
Abstract
Background and Purpose
Liver fibrosis is characterized by accumulation of extracellular matrix. Our previous study found that osteopontin (OPN) increased in plasma of cirrhotic patients and indicative of cirrhosis staging. The present study was designed to investigate the expression of OPN in liver tissues and plasma of cirrhotic patients and further explore the role of OPN in human hepatic stellate cell (HSC) activation.
Methods
We used immunohistochemical staining and enzyme-linked immunosorbent assay to evaluate the expression level of OPN in liver tissues and plasma from cirrhotic patients, respectively. We produced lentivirus particles and infected target cell to manipulate OPN expression. Infection efficiency was determined by real-time RT-PCR and western blot. Cell proliferation was determined using CCK8 assay, and phenotypes of HSC activation were determined by real-time RT-PCR. OPN promoter activity was determined by dual luciferase reporter assay.
Results
We found that OPN expression in human cirrhotic liver tissues was upregulated compared to normal controls. In addition, its expression correlated with Child-Pugh classification, MELD score and the occurrence of complications. We further explored OPN level in patients’ plasma and showed that its level correlated with transforming growth factor-β1 (TGF-β1). In human HSC cell line LX-2, we found that change of OPN expression level could not only affect the proliferation of cells but also the TGF-β1 mediated HSC activation. Moreover, OPN was increased by TGF-β1 stimulation and regulated by TGF-β1 at transcription level.
Conclusions
OPN is upregulated in liver tissues and plasma of cirrhotic patients and promotes TGF-β1 mediated HSC activation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Continuous chronic liver injury by various factors leads to liver fibrosis via disruption of the balance between extracellular matrix (ECM) production and degradation. Advanced liver fibrosis is diagnosed as liver cirrhosis. Without effective therapy, it may ultimately develop into hepatic failure or even hepatocellular carcinoma. The most effective intervention in the treatment of advanced fibrosis is liver transplantation [1]. Hepatic stellate cell (HSC) has been regarded as the main producer of ECM [2]. HSC is located in the perisinusoidal space of Disse. In the normal liver, it is mainly responsible for storing vitamin A and regulating retinoic acid homeostasis [3]. Under injury stimuli, quiescent HSC transdifferentiates into its activated form, myofibroblast, which takes on some new attributes including increased proliferation, enhanced resistance to apoptosis and the ability of generating a large amount of collagen [2, 4]. In current understanding of molecular mechanism of liver fibrosis, multiple signaling pathways are interwoven to affect the fibrogenesis process. Specifically, the sustained state of HSC activation requires autocrine or paracrine stimuli of several cytokines such as PDGF, TGF-β1, leptin and VEGF [5]. Among them, TGF-β1 is considered the most potent fibrogenic cytokine in the liver [6].
TGF-β1 binds to a latency-associated peptide to maintain an inactivated state. Once activated, TGF-β1 recognizes its receptor and initiates the ECM production through recruiting Smad proteins. Moreover, TGF-β1 regulates removal of excess collagen after cessation of liver injury [7]. In many researches, manipulating TGF-β1 has been proven as an effective method to reverse liver fibrosis [7–10]. However, since the physiological function of TGF-β1 cannot be ignored, complete abrogation of TGF-β1 may be inappropriate [11].
Osteopontin (OPN), also called SPP1, is a secreted multifunctional glycoprotein which mainly is expressed in bone, kidney and epithelial tissues [12–15]. It is involved in various physiological and pathological processes such as bone metabolism, immune regulation, wound healing, cell survival and tumor progression [16–20]. As a secreted protein, OPN functions by acting on its corresponding receptors including integrins and CD44 [21]. An increasing body of evidence demonstrates that OPN participates in the process of fibrogenesis since OPN has been identified as one of the upregulating genes in culture-related rat HSC activation [22–24]. In our previous study, we found that OPN was a promising candidate for prediction biomarkers to evaluate the risk of cirrhosis and cancer in the long-term surveillance of patients with HBV infection [25]. Moreover, recent studies revealed a potential relationship between OPN and TGF-β1 either in osteogenic differentiation or in severe refractory asthma [26, 27]. Since TGF-β1 is capable of promoting the fibrogenesis process, it is very likely that besides existing explanations of the mechanism of its involvement in fibrogenesis, OPN participates in this process by interacting with the TGF-β1 signaling pathway.
Our present study was designed to investigate the expression of OPN in liver tissues and plasma of cirrhotic patients and further explore the role of OPN in HSC activation.
Materials and Methods
Specimen Collection
Cirrhotic liver tissues were collected from 61 patients who received surgical resection or biopsy examination at Xijing Hospital of the Fourth Military Medical University, Xi’an, China, from June 2009 to December 2010 with the patients’ consents. Normal tissues were histologically normal tissues obtained from patients who underwent hepatectomy for tumor or liver biopsy. Blood samples were obtained from 31 patients at the time of admission and anti-coagulated by ethylenediaminetetraacetic acid (EDTA). The diagnosis of cirrhosis was established on the basis of clinical, biochemical, imaging (ultrasonography and computertomography) and histological examinations. Plasma was collected by centrifuging the blood samples for 15 min at room temperature at 1,000×g within 30 min after collection and thereafter aliquoted and stored at −80 °C.
Immunohistochemistry
Tissues were fixed with formalin, and embedded in paraffin wax, before 4-μm-thick consecutive sections were made. All slides were dewaxed, rehydrated and immersed in 0.3 % hydrogen peroxide for 15 min to block endogenous peroxidase activity. Antigens were retrieved by heating the tissue sections at 100 °C for 15 min. Sections were cooled down and blocked with 10 % normal goat serum at room temperature for 15 min, before incubating with primary antibodies against OPN (Assay Designs, Ann Arbor, MI, USA) or TIMP1 (Santa Cruz Biotechnology, Delaware, CA, USA), respectively, at 4 °C overnight. The sections were then incubated with HRP-labeled goat anti-mouse secondary antibody (Zhongshan, Beijing, China). Results were visualized by using DAB kit (Zhongshan). All slides were examined systematically on an image analyzer system (Olympus BX51 microscope, Japan) and scored by two independent pathologists in a blinded manner. The intensity was graded by the ratio of positive cells per specimen and scored as follows: 0 = staining of <1 %; 1 = staining of 1–10 %; 2 = staining of 10–50 %; and 3 = staining of >50 % [28].
Cell Culture and Treatment
LX-2 cells were kindly provided by Prof. Scott L. Friedman (Mount Sinai School of Medicine, USA). LX-2 cells are immortalized human HSCs with a myofibroblast-like phenotype and used in study of HSC activation. They maintain key features of human HSCs and have the advantages of viability in serum-free media and high transfectability [29]. LX-2 cells and HEK 293T cells were cultured at 37 °C in a humidified atmosphere containing 5 % CO2 with Dulbecco’s modified Eagle’s medium containing 10 % fetal bovine serum and 100 IU/ml streptomycin/penicillin. Human recombinant transforming growth factor-β1 (TGF-β1) from PeproTech Asia (Rehovot, Israel) was diluted in citrate buffer (pH 3.0) and applied at a working concentration of 2 ng/ml as previously described [30].
Vector Construction, Lentivirus Production, and Target Cells Infection
An osteopontin (OPN) (NM_001040058) sequence containing EcoRI and HpaI sites was cloned into a FUW lentivirus vector. FUW–EGFP used as control was kindly provided by Dr. Jiuxu Bai (Shenyang General Hospital of PLA, China). Human OPN-specific siRNA pairs containing EcoRI and AgeI sites were synthesized, annealed and cloned into pLKO.1 lentivirus vector. A pLKO.1 vector containing scramble siRNA sequence used as control was purchased from Addgene. Primers or oligos used for constructing vectors are listed in Table S3. The whole inserts were sequenced for confirmation.
Lentivirus was produced in HEK 293T cells using the FUW (pLKO.1)/psPax2/pMD2.G system (4, 3 and 1 μg respectively for each 6 cm dish). Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) was used to perform transfection. Forty-eight hours after transfection, 5 ml culture media containing lentivirus particles was harvested and centrifuged at 4 °C, 1,250 rpm for 5 min for pelleting HEK293T cells. Thereafter, 1 ml culture media was added to 70 % confluent cells in each well of 6-well plate to infect cells. After incubation for 24 h, the media containing lentivirus was replaced by fresh media. A selective pressure of 3 μg/ml puromycin was used to select pLKO.1 infected cells.
Immunoblotting
Protein samples were prepared from cultured cells treated in RIPA lysis buffer (25 mM Tris–HCl, pH 7.5, 150 mM NaCl, 1 mM EDTA, 1 % Triton X-100) supplemented with protease inhibitor cocktail tablet (Roche Applied Science, Mannheim Germany) and subjected to 12 % SDS–polyacrylamide gel and transferred to a nitrocellulose membrane. The membrane was blocked with Tris-buffered saline containing 0.1 % Tween-20 and 5 % nonfat milk powder for 1 h at room temperature, before incubating with anti-OPN (Assay Designs) or anti-actin (St. Louis, MO, USA) at 4 °C overnight. The membrane was then incubated with goat anti-mouse secondary antibody (Boster, Wuhan, Hubei, China) for 1 h at room temperature. Protein bands were visualized by enhanced chemiluminescence (BIO-RAD, Hercule, CA, USA) on the BIO-RAD ChemiDoc XRS Imaging system.
Enzyme-Linked Immunosorbent Assay
TGFβ-1 or OPN was detected in plasma collected from cirrhotic patients or conditioned medium by enzyme-linked immunosorbent assay (ELISA, R&D Systems, Minneapolis, MN, USA) following manufacturer’s protocol. For TGF-β1 detection, samples were activated by 1 M HCl for 10 min at room temperature and neutralized by 1 M NaOH before test. The optical density of each well was determined at 450 nm within 30 min on an automatic ELISA reader. As for the collection of conditioned medium from cultured cells, 2 ml medium from a well of a 6-well plate (90 % confluent) was collected and subjected to ELISA assay.
RNA Extraction and Real-Time RT-PCR
Total RNA of cells was extracted from cultured cells using Trizol reagent (Invitrogen) according to manufacturer’s protocol. The quantity of RNA was determined on BIO-RAD SmartSpec™ Plus spectrophotometer and a total of 500 ng RNA of each sample was subjected to reverse transcription by using TaKaRa PrimeScript RT reagent kit (TaKaRa, Shiga, Japan). The cDNA quantity was evaluated on a Roche LightCycler 480 PCR machine by monitoring SYBR Green (TaKaRa) signal. The PCR reaction was cycled 40 times and the condition for each cycle was 95 °C for 5 s and 60 °C for 30 s. The data were normalized to 18S expression and further normalized to the control group. The primers used for PCR amplification are listed in Table S4.
Proliferation Assay
LX-2 cells were seeded into a 96-well plate at a density of 1,000 cells per well and deprived of serum for 24 h before experiment. For every well containing 200 μl culture medium, 20 μl of CCK8 solution (Beyotime Institute of Biotechnology, Jiangsu, China) was added. After 1 h of incubation at 37 °C, optical density value of each well was read at 450 nm. A group of wells without seeding cells served as a blank control group. Each group included five repeated wells. The experiment was carried out for 7 days and the proliferation of cells was observed at a certain time point every day.
Dual Luciferase Reporter Assay
pGL3–OPN–Luc plasmid containing OPN promoter sequence (−1,000 to +99) was provided by Dr. Jiuxu Bai. LX-2 cells in 24-well plates were transfected with 100 ng of pGL3–OPN–Luc/pGL3–Luc and 50 ng of normalization plasmid pRL–TK Renilla, with or without 2 ng TGF-β1. The dual luciferase reporter assay was performed 2 days after transfection using a luciferase assay system (Promega, Madison, WI, USA). Activities were normalized to pRL–TK activity and further normalized to pGL3-Luc group.
Statistical Analysis
For immunohistochemistry study, Pearson χ2 test and one-way ANOVA were used to determine the significance of the difference; kappa test was adopted to compare expression levels of OPN and TIMP1. For ELISA study, Pearson's correlation test was used to analyze the correlation between OPN and TGF-β1. For the rest of the numerical data, all experiments were repeated three times and the results were expressed as the mean ± standard deviation and analyzed by Student’s t test or ANOVA as appropriate. SPSS 17.0 software (SPSS Inc., Chicago, IL, USA) was used for statistical analysis. The level of significance was set at P < 0.05.
Results
Detection of OPN Expression in Cirrhotic Liver Tissues and Plasma
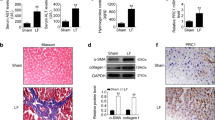
To evaluate OPN expression level in cirrhotic tissues and explore the relationship between OPN and the clinical features of cirrhotic patients, we detected the expression of OPN in human cirrhotic liver tissues and normal liver tissues by immunohistochemical staining. The results revealed that OPN was localized mainly in cytoplasm of hepatocellular cells and cholangiocytes (Fig. 1a). OPN level increased in cirrhotic tissues compared to normal control. Specifically, in 61 cases of cirrhotic tissue, 46 cases displayed positive OPN staining. As for the normal control group, 9 of 23 cases displayed positive staining (Table S1). In addition, OPN level was increased with the elevation of Child–Pugh classification and MELD scoring level (Fig. 1b) (P = 0.000, P = 0.001). We also found that patients with complications (ascites, hepatic encephalopathy, hepatic nephropathy, upper GI hemorrhage) tended to display a higher OPN expression than patients without complications (P = 0.005, Table 1). Furthermore, we compared OPN expression level with TIMP1, another profibrogenic marker. We found that OPN and TIMP1 tended to show moderate agreement in expression levels in cirrhotic liver tissues (Fig. S1, kappa = 0.518, P = 0.000). There was no difference between the expression of OPN and patients’ age, gender and etiologies.

Expression of osteopontin (OPN) in liver tissues and plasma from cirrhotic patients. a Immunohistochemical staining of OPN in normal and cirrhotic liver tissues (original magnification ×200). a Normal liver tissues showing negative OPN staining. b Cirrhotic liver tissues showing negative OPN staining. Cirrhotic liver tissues manifesting weak staining (c), moderate straining (d) and strong staining (e) in the cytoplasm of hepatocytes. f An area of fibrotic septa where cholangiocytes staining (indicated by black arrows) could be observed. b The intensity of immunohistochemical staining of OPN in cirrhotic liver tissues correlated with MELD score. Box plots represent median, quartiles and extremes (one-way ANOVA analysis, P = 0.003). c A regression line showing that OPN expression level in cirrhotic patients’ plasma correlated with TGF-β1 level (Pearson r 0.621, P = 0.000)
Since TGF-β1 is considered as a main mediator of liver fibrosis, and both TGF-β1 and OPN are secreted proteins, we performed ELISA to analyze the correlation between plasma OPN and TGF-β1 in 31 cases of patients with cirrhosis. The common characteristics of patients included in the test are listed in Table S2. We found that OPN plasma level (74.35 ± 25.47 ng/ml) tended to correlate with TGF-β1 plasma level (3.29 ± 1.43 ng/ml, Fig. 1c). Pearson correlation analysis showed a correlation coefficient of 0.621 (P = 0.000).
Lentivirus Mediated Transduction Resulted in OPN Overexpression or Knockdown in LX-2 cells
Since HSC has been widely acknowledged as the primary effector cell acting as the main origin of ECM, we chose a human HSC cell line LX-2 as the model to study the role of OPN in HSC activation [29]. OPN overexpression and knockdown were introduced into LX-2 cells by lentivirus. FUW-OPN and FUW-EGFP were transducted in parallel into LX-2 and the transduction efficiency was monitored by observing the EGFP expression in the cells. Over 90 % of the cells were labeled with EGFP 4 days after virus infection (Fig. 2a). After detecting OPN level by western blot and real-time RT-PCR, we found both OPN mRNA level and protein level were elevated by lentivirus infection (Fig. 2b, c). PLKO.1-siOPN but not pLKO.1-scram induced an 80 % decrease of OPN mRNA level in LX-2 after puromycin selection (Fig. 2d). The result was confirmed by western blot (Fig. 2e). To further corroborate the overexpression or knockdown effect of OPN, we detected OPN level in the conditioned medium and the results are shown in Table S5.

Overexpression or knockdown of osteopontin (OPN) in LX-2 cells by lentivirus infection. a The infection efficiency of lentivirus was monitored by the expression of EGFP. Overall, an infection efficiency of over 90 % was obtained. A representative image was shown. b, c The overexpression of OPN in LX-2 cells was confirmed by real-time RT-PCR and western blot. d, e The knockdown of OPN in LX-2 cells was confirmed by real-time RT-PCR and western blot (n = 3, P value was indicated vs. corresponding control)
OPN Promoted TGF-β1 Mediated HSC Activation in LX-2 cells
Proliferation is one of the key behavior changes of HSCs in sustained activation state [31], so we investigated the effect of OPN on proliferation of LX-2 cells. In a continuous 7-day observation, we found that FUW-EGFP cells showed similar growth rate to Neg-con cells, whereas FUW-OPN cells showed much faster growth rate compared to FUW-EGFP cells and Neg-con cells (Fig. 3a). Meanwhile, knockdown of OPN by pLKO.1-siOPN significantly inhibited LX-2 growth rate compared to pLKO.1-scram and Neg-con cells (Fig. 3b). To further evaluate the activation of LX-2 cells, we detected the mRNA level of α-smooth muscle actin (α-SMA), collagen I and tissue inhibitor of metalloproteinase (TIMP1) by real-time RT-PCR. α-SMA is a marker of HSC activation [2]. Collagen I represents ECM deposition and TIMP1 prevents the degradation of ECM via inhibiting metalloproteinase. We found that OPN overexpression enhanced the basal as well as the TGF-β1 induced level of HSC activation while OPN knockdown resulted in the reverse effect (Fig. 3c, d).

The effect of osteopontin (OPN) alteration on LX-2 activation phenotypes. a, b The growth curves of LX-2 showed that upregulation of OPN promoted proliferation of LX-2 (a) while downregulation of OPN inhibited proliferation (b). c, d α-SMA, Collagen I and TIMP1 mRNA level were determined to represent the activation level of LX-2. Upregulation of OPN promoted LX-2 activation (c) while downregulation of OPN caused the reverse effect (d) (n = 3, *P = 0.000 vs. corresponding control)
TGF-β1 Engages in the Transcriptional Regulation of OPN
To further investigate the relationship between OPN and TGF-β1, we stimulated LX-2 by 2 ng/ml of TGF-β1 for 24 h and observed an increase in the protein level of OPN (Fig. 4a). To examine whether the TGF-β1 induction of OPN was due to transcriptional regulation, we performed dual luciferase reporter assay in LX-2 and found that TGF-β1 could promote the promoter activity of OPN (Fig. 4b). Taken together, OPN is a transcriptional target of TGF-β1.

Osteopontin (OPN) is transcriptionally regulated by TGF-β1. a OPN protein level was increased by TGF-β1 stimulation (2 ng/ml for 24 h). b TGF-β1 increased the promoter activity of OPN (n = 3, P value was indicated vs. corresponding control)
Discussion
Liver fibrosis has been characterized by excess deposition of ECM, which is mainly caused by chronic HBV/HCV infection, alcohol abuse, immune injury and nonalcoholic steatohepatitis (NASH) [1]. Although liver biopsy is taken as the gold-standard diagnostic method for liver fibrosis, it still has some defects such as causing pain, bringing in complications and producing incorrect diagnosis when small biopsies are analyzed [32, 33]. Thus, assessment of the disease development based on non-invasive clinical examinations is promising and may replace the need for biopsy in the near future [34, 35].
To date, the only effective therapeutic method for advanced liver fibrosis is liver transplantation. In fact, much effort has contributed to reversing fibrosis, and many promising therapeutic methods have been developed in experimental models. For example, a recent study adopted low molecular weight heparin-pluronic nanogels to impede liver fibrosis in a rat model via blocking the TGF-β/Smad signaling pathway [9]. This work represents not only the typical method to reverse fibrosis via inhibiting classic profibrogenic pathway but also the attempt of introducing new materials to meet the goal. As anti-fibrotic studies go further, more novel modulators in fibrosis are revealed and provide potential therapeutic targets. For example, a study found that neuropilin-1, an axonal guidance molecule, could promote cirrhosis by enhancing PDGF/TGF-β signaling in HSC, and its neutralizing antibody could block liver fibrosis in a rat model of liver injury [36]. However, applying these findings in clinical practice still faces obstacles due to the much more complicated situation in humans [37].
In the present study, we presented the expression analysis of OPN in cirrhotic liver tissues as well as in plasma with the purpose of revealing the relationship between OPN expression and clinical characteristics. We further investigated the profibrogenic role of OPN in the human HSC cell line LX-2 and its involvement in TGF-β1 mediated HSC activation.
Our previous study showed a correlation between plasma OPN and HBV-cirrhosis, however, due to lack of tissue specimens, more accurate correlation could not be drawn. In the present study, we verified our previous findings in liver tissues and found that OPN expression increased in cirrhotic liver tissues compared to normal liver tissues. We also found OPN expression level positively correlated with Child–Pugh classification and MELD scale. Both scoring systems have been used to assess the severity of chronic liver diseases, evaluate the prognosis, and determine the necessity of performing liver transplantation [38]. The result that OPN expression positively correlates with the scoring level and the occurrence of complications suggests that OPN could be used as a biomarker to predict the severity of cirrhosis. In line with our findings, previous studies demonstrated that plasma level of OPN could serve as a useful indicator for evaluating the severity of cirrhosis and the risk of hepatocellular carcinoma [25, 39, 40]. As mentioned above, although liver biopsy is still considered as gold standard to identify liver histological stages, there is a need for developing non-invasive diagnostic methods. Imaging methods such as FibroTest, FibroMAX and transient elastography facilitate the clinical management of patients with cirrhosis [41]. And a recent study pointed out that genetic markers in combination with imaging techniques may be the better non-invasive diagnostic method in the future [34]. Since OPN acts as a secreted protein which is detectable in blood, it would be beneficial to adopt OPN as a marker contributing to the non-invasive diagnostic methods.
We then moved to evaluate plasma OPN level and investigate its relationship with one of the major fibrosis promoters TGF-β1 in the hope of proposing a possible explanation for OPN increase in cirrhosis. The results of plasma analysis and in vitro experiments indicated that OPN increase could partly be a result of TGF-β1 stimulation. In fact, there have been reports about several OPN relating pathways involved in fibrosis. OPN was proven in a NASH model as a Hedgehog-regulated profibrogenic factor [23]. Moreover, it could be induced by oxidant stress and its contribution to fibrogenesis response is partly realized by activating the PI3K–pAkt–NFκB pathway [24]. Our study revealed a possible transcriptional regulation of TGFβ-1 on OPN, which supplements the complexity of OPN related profibrogenesis network. However, the specific transcription target and the transcription factor engaging in the TGF-β1 pathway are not yet clarified in the present study. Our preliminary findings imply that the TGF-β1 promotion of fibrosis takes effect partly by mediating OPN.
It has been reported elsewhere that OPN could affect fibrosis by increasing collagen deposition, which contributes most to the ECM accumulation [42]. It was corroborated in our findings that OPN could promote ECM deposition at both basal and TGF-β1 induced levels. The increase of collagen I caused by OPN overexpression was probably a result of MMP13 decrease as reported before [24, 43]. Moreover, we proved that OPN could increase the proliferation of LX-2 cells. Several methods targeting HSC such as inducing the apoptosis of HSC and inhibiting the HSC activation related signaling pathways have been proposed as resolutions of fibrosis [44–46]. Our results indicate that OPN downregulation could significantly inhibit the activation phenotypes of HSC and support the idea of using OPN as a therapeutic target in reversing fibrosis.
In conclusion, our results suggest that OPN is a relevant biomarker indicating the severity and progress of liver fibrosis. Although our mechanism study was limited to in vitro cell lines, it offered substantial evidence that OPN could promote the activation of HSC and contribute to liver fibrosis in a TGF-β1 mediated way.
References
Bataller R, Brenner DA. Liver fibrosis. J Clin Invest. 2005;115:209–218.
Moreira RK. Hepatic stellate cells and liver fibrosis. Arch Pathol Lab Med. 2007;131:1728–1734.
Carpino G, Franchitto A, Morini S, et al. Activated hepatic stellate cells in liver cirrhosis. A morphologic and morphometrical study. Ital J Anat Embryol. 2004;109:225–238.
Kawada N. Human hepatic stellate cells are resistant to apoptosis: implications for human fibrogenic liver disease. Gut. 2006;55:1073–1074.
Lee UE, Friedman SL. Mechanisms of hepatic fibrogenesis. Best Pract Res Clin Gastroenterol. 2011;25:195–206.
Gressner AM, Weiskirchen R. Modern pathogenetic concepts of liver fibrosis suggest stellate cells and TGF-beta as major players and therapeutic targets. J Cell Mol Med. 2006;10:76–99.
Ueberham E, Low R, Ueberham U, et al. Conditional tetracycline-regulated expression of TGF-beta1 in liver of transgenic mice leads to reversible intermediary fibrosis. Hepatology. 2003;37:1067–1078.
Cheng K, Yang N, Mahato RI. TGF-beta1 gene silencing for treating liver fibrosis. Mol Pharm. 2009;6:772–779.
Lee JH, Lee H, Joung YK, et al. The use of low molecular weight heparin-pluronic nanogels to impede liver fibrosis by inhibition the TGF-beta/Smad signaling pathway. Biomaterials. 2011;32:1438–1445.
Du SS, Qiang M, Zeng ZC, et al. Radiation-induced liver fibrosis is mitigated by gene therapy inhibiting transforming growth factor-beta signaling in the rat. Int J Radiat Oncol Biol Phys. 2010;78:1513–1523.
Breitkopf K, Godoy P, Ciuclan L, Singer MV, Dooley S. TGF-beta/Smad signaling in the injured liver. Z Gastroenterol. 2006;44:57–66.
Nagao M, Feinstein TN, Ezura Y, et al. Sympathetic control of bone mass regulated by osteopontin. Proc Natl Acad Sci USA. 2011;108:17767–17772.
Schordan S, Schordan E, Endlich K, Endlich N. AlphaV-integrins mediate the mechanoprotective action of osteopontin in podocytes. Am J Physiol Renal Physiol. 2011;300:F119–F132.
Maki M, Hirota S, Kaneko Y, Morohoshi T. Expression of osteopontin messenger RNA by macrophages in ovarian serous papillary cystadenocarcinoma: a possible association with calcification of psammoma bodies. Pathol Int. 2000;50:531–535.
Apparao KB, Murray MJ, Fritz MA, et al. Osteopontin and its receptor alphavbeta(3) integrin are coexpressed in the human endometrium during the menstrual cycle but regulated differentially. J Clin Endocrinol Metab. 2001;86:4991–5000.
Giachelli CM, Steitz S. Osteopontin: a versatile regulator of inflammation and biomineralization. Matrix Biol. 2000;19:615–622.
Mazzali M, Kipari T, Ophascharoensuk V, et al. Osteopontin—a molecule for all seasons. QJM. 2002;95:3–13.
O’Regan A. The role of osteopontin in lung disease. Cytokine Growth Factor Rev. 2003;14:479–488.
Johnson GA, Burghardt RC, Bazer FW, Spencer TE. Osteopontin: roles in implantation and placentation. Biol Reprod. 2003;69:1458–1471.
Philip S, Bulbule A, Kundu GC. Osteopontin stimulates tumor growth and activation of promatrix metalloproteinase-2 through nuclear factor-kappa B-mediated induction of membrane type 1 matrix metalloproteinase in murine melanoma cells. J Biol Chem. 2001;276:44926–44935.
Denhardt DT, Noda M, O’Regan AW, Pavlin D, Berman JS. Osteopontin as a means to cope with environmental insults: regulation of inflammation, tissue remodeling, and cell survival. J Clin Invest. 2001;107:1055–1061.
Lee SH, Seo GS, Park YN, Yoo TM, Sohn DH. Effects and regulation of osteopontin in rat hepatic stellate cells. Biochem Pharmacol. 2004;68:2367–2378.
Syn WK, Choi SS, Liaskou E, et al. Osteopontin is induced by hedgehog pathway activation and promotes fibrosis progression in nonalcoholic steatohepatitis. Hepatology. 2011;53:106–115.
Urtasun R, Lopategi A, George J, et al. Osteopontin, an oxidant stress-sensitive cytokine, up-regulates collagen-i via integrin alpha(V) beta(3) engagement and PI3K–pAkt–NFkappaB signaling. Hepatology. 2012;55:594–608.
Zhao L, Li T, Wang Y, et al. Elevated plasma osteopontin level is predictive of cirrhosis in patients with hepatitis B infection. Int J Clin Pract. 2008;62:1056–1062.
Delimpoura V, Bakakos P, Tseliou E, et al. Increased levels of osteopontin in sputum supernatant in severe refractory asthma. Thorax. 2010;65:782–786.
Zhao L, Jiang S, Hantash BM. Transforming growth factor beta1 induces osteogenic differentiation of murine bone marrow stromal cells. Tiss Eng Part A. 2010;16:725–733.
Wang WH, Jiang CL, Yan W, et al. FOXP3 expression and clinical characteristics of hepatocellular carcinoma. World J Gastroenterol. 2010;16:5502–5509.
Xu L, Hui AY, Albanis E, et al. Human hepatic stellate cell lines, LX-1 and LX-2: new tools for analysis of hepatic fibrosis. Gut. 2005;54:142–151.
Li YL, Wu J, Wei D, et al. Newcastle disease virus represses the activation of human hepatic stellate cells and reverses the development of hepatic fibrosis in mice. Liver Int. 2009;29:593–602.
Paradis V, Dargere D, Bonvoust F, et al. Effects and regulation of connective tissue growth factor on hepatic stellate cells. Lab Invest. 2002;82:767–774.
Thampanitchawong P, Piratvisuth T. Liver biopsy: complications and risk factors. World J Gastroenterol. 1999;5:301–304.
Regev A, Berho M, Jeffers LJ, et al. Sampling error and intraobserver variation in liver biopsy in patients with chronic HCV infection. Am J Gastroenterol. 2002;97:2614–2618.
Ahmad W, Ijaz B, Gull S, et al. A brief review on molecular, genetic and imaging techniques for HCV fibrosis evaluation. Virol J. 2011;8:53.
Denzer UW, Luth S. Non-invasive diagnosis and monitoring of liver fibrosis and cirrhosis. Best Pract Res Clin Gastroenterol. 2009;23:453–460.
Cao S, Yaqoob U, Das A, et al. Neuropilin-1 promotes cirrhosis of the rodent and human liver by enhancing PDGF/TGF-beta signaling in hepatic stellate cells. J Clin Invest. 2010;120:2379–2394.
Wynn TA. Cellular and molecular mechanisms of fibrosis. J Pathol. 2008;214:199–210.
Kamath PS, Kim WR. The model for end-stage liver disease (MELD). Hepatology. 2007;45:797–805.
Huang W, Zhu G, Huang M, et al. Plasma osteopontin concentration correlates with the severity of hepatic fibrosis and inflammation in HCV-infected subjects. Clin Chim Acta. 2010;411:675–678.
El-Din BS, Elwan NM, Suliman GA, El-Shourbagy SH. Clinical significance of plasma osteopontin level in Egyptian patients with hepatitis C virus-related hepatocellular carcinoma. Arch Med Res. 2010;41:541–547.
Stevenson M, Lloyd-Jones M, Morgan M, Wong R. Non-invasive diagnostic assessment tools for the detection of liver fibrosis in patients with suspected alcohol-related liver disease: a systematic review and economic evaluation. Health Technol Assess. 2012;16:1–174.
Hunter C, Bond J, Kuo PC, Selim MA, Levinson H. The role of osteopontin and osteopontin aptamer (OPN-R3) in fibroblast activity. J Surg Res. 2011.
Schaefer B, Rivas-Estilla AM, Meraz-Cruz N, et al. Reciprocal modulation of matrix metalloproteinase-13 and type I collagen genes in rat hepatic stellate cells. Am J Pathol. 2003;162:1771–1780.
Kisseleva T, Brenner DA. Role of hepatic stellate cells in fibrogenesis and the reversal of fibrosis. J Gastroenterol Hepatol. 2007;22:S73–S78.
Borkham-Kamphorst E, Herrmann J, Stoll D, et al. Dominant-negative soluble PDGF-beta receptor inhibits hepatic stellate cell activation and attenuates liver fibrosis. Lab Invest. 2004;84:766–777.
Di Sario A, Bendia E, Svegliati BG, et al. Effect of pirfenidone on rat hepatic stellate cell proliferation and collagen production. J Hepatol. 2002;37:584–591.
Acknowledgments
This work was supported by National Natural Sciences Foundation of China, No. 81070350. We thank Technician Zheng Chen in our lab and Dr. Jiuxu Bai from Shenyang General Hospital of PLA for technical assistance. We thank Professor Gaoliang Ouyang (School of Life Sciences, Xiamen University) for providing the OPN plasmid. We also thank Prof. Scott L. Friedman (Mount Sinai School of Medicine, USA) for providing LX-2 cells.
Conflict of interest
The authors declare no conflicts of interest related to this work.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Xiao Xiao and Yi Gang contributed equally to this work.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10620_2012_2248_MOESM1_ESM.tif
Fig. S1 Comparison of OPN and TIMP1 expression levels in consecutive sections of cirrhotic liver tissues. Representative images of coordinate expression of OPN and TIMP1 (kappa = 0.518, P = 0.000) were showed (original magnification, 200×). (A, B) Positive OPN staining (A) and positive TIMP1 staining (B) were observed in consecutive sections of cirrhotic liver tissues. (C, D) Negative OPN staining (C) and negative TIMP1 staining (D) were observed in consecutive sections of cirrhotic liver tissues. Supplementary material 1 (TIFF 10,280 kb)
Rights and permissions
About this article
Cite this article
Xiao, X., Gang, Y., Gu, Y. et al. Osteopontin Contributes to TGF-β1 Mediated Hepatic Stellate Cell Activation. Dig Dis Sci 57, 2883–2891 (2012). https://doi.org/10.1007/s10620-012-2248-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10620-012-2248-7