1071/MF13064
Megalofonou P, De Metrio G, Lenti MC (1987) Età e dimensioni di prima maturità sessuale de1 pesce spada Xiphias gladiusl. Parte I Atti Sot It Sci Vet XLI:234–347
Milana V, Ciampoli M, Sola L (2014) mtDNA sequences of Sphyraena viridensis (Perciformes: Sphyraenidae) from Italy: insights into historical events and the phylogeny of the genus. Biol. J Linn Soc 113:635–641
Miller SJ, Storck T (1982) Daily growth rings in otoliths of young-of-the-year largemouth bass. Trans Am Fish Soc 111:527–530
Nelson RJS (2006) Fishes of the world. John Wiley & Sons. Inc., New York
Pannela G (1971) Fish otoliths, growth layers and periodical patterns. Science 173:1124–1127
Pastore MA (2009) Sphyraena intermedia sp. nov. (Piseces: Sphyraenidae): a potential new species of barracuda identified from the Central Mediterranean Sea. J Mar Biolog Assoc 89:1299–1303
Plaza G, Katayama S, Omori M (2001) Otolith microstructure of the black rockfish, Sebastes inermis. Mar Biol 139:797–805. https://doi.org/10.1007/S002270100620
Raventos N, Macpherson E (2001) Planktonic larval duration and settlement marks on otoliths of Mediterranean littoral fishes. Mar Biol 138:1115–1120. https://doi.org/10.1007/S002270000535
Relini M, Orsi-Relini L (1997) The two species of barracuda (Sphyraenidae) in the western Mediterranean. Cybium 21:216–222
Secor DH, Dean JM, Laban EH (1991) Manual for otolith removal and preparation for microstructural examination. Electric Power Research Institute and the belle. University of South Carolina, Columbia, South Carolina, W.Baruch Institute for Marine Biology and Coastal Research, 85 p
Sponaugle S, Walter KD, Denit KL, Llopiz JK, Cowen RK (2010) Variation in pelagic larval growth of Atlantic billfishes: the role of prey composition and selective mortality. Mar Biol 157:839–849
Tanaka M, Kaji T, Nakamura Y (1996) Developmental strategy of scombrid larvae: high growth potential related to food habits and precocious digestive system development. In: Watanabe Y, Yamashita Y, Oozeki Y (eds) Survival strategies in early life stages of marine resources. A. A. Balkema, Rotterdam, pp 125–139
Vacchi M, Boyer M, Bussotti S, Guidetti P, La Mesa G (1999) Some interesting species in the coastal fish fauna of Ustica Island (Mediterranean Sea). Cybium 23:323–331
Victor BC (1986) Duration of the planktonic larval stage of one hundred species of Pacific and Atlantic wrasses (family Labridae). Mar Biol 90:317–326. https://doi.org/10.1007/BF00428555
Villegas-Hernandez H, Munoz M, Lloret J (2014) Life-history traits of temperate and thermophilic barracudas (Teleostei: Sphyraenidae) in the context of sea warming in the Mediterranean Sea. J Fish Biol 84:1940–1957. https://doi.org/10.1111/jfb.12411
Wellington GM, Victor BC (1989) Planktonic larval duration of one hundred species of Pacific and Atlantic damsel fishes (Pomacentridae). Mar Biol 101:557–567. https://doi.org/10.1007/BF00541659
Wilson DT, McCormick MI (1997) Spatial and temporal settlement-marks in otoliths of tropical reef fishes. Mar Ecol Prog Ser 153:259–271. https://doi.org/10.3354/MEPS153259
Yamashita RA, Sellers JR, Anderson JB (2000) Identification and analysis of the myosin superfamily in Drosophila: a database approach. J Muscle Res Cell M 21(6):491–505. https://doi.org/10.1023/A:1026589626422
Reference to awebsite
CIESM Tropical Signals Program: monitoring macrodescriptor species of climate warming. http://www.ciesm.org/marine/programs/tropicalization.htm
Froese R, Pauly D (2014) Fish Base. http://www.fishbase.org (Accessed 25 June 2014)
Acknowledgements
The authors thank the Algerian Ministry for Higher education and scientific research (General directorate for scientific research and technology development, GDRSDT) which financially supported this study, within the framework of the National Funds of Research (NFR).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
On behalf of all authors, the corresponding author states that there is no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Bourehail, N., Kara, M.H. Young-of-the-Year Yellowmouth Barracuda Sphyraena viridensis (Cuvier, 1829) Growth in Eastern Algeria Based on Otolith Microstructure Analysis. Thalassas 37, 107–112 (2021). https://doi.org/10.1007/s41208-020-00256-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s41208-020-00256-2