
Abstract
A total of 23 whale sharks were identified over a 5 d period in the Arta Bay region of the Gulf of Tadjora, Djibouti. Most of the sharks aggregating in this area were small (<4 m TL) males. Individuals were identified using photographs of distinctive scars and spot and stripe patterns on the sides of the animals. Of these, 65% had scarring that was attributable to boat or propeller strikes. Most of the whale sharks we encountered were feeding on dense accumulations of plankton in shallow water just off (10–200 m) the shoreline. This food source may account for the aggregation of sharks in this area. One 3 m male shark was tagged with an ARGOS (Splash) satellite tag for 9 d. During this time the shark traversed to the shoreline on the opposite side of the Gulf (a distance of 14 km) and then returned to the Arta Bay area before retracing his path to the other shore. The shark spent most of the daylight hours at the surface, while at night dives were more frequent, deeper and for longer durations.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Whale sharks, Rhincodon typus, are found throughout the world’s tropical oceans (Compagno 2001) with the first scientific description being made from a specimen from the Indian Ocean (Smith 1829). Seasonal aggregations of these animals have been recorded at a number of coastal localities in the Indian Ocean such as off north-western Australia (Taylor 1989; Taylor 1996; Colman 1997; Meekan et al. 2006); India (Silas 1986; Hanfee 2001), the Maldives (Anderson and Ahmed 1993) and off Seychelles (Colman 1997; Newman et al. 1997; Rowat 1997). At many of these places, the predictable timing of these aggregations has allowed significant ecotourism industries to develop that support regional economies (Newman et al. 1997), while in others, aggregations have been targeted by fishermen (Hanfee 2001). In view of the long distance migrations of this species (Eckert and Stewart 2001; Eckert et al. 2002; Wilson et al. 2006) and their relative economic importance in some areas, it is valuable to characterise these aggregations and the movements of sharks within them.
Coastal aggregations of whale sharks are usually dominated by immature males of around 5–7 m in length (Heyman et al. 2001; Meekan et al. 2006). Spatial and/or temporal segregation of populations by size and gender is typical of sharks (Springer 1967; Sims et al. 2000), but raises questions as to the location of the remainder of the population, and the reasons for these aggregations. Coastal aggregations of small whale sharks in the Sea of Cortez appear to occur in response to seasonal aggregations of their planktonic food (Clarke and Nelson 1997), as is the case with larger members of the species (Wilson et al. 2001). There are, however, very few records of whale sharks from size at birth (around 58 cm to 64 cm), (Joung et al. 1996) to 3 m (Colman 1997). New-born whale sharks have occasionally been found in the guts of blue sharks, Prionace glauca, and blue marlin, Makaira mazara, suggesting that very small sharks may be pelagic (Colman 1997).
In 2004, anecdotal reports by a local conservation organisation suggested that a previously unknown aggregation site for whale sharks existed in coastal waters of the Gulf of Tadjoura, Djibouti, at the northern eastern margin of the Indian Ocean (Staszewski et al. in press). This aggregation appeared to be unusual in that it was composed of relatively small juvenile (3–4 m) sharks, similar to the aggregation reported in the Sea of Cortez, and the first reported in the Indian Ocean. Here, we describe the demography, behaviour and short-term (daily) movements and diving patterns of whale sharks in this aggregation in the Gulf of Tadjoura, Djibouti.
Methods
Study area
The Gulf of Tadjoura, (latitude 11° 40′N, longitude 43° 00′E) at the southern entrance to the Red Sea, is an inlet of the Indian Ocean caused by the fault line of the northerly end of the East African Rift Valley that transects Djibouti, Ethiopia and Kenya. The area is geologically and volcanically active as evidenced by the 1978 eruption of the Ardoukoba volcano. The seabed shelves steeply from the coast of the Gulf dropping to 100 m depth around 2 km offshore and to around 450 m depth in the centre of the Gulf of Tadjoura.
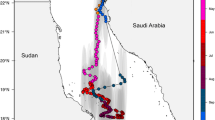
Anecdotal reports suggested that whale sharks occur during the months of October to February in the Arta Bay area, (latitude 11° 35′N, longitude 42° 49′E) on the southern coast of the western end of the Gulf of Tadjoura some 33 km from Djibouti city (Fig. 1).

Study area, Arta Bay on the Gulf of Tadjoura, Djibouti, north east Africa showing ARGOS satellite track of a 3 m male whale shark
Demographic study
From the 20th to the 24th of January 2006, whale sharks along a 3–5 km stretch of coastline centered on Arta Beach were found by haphazard surface searches from a small (6 m) boat. The sharks were found either by sighting their dorsal or tail fins breaking the surface or by watching for the swirling, vortex patterns on the water caused by the sharks’ suction feeding just below the surface. On sighting a shark, a recorder aboard the boat would note the time and location and the shark would be photographed using an underwater digital camera by a snorkeller. Where possible, images were recorded of the area behind the gills and forward of the dorsal fin for both sides of the shark, as well as of any obvious scarring. Details of each shark’s size, sex, behaviour and any other associated fauna were recorded, when possible, for each encounter. Size estimation was made by one of three trained researchers, each with at least 10 years of experience, by comparison of the shark against a nearby object of known length, such as the boat or snorkeller, or by the use of a graduated 1.5 m pole spear as a measuring aid. These by-eye estimates of length were usually within 25 cm of those made by measuring sharks with a tape measure.
Short-term shark movements were determined by evaluation of the daily sighting log and digital photography. Spot and stripe patterns on the dorsal surface were used to individually identify sharks from photographs (Meekan et al. 2006). Photographs were first separated into individual encounters, each encounter relating to a single shark. Obvious matches were then made by eye based on characteristics such as major scars and the presence of marker tags or tag remnants. The remainder were compared using spot and stripe patterns. The record of sightings based on identification photographs was analysed to establish the frequency of sighting and resighting of individuals.
Long distance movement and diving patterns were investigated by tagging one shark with a Splash satellite tag (Wildlife Computers, Seattle). The tag was attached through base of the dorsal fin of the shark on a 1 m stainless steel wire trace of 150 kg breaking strain, by a stainless steel anchor dart applied in-water by a modified spear-gun. The tag recorded position via ARGOS satellites when the shark was at the surface and depth and temperature at 2 s intervals. Summary histograms of these depth and temperature data sets were uploaded to the satellites when the shark was at the surface.
Histogram data was decoded using the ‘SATPAK2003’ software, provided by the tag manufacturer, and the tag’s location was plotted daily using the ‘STAT’ Satellite Tracking and Analysis Tool (Coyne and Godley 2005)Footnote 1. Distances between locations were calculated by STAT on high quality ARGOS location codes (1–3) only, to minimise errors and allow speeds to be accurately calculated.
Water temperatures were recorded using a ‘Hobo Tid-bit’ temperature logger (Onset Technologies). Where groups of sharks were found in a confined area the logger was lowered on a line to the sea-bed and then raised in 5 m increments to record a temperature depth profile. Weather and in-water visibility were recorded for each shark encounter.
Results
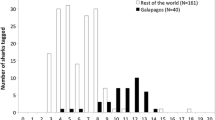
Nearly all the whale sharks were sighted along a 1.5 km stretch of coastline centred on Arta Beach and less than 500 m from shore. There appeared to be no consistent pattern in the distribution of sharks within this area. A total of 127 encounters with whale sharks were recorded during 1,044 min (17.4 h) of survey so that over the 5 d period of the study, an average of 7.3 sharks were encountered per hour. A total of 23 separate individuals were identified from photographs of which 16 were male, two female and for five the gender was undetermined (Table 1). The smallest shark was a 2.5 m male and the largest a 6 m female; the mean size was 4.47 m (SD = 0.84, n = 19). Size frequency distributions of these sharks were dominated by very small male sharks of 4 m or less (Fig. 2). The maximum number of resightings (based on photo-identification) of the same individual during our 5 d survey was 10, although most individuals were resighted on multiple occasions (mean resightings = 3.6 ± 2.85 SD, n = 23). The highest number of sightings made in any one session was 27 of a total of 10 identified individuals; the lowest number of sightings in any session was of three sharks that were each seen only once.

Size frequency distributions of individually identified whale sharks found at Arta Bay on the Gulf of Tadjoura, Djibouti
Of the 23 individual sharks identified during the 5 d study, 15 (65%) had scarring on fins or body. In many cases, the shape and size of the scars suggested that they were due to impacts with boats, propellers or possibly fishing gear.
Most of the whale sharks we encountered were actively feeding on dense aggregations of zooplankton that accumulated in shallow water along the steeply-shelving Arta Beach. Feeding behaviour recorded included both ram filter feeding and vertical feeding at the surface. The zooplankton on which the sharks were feeding were dense concentrations of chaetognaths, salps and mantis shrimp larvae. Temperature profiles at the 20 m depth contour off the beach where whale sharks were seen feeding varied from 27°C at the surface to 25.5°C at 20 m in the morning, while in the afternoon surface temperature increased to 29.3°C with a minimum of 27.1°C at 20 m.
Movement and diving patterns of a tagged shark
The 3 m male shark that was tagged with an ARGOS (Splash) satellite transmitter remained in the Arta area for the first few days after tagging then moved 14 km to the northwest coast of the Bay of Tadjoura (Fig. 1). It journeyed between these localities until on the 31st of January, 9 d after deployment, the tag was lost from the shark.
During deployment the maximum swimming speed of the shark was 2.99 km h−1 with an overall average of 0.42 km h−1. All speeds greater than 1 km h−1 were achieved over distances greater than 2 km, however, not all transits longer than 2 km were at speeds greater than 1 km h−1. The mean distance between surface locations logged by the satellite was 3.9 km (SD = 3.78, n = 23) with the maximum distance 14.42 km. The average daily distance covered was 9.01 km (SD = 5.77, n = 10).
The number and duration of dives below 15 m by the shark were found to be significantly less during the day than at night (Wilcoxon matched pairs test Z = 2.66, P = 0.007, n = 9, Mann–Whitney U test, n = 9 Table 2a) (Fig. 3).

Number and duration of dives made during the day compared to those made at night
The maximum depth of dives during the night was also significantly deeper than those made during the day (Mann–Whitney U test, n = 9 Table 2b) with the deepest dive from 175 m to 200 m. Time at depth data confirmed that 85% of the day (SD = 0.32, n = 8) was spent shallower than 15 m, while 52% (SD = 0.16, n = 9) of the night was spent at less than 15 m.
There was a significant difference between day and night in the time spent at different temperatures (Mann–Whitney U test, n = 9 Table 2c), with 60% of the day (SD = 0.12, n = 9) spent in water warmer than 29°C, while 87% of the night (SD = 0.3, n = 10) was spent in water colder than 29°C. The coldest temperature encountered by sharks during the day was 25–26°C while at night it was 22–23°C.
Discussion
The whale sharks participating in the aggregation at Arta Beach were relatively small (60% <4 m TL), as suggested by the ecotourism operators. The size of these sharks contrasts to those in aggregations in Belize (Heyman et al. 2001), Ningaloo in Western Australia (Meekan et al. 2006) and the Seychelles (Rowat and Gore in press) where sharks average closer to 6–7 m TL. One aggregation of small (3–4 m TL) juveniles has been reported in the north of the Sea of Cortez (Clarke and Nelson 1997), but our observations are the first reports of an aggregation of small whale sharks in the Indian Ocean. Similar to these other sites, most of the sharks gathering off the coast of Djibouti appeared to be male and few females were encountered.
While 23 individual sharks were identified during the study the total number may have been larger due to some photographs being unsuitable for identification, and some sharks eluding photography. Sharks were concentrated in a very small area of coastline, so that on some days, it was possible to see 4–5 sharks just offshore of a 300 m section of beach; on one occasion we counted 14 sharks within this area. Dense patches of zooplankton are thought to attract juvenile whale sharks to aggregate in the Sea of Cortez (Clark and Nelson 1997) and most of the sharks we encountered were feeding. Both vertical feeding and ram-surface feeding behaviours were observed in dense food patches.
The track of the tagged shark revealed that Arta Beach was not the only area frequented; over the 9 d that the tag was deployed on the shark, it transited 3 times between Arta and the opposite side of the Gulf suggesting that this may be another area worthy of investigation to determine if aggregations also occur on this northern shore. The relatively restricted area of movements recorded by this shark are similar to localised movement patterns from telemetry studies on whale sharks off Malaysia (Eckert et al. 2002). In that study, the four sharks tracked for periods of less than 12 d averaged 12.51 km per 24 h period (max = 18.92 km, min = 6.91 km), similar to the average distances recorded by our study.
In other localities, most whale sharks tend to spend more time in surface waters during the night than during the day (J.S. Gunn et al. unpublished; Heyman et al. 2001; Graham et al. 2005; Wilson et al. 2006); in contrast, the whale shark we tagged displayed the reverse pattern, spending more time at the surface during the day than at night. Bearing in mind that our study only gave a 9 d snapshot of the behaviour of a single shark, other studies have described similar patterns, notably J.S. Gunn et al. (unpublished) and Wilson et al. (2006), both of whom tracked whale sharks off Ningaloo Reef. In this Australian locality, whale sharks target surface swarms of a tropical krill, Pseudeuphausia latifrons (Taylor 1994; Wilson and Newbound 2001; Wilson et al. 2001; Wilson et al. 2006).
In the Gulf of Tadjoura, reverse diel vertical migration (DVM) of plankton might also contribute to this pattern. Studies in the temperate Irish Sea have shown that the large planktivorous basking shark, Cetorhinus maximus, also spends more time at the surface during the day in coastal waters (Sims et al. 2005). This occurs because calanoid copepods reverse their usual DVM pattern by ascending into shallow water during daylight hours to avoid invertebrate predators, specifically chaetognaths (Conway and Williams 1986). While chaetognaths have not been identified as prey for basking sharks, plankton samples taken from around those basking sharks that exhibited reverse DVM had very high concentrations of chaetognaths, as well as lower levels of calanoid copepods (Sims et al. 2005). In our study site off Arta Beach, chaetognaths formed dense swarms in surface waters and we witnessed whale sharks feeding on these swarms on many occasions.
Although the sharks were feeding in shallow water, the tagged shark also performed deep dives on transits between coasts and to the north of Arta in an area with steeply sloping bottom topography. Although the purpose of this diving is unknown, other studies have recorded whale sharks regularly descending to depths of over 1 km (Graham et al. 2005; Wilson et al. 2006). It has been suggested that in deep water, whale sharks are capable of feeding on epipelagic and mesopelagic prey such as euphausiids, myctophids and squids (Wilson et al. 2006) and that frequent deep dives may allow the sharks to detect prey concentrations (Josse et al. 1998) or olfactory trails leading to them (Carey and Scharold 1990, Gunn et al. 1999). Deep dives may also be important for thermoregulation (Carey and Scharold 1990).
The incidence of scarring of the whale sharks seen in the aggregation is of concern for the future of the species at this locality. Of the individuals we identified, 65% had large scars that could be attributed to boat or propeller strike. Although the coast of Arta is sparsely populated, the area is frequented on weekends by many boat-based sightseers and is also traversed by high-speed military boats. The susceptibility of whale sharks to boat strikes has long been recognised (Gudger 1937) and during this study one shark was cut by the propeller of a military vessel. Lack of management of vessel use and unrestricted access for large numbers of people could have detrimental affects on the aggregation as has been seen in Belize (Carne in press; Graham in press) and the Philippines (Pine et al. in press; Quiros in press). The relatively limited area in which the sharks were found make it both feasible, and an urgent priority, to implement some form of management plan to ensure the long term future of the whale sharks in this aggregation.
Notes
www.seaturtle.org
References
Anderson RC, Ahmed H (1993) Shark fisheries of the Maldives. Ministry of Fisheries and Agriculture, Maldives, and FAO, Rome 73 pp
Carey FG, Scharold JV (1990) Movements of blue sharks (Prionace glauca) in depth and course. Mar Biol 106:329–342
Carne L. Whale shark tourism at Gladden Spit marine reserve Belize, Central America. In: Irvine TR, Keesing JK (eds) The first international whale shark conference: Promoting international collaboration in whale shark conservation, science and management. CSIRO Marine and Atmospheric Research, Australia (in press)
Clarke E, Nelson DR (1997) Young whale sharks, Rhincodon typus, feeding on a copepod bloom near La Paz, Mexico. Environ Biol Fishes 50:63–73
Colman JG (1997) A review of the biology and ecology of the whale shark. J Fish Biol 51:1219–1234
Compagno LJV (2001) Sharks of the world. An annotated and illustrated catalogue of shark species known to date. Food and Agriculture Organization of the United Nations, Rome
Conway DVP, Williams R (1986) Seasonal population structure, vertical distribution and migration of the chaetognath Sagitta elegans in the Celtic sea. Mar Biol 93:377–387
Coyne MS, Godley BJ (2005) Satellite Tracking and Analysis Tool (STAT): an integrated system for archiving, analyzing and mapping animal tracking data. Mar Ecol Prog Ser 301:1–17
Eckert S, Dolar L, Kooyman G, Perin W, Rahman A (2002) Movements of whale sharks, (Rhyncodon typus), in South East Asian waters as determined by satellite telemetry. J Zool 257:111–115
Eckert S, Stewart B (2001) Telemetry and satellite tracking of whale sharks, Rhyncodon typus, in the Sea of Cortez, Mexico, and north Pacific Ocean. Environ Biol Fishes 60:299–308
Graham, R. Iterative planning and adaptive management of whale shark tourism in Belize: global implications of lessons learnt from 1998 and 2004. In: Irvine TR, Keesing JK (eds) The first international whale shark conference: Promoting international collaboration in whale shark conservation, science and management. CSIRO Marine and Atmospheric Research, Australia (in press)
Graham, RT, Roberts CM, Smart JCR (2005) Diving behaviour of whale sharks in relation to a predictable food pulse. Interface. J R Soc. doi:10.1098/rsif.2005.0082:1–8
Gudger EW (1937) A whale shark speared on the bow of a steamer in the Caribbean Sea. Copeia 1:60
Gunn JS, Stevens JD, Davis TLO, Norman BD (1999) Observations on the short term movements and behaviour of whale sharks (Rhincodon typus) at Ningaloo Reef, Western Australia. Mar Biol 135:553–559
Hanfee F (2001) Trade in Whale shark and its products in the coastal state of Gujarat, India. TRAFFIC India, New Delhi
Heyman W, Graham R, Kjerfve B, Johannes RE (2001) Whale sharks Rhincodon typus aggregate to feed on fish spawn in Belize. Mar Ecol Prog Ser 215:275–282
Josse E, Bach P, Dagorn L (1998) Simultaneous observations of tuna movements and their prey by sonic tracking and acoustic surveys. Hydrobiologia 372:61–69
Joung SJ, Chen CT, Clark E, Uchida S, Huang WYP (1996) The whale shark, Rhincodon typus, is a live-bearer: 300 embryos found in one ‘megamamma’ supreme. Environ Biol Fishes 46:219–223
Meekan MG, Bradshaw CJA, Press M, McLean C, Richards A, Quasnichka S, Taylor JA (2006) Population size and structure of whale sharks (Rhincodon typus) at Ningaloo Reef, Western Australia. Mar Ecol Prog Ser 319:275–285
Newman HE, Medcraft AJ, Colman JG (1997) Whaleshark Tagging and Ecotourism Elasmobrach Biodiversity, Conservation and Management, International Seminar and Workshop, IUCN, Sabah, Malaysia
Pine R, Alava MNR, Yaptinchay AA. Challenges and lessons learned in setting-up a community-based whale shark eco-tourism program: The case in Donsol, Philippines. In: Irvine TR, Keesing JK (eds) The first international whale shark conference: Promoting international collaboration in whale shark conservation, science and management. CSIRO Marine and Atmospheric Research, Australia (in press)
Quiros, AL. Tourist compliance to a code of conduct and the resulting effects on whale shark (Rhincodon typus) behavior in Donsol, Philippines. In: Irvine TR, Keesing JK (eds) Whale sharks: Science, conservation and management. Proceedings of the first international whale shark conference, 9–12 May 2005, Australia, Fisheries Research special issue (in press)
Rowat D (1997) Seychelles whale shark tagging project—pilot project report. Phelsuma, Nat Prot Trust Seychelles 5:77–80
Rowat D, Gore M. Regional scale horizontal and local scale vertical movements of whale sharks in the Indian Ocean off Seychelles. In: Irvine TR, Keesing JK (eds) Whale sharks: Science, conservation and management. Proceedings of the first International whale shark conference, 9–12 May 2005, Australia. Fisheries Research special issue (in press)
Silas EG (1986) The whale shark (Rhincodon typus Smith) in Indian coastal waters: is the species endangered or vulnerable? CMFRI, Marine Fisheries Information Service, Cochin, pp 1–19
Sims D, Southall E, Quayle V, Fox M (2000) Annual social behaviour of basking sharks associated with coastal front areas. Proceedings of the Royal Society, London B:1897–1904
Sims DW, Southall EJ, Tarling GA, Metcalfe JD (2005) Habitat-specific normal and reverse diel vertical migration in the plankton-feeding basking shark. J Anim Ecol 74:755
Smith A (1829) Contributions to the natural history of South Africa. Zool J 443–444
Springer S (1967) Social organisation of shark populations. In: Gilbert PW, Mathewson RF, Rall DP (eds) Sharks, skates and rays, Johns Hopkins Press, Baltimore, pp 149–174
Staszewski V, Conte E, Pardigon B, Takeda S, Jouannet D, Vely M. Whale shark (Rhincodon typus) in Djibouti (Horn of Africa): Conciliation of ecotourism development, conservation and development of a scientific programme. In: Irvine TR, Keesing JK (eds) The first international whale shark conference: Promoting international collaboration in whale shark conservation, science and management. CSIRO Marine and Atmospheric Research, Australia (in press)
Taylor G (1989) Whale sharks of Ningaloo Reef, Western Australia: a preliminary study. West Austr Nat 18:7–12
Taylor G (1994) Whale sharks. Angus, Robertson, Sydney, Australia
Taylor G (1996) Seasonal occurrence, distribution and movements of the whale shark, Rhincodon typus, at Ningaloo Reef, Western Australia. Mar Freshw Res 47:637–642
Wilson SG, Newbound DR (2001) Two whale shark faecal samples from Ningaloo Reef, Western Australia. Bull Mar Sci 68:367–362
Wilson SG, Polovina JJ, Stewart BS, Meekan MG (2006) Movements of whale sharks (Rhincodon typus) tagged at Ningaloo Reef, Western Australia. Mar Biol 148:1157–1166
Wilson SG, Taylor JG, Pearce AF (2001) The seasonal aggregation of whale sharks at Ningaloo Reef, Western Australia: currents, migrations and El Nino/Southern Oscillation. Environ Biol Fishes 61:1–11
Acknowledgements
We would like to thank the Marine Conservation Society Djibouti and their principle partner ‘Megaptera’ for the invitation to visit and participate in this research programme. The costs of the programme were sponsored largely by the Cooperation Service of the French Embassy in Djibouti (SCAC), by local tour operator Dolphin Excursions Sarl, and by the Fondation Nature & Decouvertes. Our thanks also to the Ministry of Environment, Djibouti, for their support and permission to undertake this research, to the Australian Institute of Marine Science and the Marine Conservation Society Seychelles for technical support. We would also acknowledge the valuable comments of two anonymous reviewers.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Rowat, D., Meekan, M.G., Engelhardt, U. et al. Aggregations of juvenile whale sharks (Rhincodon typus) in the Gulf of Tadjoura, Djibouti. Environ Biol Fish 80, 465–472 (2007). https://doi.org/10.1007/s10641-006-9148-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10641-006-9148-7