
Abstract
Exclusive enteral nutrition using polymeric formula (PF) is a well-established therapeutic option for active Crohn’s disease; however, its mechanisms of action are unknown. We investigated the anti-inflammatory effects of PF in an in vitro model of epithelial cell inflammation. PF did not affect cell viability over a range of dilutions, but when PF was added to the culture medium the interleukin (IL)-8 response to proinflammatory stimuli was significantly reduced. This effect was due to PF acting directly on the cells as the IL-8 response was still reduced when PF was separated from the proinflammatory stimuli in a 2-compartment system. In the presence of PF, nuclear factor (NF)-κB nuclear migration was not inhibited; however, IκBα degradation was delayed. PF has direct anti-inflammatory effects upon immortalized colonic enterocytes. Therefore PF may, in part, modulate gut inflammation by directly reducing the inflammatory response of the intestinal epithelium.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Crohn’s disease (CD) and ulcerative colitis (UC), collectively known as inflammatory bowel disease (IBD), are chronic illnesses that predominantly affect the gastrointestinal tract. Incidence of IBD, especially CD, has increased in the last decades in developing countries [1–3]. Currently, there is no cure for IBD with this chronic illness characterized by remissions and exacerbations continuing throughout the patient’s life. Although recent research has progressed our understanding of IBD pathogenesis, its etiology remains unknown. The current accepted hypothesis is that interactions between the intestinal epithelium, the innate immune system, and environmental factors including the intestinal microflora, in genetically susceptible individuals, lead to a dysregulated immune response [4].
IBD is often diagnosed during young adulthood, although in up to 25% of patients initial disease presents during adolescence or early childhood, with 2% of pediatric diagnoses occurring before 2 years of age [2, 5, 6]. Classic presenting features of pediatric CD are abdominal pain, diarrhea, and weight loss; if left untreated growth failure and pubertal delay can follow [5, 6]. Growth failure is present in almost 50% of pediatric IBD patients and is caused by malnutrition associated with reduced oral intake, increased energy requirement, increased nutrient loss, and malabsorption [2, 7, 8]. Consequently, attention to nutrition plays a central role in the management of IBD in children [7, 9, 10]. Such an approach may involve the administration of an enteral formula provided exclusively (EEN). Although reports vary with regard to the efficacy of EEN in adults [11–14], there is support for EEN as a treatment modality in children with efficacy equivalent to steroids in the induction of remission in active CD, while avoiding steroid-related side effects [9, 10]. In addition, EEN offers nutritional and linear growth benefits [10, 15]. Although both polymeric and elemental formulae are effective at inducing disease remission [16–18], polymeric formulae (PFs) lead to greater weight gains [18], are more palatable (thus permitting oral administration), and are cheaper than elemental formulae [17].
Over the 2 decades that EEN has been used as a therapeutic option in the management of CD, it is noted to induce disease remission; aid mucosal healing [19] and histologic improvement [20]; has been shown to suppress inflammatory markers [20–23] and increase expression of anti-inflammatory cytokines [24], including transforming growth factor β (TGF-β) [21]. Despite these findings, the precise mechanism by which EEN reduces intestinal inflammation is unknown. The aim of the current study was to further investigate the anti-inflammatory properties of PFs used for EEN and determine if PF has direct anti-inflammatory effects on enterocytes in an in vitro model of intestinal epithelial cell inflammation.
Materials and methods
Materials
Tumor necrosis factor α (TNF-α), interleukin-1α (IL-1α), lipopolysaccharide (LPS), TGF-β, anti–TGF-β, anti–nuclear factor-κB p65 (NF-κB p65) and anti-goat immunoglobulin G (IgG) were obtained from Sigma Chemical Co. (St. Louis, MO). NF-κB SN50 was obtained from Biomol Research Laboratories (New South Wales, Australia). For all experiments, cytokines were used at the following concentrations: TNF-α (100 ng/mL), IL-1 (10 ng/mL), and LPS (1 μg/mL). The PFs tested were Osmolite (Abbott, Columbus, OH), Ensure (Abbott), and Modulen (Nestlé Nutrition, Glendale, CA).
Intestinal epithelial cell lines and culture conditions
HT-29 cells (ATCC HTB-38) were maintained in McCoy’s 5A medium, (Gibco Invitrogen, Carlsbad, CA) containing 10% fetal bovine serum (FBS; Gibco) and 100 U/mL penicillin/streptomycin (Gibco). Caco-2 cells (ATCC HTB-37) were maintained in minimum essential medium with Earle’s salt and l-glutamine (Gibco) containing 20% FBS, 100 U/mL penicillin/streptomycin (Gibco), and 0.1 mmol nonessential amino acids (Gibco). INT407 cells (ATCC CCL-6) were maintained in basal medium Eagle with Earle’s salt (Gibco) containing 10% FBS and 100 U/mL penicillin/streptomycin.
For experimentation, Caco-2 and INT407 cells were seeded in 24-well plates (Becton Dickinson, Franklin Lakes, NJ), with HT-29 cells seeded in 24-well plates (Becton Dickinson) coated with rat tail collagen (BD Biosciences, Bedford, MA). For experiments requiring a 2-compartment system, HT-29 cells were grown to a monolayer on the membrane of inserts (Bio Coat Collagen I inserts 24-well 3.0 micron; BD Biosciences). For immunostaining, HT-29 cells were grown on 4-well culture slides (BD Biosciences) and for the IκB immunoassay, HT-29 cells were grown in 96-well culture plates (Microwell Nunc, Roskilde, Denmark). Cells were incubated in an atmosphere of 5% CO2 in air at 37°C until at least 80% confluent. In each series of experiments, cell-free supernatants were collected and stored at −80°C until required for subsequent analysis.
Assessment of cell viability by trypan blue exclusion
Intestinal epithelial cells (IECs), grown in 24-well culture plates, were washed twice with warm phosphate-buffered saline (PBS, 500 μL/well; Gibco) then incubated with Tryspin-EDTA (500 μL/well; Gibco) for 15 minutes. Warm FBS (500 μL/well) was then added to inhibit trypsin with the contents of each well transferred to a separate eppendorf tube. Tubes were then centrifuged at 1200 g for 5 minutes. The supernatant was discarded and the pellet containing the cells gently resuspended in warm media. Equal volumes of cells and Trypan blue (Sigma) were combined with cell viability counts conducted using a hemocytometer and a light microscope.
Enzyme-linked immunosorbent assay to measure Interleukin-8
The concentration of IL-8 in culture supernatant was measured using a commercial enzyme-linked immunosorbent assay (ELISA; Biosource International, Camarillo, CA) according to the manufacturer’s protocol. Briefly, samples added in duplicate to 96-well microtiter plates (Maxisorp Nunc) coated with monoclonal IL-8 antibody and detected with biotinylated secondary antibody and streptavidin-horseradish peroxidase conjugate (Pharmingen, San Diego, CA). TMB (3,3′,5,5′-tetramethylbenzidine [Sigma] in 0.1 mol citrate acetate buffer with H2O2) was added and the color reaction stopped after 5 minutes with the addition of 5 N H2SO4. A450 nm was measured with an ELISA plate reader (BioRad, Hercules, CA).
Enzyme-linked immunosorbent assay to measure IκBα
Total IκBα protein was measured using a commercial ELISA (SuperArray, Frederick, MD) according to the manufacturer’s protocol. Briefly, HT-29 cells grown to confluence in 96-well microtiter plates were subjected to experimental procedure then fixed with 4% fixing buffer (4% paraformaldehyde [ProSciTech, Thuringowa, Australia;] pH 7, 100 μL/well) overnight at 4°C. Plates were washed with wash buffer (200 μL/well) on a platform mixer (Ratek, Boronia Victoria, Australia) for 5 minutes, after which the wash step was repeated. Plates were quenched by incubating with quenching buffer (100 μL/well) for 20 minutes at room temperature and then washed. Antigen retrieval buffer (100 μL/well) was added and the plates heated in a microwave oven (Samsung, Seoul, Korea) for 3 minutes at 300 V. Plates were allowed to cool, then washed and blocked with blocking buffer (100 μL/well) for 1 hour at room temperature. After washing, IκBα-specific antibody (1:100 dilution, 50 μL/well) was added and the plates incubated for 1 hour at room temperature, then washed once. Secondary antibody (1:16 dilution, 100 μL/well) was added and the plates incubated for 1 hour at room temperature, then washed twice with wash buffer and once with PBS (100 μL/well). Developing solution was added (100 μL/well) with the color reaction proceeding for 10 minutes. Stop solution was added (100 μL/well) with A450 nm measured on an ELISA plate reader (BioRad). Plates were then washed once with wash buffer (200 μL/well) and once with water (200 μL/well) then air dried for 5 minutes. Cell-staining buffer (200 μL/well) was added and plates incubated for 30 minutes at room temperature. Following 2 rinses then 2 washes with water, a 1% sodium dodecyl sulfate solution (100 μL/well) was added and the plate incubated for 1 hour at room temperature, after which A595 nm was measured with a plate reader (BioRad).
Western blot analysis
Fifteen microliters of PFs were combined with 15 μL reducing buffer (5% glycerol [Sigma], 0.03% bromophenol blue [BioRad], 10 mmol dithiothreitol [Sigma]), separated on a 4–20% polyacrylamide gel (Gradipore, Frenches Forest, NSW, Australia) and transferred to a nitrocellulose membrane using the PROTEAN II miniblot system (BioRad). The membrane was blocked by incubating with 3% BSA in PBS overnight at 4°C and probed with anti–TGF-β (1:1000 dilution) for 1 hour at room temperature on a platform mixer. The membrane was washed with Tris-buffered saline (15 mmol Tris-Base [Promega, Madison, WI) and 150 mmol NaCl containing 0.05% Tween 20 (polyoxyethylene [20] sorbitan monolaurate; Ajax Finechem, Seven Hills, NSW, Australia) and incubated with anti-goat IgG (1:5000 dilution) for 1 hour at room temperature on a platform mixer. Following washing, developing substrate (Nitro Blue Tetrazolium [Promega] and 5-bromo-4-chloro-3-indolyl phosphate [Promega] in 100 mmol Tris-Base, 100 mmol NaCl and 5 mmol MgCl2.6H2O, pH 9.5) was added and incubated at room temperature for 5 minutes with the reaction stopped by immersing the membrane in water.
NF-κB immunofluorescence
HT-29 cells were grown on a 4-well culture slide to confluence, washed with PBS then fixed with Zamboni’s fixative (1 mL/well, 1.5% saturated picric acid, 2% formaldehyde in PBS, pH 7.0) for 20 minutes at room temperature. Slides were washed with PBS and incubated with 10% goat serum in 0.1% BSA (Sigma) in PBS for 20 minutes at room temperature. After 2 washes, slides were incubated overnight at 4°C with anti–NF-κB p65 (1:100 in 0.05% saponin [Sigma] BSA/PBS). Following 3 washes, slides were incubated with Alexa Fluor 488 goat anti-mouse Ig (1:200 v/v in saponin/BSA/PBS; Molecular Probes, Eugene, OR) for 2 hours at room temperature in the dark, then washed and mounted in media containing DAPI (Vector Laboratories Inc, Burlingame, CA). Slides were viewed at ×200 and ×400 magnification on an Axioplan 2 Fluorescent microscope (Zeiss, Oberkochen, Germany) with images collected and analyzed using Axio Vision software (Zeiss).
Statistical analysis
GraphPad Prism version 3.00 for Windows (GraphPad Software, San Diego, CA) was used to generate graphs and statistical analysis. ANOVA was used for multiple comparisons, followed by t-test to determine individual differences between groups. Results are presented as mean values±SEM. Significant was accepted if P< .05. All experiments were repeated a minimum of 3 times.
Results
Effect of polymeric formulae on cell viability
HT-29 cells were incubated for 18 hours with PF (formula A, B, or C) at a dilution of 1 in 2, 1 in 5, 1 in 10, 1 in 20, or 1 in 50 in media. Following incubation, cell viability was assessed by Trypan blue exclusion. There was no difference in cell viability over the range of dilutions for any of the formulae (data not shown). Polymeric formula A (PFA) at 1 in 5 dilution in media was used for all further experiments.
Anti-inflammatory effects of polymeric formula A
IEC were incubated with or without PFA for 18 hours to condition cells. Following conditioning, IEC were replenished with similar media (either with or without PFA) containing inflammatory stimuli (TNF-α, IL-1, or LPS) then incubated for a further 6 hours. Following incubation, cell-free culture supernatants were collected and assayed for IL-8. Experiments were repeated at least twice.
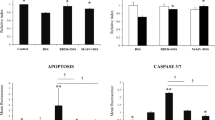
PFA alone did not have an inflammatory effect as similar IL-8 levels were measured in culture with PFA compared to culture with media alone (Fig. 1). However, PFA had a pronounced anti-inflammatory effect; conditioning and maintaining cells in PFA dramatically decreased the IL-8 response to inflammatory stimuli. HT-29 cells, cultured in the absence of PFA produced 10,230±1113 pg/mL IL-8 in response to TNF-α; however, this was reduced to 2205±554 pg/mL IL-8 when cells were conditioned and maintained in PFA and exposed to TNF-α (P< .0001; Fig. 1A). Similarly, HT-29 cells cultured without PFA produced 2610±632 pg/mL IL-8 in response to IL-1, but only 390±126 pg/mL IL-8 when cultured with PFA and stimulated with IL-1 (P< .002). Furthermore, HT-29 cells produced 301±20 pg/mL IL-8 in response to LPS in the absence of PFA and 198±13 pg/mL IL-8 when PFA was present in the culture media (P< .0001; Fig. 1B).


PFA exerts an anti-inflammatory effect on intestinal epithelial cell lines. HT-29 (A, B), Caco-2 (C), and INT407 cells (D) were incubated for 18 hours, with or without PFA, then exposed to TNF-α (100 ng/mL), IL-1 (10 ng/mL), or LPS (1 μg/mL) and incubated for a further 6 hours. Cell-free supernatants were collected with IL-8 levels measured. In the 3 cells lines, PFA significantly reduced the IL-8 response to TNF-α, IL-1, and LPS (* P< .005)
Similar results were observed for Caco-2 and INT407 cells. Caco-2 cells, cultured in media alone, produced 75±14 pg/mL IL-8 in response to TNF-α but only 27±3 pg/mL in response to TNF-α in the presence of PFA (P< .003; Fig. 1C). Caco-2 cells, stimulated with IL-1 produced 217±17 pg/mL IL-8 in the absence of PFA but only 130 ± 20 pg/mL IL-8 in the presence of PFA (P< .003; Fig. 1C). INT407 cells cultured in the absence of PFA produced 5678±1434 pg/mL IL-8 in response to TNF-α, but only 844±90 pg/mL IL-8 when PFA was included in the media (P< .003; Fig. 1D). Also, INT407 cells produced 2201±317 pg/mL IL-8 when cultured with IL-1 and media alone, but only 741±91 pg/mL IL-8 when cultured with IL-1, media and PFA (P< .0001; Fig. 1D).
Further experiments were conducted to determine if PFA maintained its anti-inflammatory effect on unconditioned cells following prolonged exposure to proinflammatory stimulus. HT-29 and INT407 cells were cultured for 18 hours with either TNF-α or IL-1. Following this incubation, media was replenished with fresh media, with or without PFA. Cells were then incubated for a further 6 hours with the culture supernatant collected and assayed for IL-8.
The anti-inflammatory effect of PFA was maintained in unconditioned cells after prolonged exposure to inflammatory stimuli. HT-29 cells produced 1383±135 pg/mL IL-8 in response to TNF-α stimulation, but only 576±34 pg/mL IL-8 upon TNF-α exposure in the presence of PFA (P< .0001). However, IL-1 stimulation produced a similar IL-8 response, with or without PFA, after IL-1 stimulation (Fig. 2A). PFA was effective at reducing the IL-8 response in INT407 cells on exposure to both TNF-α and IL-1. With TNF-α exposure, INT407 cells produced 1134±125 pg/mL IL-8, but only 763±35 pg/mL IL-8 upon TNF-α exposure followed by PFA in the media (P< .01; Fig. 2B). For IL-1 exposure, INT407 cells produced 877±133 pg/mL IL-8 in the absence of PFA and only 540±65 pg/mL IL-8 in the presence of PFA (P=.02; Fig. 2B).

The anti-inflammatory effect of PFA was maintained after intestinal epithelial cells were preexposed to inflammatory stimuli. (A) HT-29 and (B) INT407 cells were pretreated with a proinflammatory stimulus (TNF-α [100 ng/mL] or IL-1 [10 ng/mL]) for 18 hours. Cells were then incubated with fresh media, with or without PFA, for a further 6 hours with supernatants collected and assayed for IL-8. After inflammatory stimulus exposure, PFA still had an anti-inflammatory effect on HT29 cells in response to TNF-α (* P< .0001) and on INT407 cells in response to both TNF-α and IL-1 (** P< .02)
Separation of polymeric formula A and inflammatory stimuli in a two-compartment system
To determine if PFA had direct effects on IECs, PFA was separated from TNF-α. Conditioned HT-29 cells, grown on inserts, were exposed to PFA in the apical compartment and TNF-α in the basal compartment then incubated for 6 hours. Supernatants were collected from both compartments and assayed for IL-8. PFA exerted an anti-inflammatory effect, as IEC cells secreted 3085±777 pg/mL IL-8 basally in the absence of PFA but only 750±136 pg/mL IL-8 when PFA was present (P< .007; Fig. 3).

The anti-inflammatory effect of PFA is maintained in a 2-compartment system. TNF-α and PFA were separated in a 2-compartment system with HT-29 cells grown as a monolayer on a membrane support, separating an apical and a basal compartment. Cells were conditioned with PFA in the basal compartment for 18 hours, then incubated for 6 hours with TNF-α (100 ng/mL) in the apical compartment and PFA in the basal compartment. Cell-free supernatants were collect and assayed for IL-8. PFA still exerted its anti-inflammatory effect on HT-29 cells when separated from TNF-α, as significantly less IL-8 was excreted basally when cells were in contact with PFA (* P< .007)
Detection of TGF-β in polymeric formula by western blotting
Previous studies have demonstrated that PFs containing TGF-β have positive effects on mucosal healing and disease remission in IBD [21]. Western blotting was used to determine if formulae A, B (PFB), or C (PFC) contained TGF-β. TGF-β was detected in PFC but not in PFA or PFB (data not shown). These findings are consistent with the manufacturer’s statement that PFC contains bovine TGF-β.
Effect of heating or freezing/thawing on polymeric formula A
To determine if the anti-inflammatory component of PFA is heat sensitive, PFA was frozen and thawed or heated at 100°C for 20 minutes. Fresh or treated PFA was then added to HT-29 in the presence of TNF-α and incubated for 6 hours. There was no difference in the level of IL-8 produced by HT-29 cells in response to TNF-α, in the presence of fresh, frozen/thawed, or heated PFA (data not shown).

PFA delays IκBα degradation in response to TNF-α stimulation. HT-29 cells stimulated with TNF-α for 15, 30, 45, 60, 90, or 120 minutes with or without PFA were fixed with total IκBα levels measured. Total IκBα levels are presented as percentage of total IκBα levels in unstimulated (baseline) cells. (A) In the absence of PFA after 30 minutes of TNF-α stimulation, IκBα levels were at their lowest at 47% (P=.0473) of baseline, then returned to baseline levels after 60 minutes of stimulation. (B) In the presence of PFA, IκBα degradation was delayed 15 minutes, where levels reached their lowest point (40%, P=.0189) after 45 minutes of stimulation. IκBα levels then returned to baseline at 60 minutes but were elevated compared to baseline after 90 minutes (163%, P=.0243) of TNF-α stimulation
Effect of polymeric formula A on nuclear localization of NF-κB p65
To determine if the anti-inflammatory effect of PFA was due to inhibition of NF-κB nuclear localization, HT-29 cells, stimulated with TNF-α for periods of 15, 30, 45, 60, 90, or 120 minutes in the presence or absence of PFA were fixed and stained with anti-NF-κB p65. In unstimulated cells, NF-κB was observed only in the cytoplasm. After 30 minutes of TNF-α exposure, NF-κB was predominantly seen in and around the nucleus. After 120 minutes of exposure, NFκB was observed in the cytoplasm with very minimal nuclear staining (data not shown). Exposure to PFA in the absence of TNF-α did not appear to induce NF-κB nuclear localization as NF-κB staining appeared predominantly cytoplasmic following 30 minutes exposure to PFA alone. PFA also did not inhibit NF-κB nuclear localization; after 30 minutes exposure to TNF-α in the presence of PFA, NF-κB staining was predominantly nuclear. Pretreatment with SN50 inhibited NF-κB nuclear localization. Following 30 minutes of exposure to TNF-α, NF-κB staining was predominantly cytoplasmic in HT-29 cells pretreated with SN50 (data not shown).
Effect of polymeric formula A on IκBα degradation
Further studies investigated whether PFA affected IκBα degradation. Levels of total IκBα in HT-29 cells were measured following 15-, 30-, 45-, 60-, 90-, or 120-minute exposure to TNF-α in the presence or absence of PFA. In the absence of PFA after 30 minutes of TNF-α exposure, IκBα levels fell to 47% (P=.0473) of unstimulated (baseline) IκBα levels (Fig. 4A). Total IκBα remained low at 61% (P> .05) after 45 minutes exposure then returned to baseline levels after 60 minutes (116%, P> .05), and remand constant following 90 (132%, P> .05) and 120 minutes (106%, P> .05) of exposure to TNF-α (Fig. 4A). Incubation of HT-29 cells with PFA alone had no effect on IκBα levels over the 120-minute period (data not shown). However, TNF-α stimulation in the presence of PFA delayed the degradation of IκBα. IκBα level after 30 minutes of exposure to TNF-α with PFA was 79% of baseline (P> .05). IκBα was significantly decreased after 45 minutes exposure to TNF-α with PFA: levels were 40% of baseline (P=.0189; Fig. 4B). Levels then rose at 60 minutes (79%, P> .05) and continued to increase until they were significantly elevated compared to baseline at 90 minutes (163%, P=.0243) with levels beginning to fall by 120 minutes (152%, P> .05; Fig. 4B).
Discussion
EEN is used in many pediatric centers to induce remission in children with active CD. Although utilized for many years, the precise mechanisms of action of this therapy have not been clearly elucidated. Recent reports show that EEN may lead to alterations in intestinal microflora [25]. The results of the current studies and a previous report by Meister et al. [24] indicate that PFs, as used for EEN, have specific anti-inflammatory properties. It is likely, therefore, that EEN has direct anti-inflammatory effects on the intestinal epithelium in addition to indirect effects secondary to modification of intestinal microflora.
The model utilized in this study is similar to previously published models used to study the intestinal epithelial response in vitro [24]. Here, immortalized colonic epithelial cells were grown as a monolayer to imitate the epithelial cell lining of the colonic mucosa. TNF-α, IL-1, and LPS, important proinflammatory stimuli involved in IBD pathogenesis [20, 26, 27], were used to mimic inflammation and IL-8, a sensitive indicator of inflammation and marker of NF-κB activation [28], was used to quantitate the epithelial cell response. A dilution of 1 in 5 of PF was considered to represent a physiologic dilution as the average adolescent receives 2 L/d PF as IBD therapy, and has a daily intestinal luminal fluid turnover of approximately 10 L [29]. With this model, PF was shown to have no effect on cell viability and PF itself did not induce an inflammatory response. However, PF was shown to significantly reduce IL-8 production in response to a range of proinflammatory stimuli in 3 different epithelial cell lines.
The nature of this anti-inflammatory effect was investigated further. The action of PF could be due to either an intracellular mechanism or an interference of inflammatory stimuli binding with their respective receptors. To clarify this, PF and inflammatory stimuli were separated in a 2-compartment system. In this system, epithelial cells were in contact with TNF-α only through their upper surface and PF only through the lower surface. Here the epithelial cells still produced significantly less IL-8 in response to stimulation when in contact with PF compared to media alone. Therefore, it is likely that PF acts directly on the epithelial cells to reduce IL-8 production.
The NF-κB pathway, a central intracellular signal transduction pathway, was considered a possible target for the intracellular action of PF. Schreiber et al. [30] described increased levels of NF-κB p65 in the lamina propria of patients with IBD, and demonstrated that levels decrease during corticosteroid administration. These findings have been confirmed by other investigators [31]. NF-κB exists as either a homodimer or heterodimer of Rel family proteins including p50/100, p52/p100, p65, RelB, and c-Rel, and is retained in the cytoplasm of unstimulated cells in a complex with IκB proteins [32]. Surface receptor activation, by ligation to proinflammatory stimuli, leads to a cascade of phosphorylation events resulting in the phosphorylation and degradation of IκBα by the ubiquination pathway [33]. Phosphorylation of IκBα releases NF-κB, which then translocates to the nucleus and promotes the transcription of multiple proinflammatory and immunoregulatory genes, including IL-8 [33]. We initially investigated whether PF interferes with NF-κB nuclear translocation. The results indicated that in the presence of PF, NF-κB nuclear translocation proceeds uninhibited. Next we investigated whether PF affected IκBα degradation. Our results indicate that PF does not inhibit IκBα phosphorylation or degradation, but does appears to influence the rate of these events and promotes IκBα expression. The delay in IκBα phosphorylation and enhanced IκBα production may, in part, account for the anti-inflammatory properties of PF.
To investigate this further, we tested for the presence of anti-inflammatory cytokine TGF-β in PF. Fell et al. [21] reported that 79% of children with active CD went into clinical remission after 8 weeks of treatment with PF containing TGF-β. Indeed, TGF-β is an essential cytokine in maintaining mucosal integrity. TGF-β inhibits IEC growth, stimulates IEC differentiation, [34, 35] regulates gut inflammation [36], and acts on tight junctions to limit intestinal permeability [37, 38]. TGF-β was present in only 1 of the 3 PFs used in this study. However, PFA—used for the majority of experiments—displayed anti-inflammatory properties in the absence of TGF-β. Further, freeze–thawing or boiling did not destroy the anti-inflammatory activity of PFA, suggesting that its anti-inflammatory component is not readily labile. The NF-κB pathway can be inhibited by a range of molecules that interact at different locations in the pathway. An example is aspirin and its metabolite sodium salicylate, which inhibit NF-κB activation by binding IκBα and preventing its phosphorylation and degradation [39]. A component of PF may similarly interact with IκBα and may therefore reduce its functionality.
In summary, the use of EEN in the treatment and management of IBD is accepted, although its mode of action is poorly understood. What is known is that EEN offers nutritional benefits and may limit intestinal inflammation by altering intestinal microflora [25]. EEN has also been shown to influence the ratio of proinflammatory and anti-inflammatory cytokines to favor an anti-inflammatory intestinal environment [24]. In addition, we have shown that PF, used for EEN, has direct anti-inflammatory effects on enterocytes, limiting their chemokine response to proinflammatory stimuli. Consequently, this work has provided further evidence to support the validity of EEN as a treatment modality in CD. Continued investigation into the anti-inflammatory mechanism of PF must now be undertaken to help improve the therapeutic properties of PF.
References
Binder V (2004) Epidemiology of IBD during the twentieth century: an integrated view. Baillieres Best Pract Res Clin Gastroenterol 18:463–479
Kim SC, Ferry GD (2004) Inflammatory bowel diseases in pediatric and adolescent patients: clinical, therapeutic, and psychosocial considerations. Gastroenterology 126:1550–1560
Phavichitr N, Cameron DJ, Catto-Smith AG (2003) Increasing incidence of Crohn’s disease in Victorian children. J Gastroenterol Hepatol 18:329–332
Schmidt C, Stallmach A (2005) Etiology and pathogenesis of inflammatory bowel disease. Minerva Gastroenterol Dietol 51:127–145
Lissauer T, Clayden G (2004) Gastroenterology. In: Illustrated textbook of paediatrics. Mosby, St. Louis, pp 182–183
Griffiths AM (2004) Specificities of inflammatory bowel disease in childhood. Baillieres Best Pract Res Clin Gastroenterol 18:509–523
Goh J, O’Morain CA (2003) Review article: nutrition and adult inflammatory bowel disease. Aliment Pharmacol Ther 17:307–320
Thomas AG, Taylor F, Miller V (1993) Dietary intake and nutritional treatment in childhood Crohn’s disease. J Pediatr Gastroenterol Nutr 17:75–81
Heuschkel RB, Menache CC, Megerian JT, Baird AE (2000) Enteral nutrition and corticosteroids in the treatment of acute Crohn’s disease in children. J Pediatr Gastroenterol Nutr 31:8–15
Ruuska T, Savilahti E, Maki M, Ormala T, Visakorpi JK (1994) Exclusive whole protein enteral diet versus prednisolone in the treatment of acute Crohn’s disease in children. J Pediatr Gastroenterol Nutr 19:175–180
Lochs H, Steinhardt HJ, Klaus-Wentz B, Zeitz M, Vogelsang H, Sommer H, Fleig WE, Bauer P, Schirrmeister J, Malchow H (1991) Comparison of enteral nutrition and drug treatment in active Crohn’s disease. Results of the European Cooperative Crohn’s Disease Study. IV. Gastroenterology 101:881–888
Malchow H, Steinhardt HJ, Lorenz-Meyer H, Strohm WD, Rasmussen S, Sommer H, Jarnum S, Brandes JW, Leonhardt H, Ewe K (1990) Feasibility and effectiveness of a defined-formula diet regimen in treating active Crohn’s disease. European Cooperative Crohn’s Disease Study III. Scand J Gastroenterol 25:235–244
O’Morain C, Segal AW, Levi AJ (1984) Elemental diet as primary treatment of acute Crohn’s disease: a controlled trial. Br Med J 1859–1862
Zoli G, Care M, Parazza M, Spano C, Biagi PL, Bernardi M, Gasbarrini G (1997) A randomized controlled study comparing elemental diet and steroid treatment in Crohn’s disease. Aliment Pharmacol Ther 11:735–740
Day AS, Whitten KE, Lemberg DA, Clarkson C, Vitug-Sales M, Jackson R, Bohane TD (2004) Exclusive enteral nutrition as primary therapy in children with Crohn’s disease. J Gastroenterol Hepatol 19:A277
Griffiths AM, Ohlsson A, Sherman PM, Sutherland LR (1995) Meta-analysis of enteral nutrition as a primary treatment of active Crohn’s disease. Gastroenterology 108:1056–1067
Verma S, Brown S, Kirkwood B, Giaffer MH (2000) Polymeric versus elemental diet as primary treatment in active Crohn’s disease: a randomized, double-blind trial. Am J Gastroenterol 95:735–739
Ludvigsson JF, Krantz M, Bodin L, Stenhammar L, Lindquist B (2004) Elemental versus polymeric enteral nutrition in paediatric Crohn’s disease: a multicentre randomized controlled trial. Acta Paediatr 93:327–335
Beattie RM, Bentsen BS, MacDonald TT (1998) Childhood Crohn’s disease and the efficacy of enteral diets. Nutrition 14:345–350
Breese EJ, Michie CA, Nicholls SW, Murch SH, Williams CB, Domizio P, Walker-Smith JA, MacDonald TT (1994) Tumor necrosis factor alpha-producing cells in the intestinal mucosa of children with inflammatory bowel disease. Gastroenterology 106:1455–1466
Fell JM, Paintin M, Arnaud-Battandier F, Beattie RM, Hollis A, Kitching P, Donnet-Hughes A, MacDonald TT, Walker-Smith JA (2000) Mucosal healing and a fall in mucosal pro-inflammatory cytokine mRNA induced by a specific oral polymeric diet in paediatric Crohn’s disease. Aliment Pharmacol Ther 14:281–289
Breese EJ, Michie CA, Nicholls SW, Williams CB, Domizio P, Walker-Smith JA, MacDonald TT (1995) The effect of treatment on lymphokine-secreting cells in the intestinal mucosa of children with Crohn’s disease. Aliment Pharmacol Ther 9:547–552
Ferguson A, Glen M, Ghosh S (1998) Crohn’s disease: nutrition and nutritional therapy. Baillieres Clin Gastroenterol 12:93–114
Meister D, Bode J, Shand A, Ghosh S (2002) Anti-inflammatory effects of enteral diet components on Crohn’s disease-affected tissues in vitro. Dig Liver Dis 34:430–438
Lionetti P, Callegari ML, Ferrari S, Cavicchi MC, Pozzi E, de Martino M, Morelli L (2005) Enteral nutrition and microflora in pediatric Crohn’s disease. J Parenter Enteral Nutr 29:S173–S178
Caradonna L, Amati L, Magrone T, Pellegrino NM, Jirillo E, Caccavo D (2000) Enteric bacteria, lipopolysaccharides and related cytokines in inflammatory bowel disease: biological and clinical significance. J Endotoxin Res 6:205–214
Sartor RB (1997) Pathogenesis and immune mechanisms of chronic inflammatory bowel diseases. Am J Gastroenterol 92:5S–11S
Hoffmann E, Dittrich-Breiholz O, Holtmann H, Kracht M (2002) Multiple control of interleukin-8 gene expression. J Leukoc Biol 72:847–855
Chang EB, Sitirin MD, Black DD (1996) Gastrointestinal, hepatobiliary, and nutritional physiology. Philadelphia: Lippincott-Raven
Schreiber S, Nikolaus S, Hampe J (1998) Activation of nuclear factor kappa B inflammatory bowel disease. Gut 42:477–484
Auphan N, DiDonato JA, Rosette C, Helmberg A, Karin M (1995) Immunosuppression by glucocorticoids: inhibition of NF-kappa B activity through induction of I kappa B synthesis. Science 270:286–290
Baeuerle PA, Baltimore D (1996) NF-kappa B: ten years after. Cell 87:13–20
Baldwin ASJ (1996) The NF-kappa B and I kappa B proteins: new discoveries and insights. Annu Rev Immunol 14:649–683
Oz HS, Ray M, Chen TS, McClain CJ (2004) Efficacy of a transforming growth factor beta 2 containing nutritional support formula in a murine model of inflammatory bowel disease. J Am Coll Nutr 23:220–226
Campos FG, Waitzberg DL, Teixeira MG, Mucerino DR, Habr-Gama A, Kiss DR (2002) Inflammatory bowel diseases: principles of nutritional therapy. Rev Hosp Clin Fac Med Sao Paulo 57:187–198
Kulkarni AB, Huh CG, Becker D, Geiser A, Lyght M, Flanders KC, Roberts AB, Sporn MB, Ward JM, Karlsson S (1993) Transforming growth factor beta 1 null mutation in mice causes excessive inflammatory response and early death. Proc Natl Acad Sci U S A 90:770–774
Dignass AU, Podolsky DK (1993) Cytokine modulation of intestinal epithelial cell restitution: central role of transforming growth factor beta. Gastroenterology 105:1323–1332
Planchon SM, Martins CA, Guerrant RL, Roche JK (1994) Regulation of intestinal epithelial barrier function by TGF-beta 1. Evidence for its role in abrogating the effect of a T cell cytokine. J Immunol 153:5730–5739
Kutuk O, Basaga H (2004) Aspirin inhibits TNFalpha- and IL-1–induced NF-kappaB activation and sensitizes HeLa cells to apoptosis. Cytokine 25:229–237
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
de Jong, N.S.H., Leach, S.T. & Day, A.S. Polymeric Formula Has Direct Anti-Inflammatory Effects on Enterocytes in an in VitroModel of Intestinal Inflammation. Dig Dis Sci 52, 2029–2036 (2007). https://doi.org/10.1007/s10620-006-9449-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10620-006-9449-x