
Abstract
Glucose-regulated protein 78 (GRP78) has been implicated in the protection of tumor cells from cytotoxic damage and apoptosis and thus assists cells in survival under oxygen-deprivation and nutrient-stress conditions. However, its expression and potential role in gastric cancer development and progression have not been reported. In the present study, we determined the level of GRP78 expression in the primary tumor in 86 cases of resected gastric cancer by using immunohistochemistry and analyzed the relationships between GRP78 and clinicopathological characteristics. We found that GRP78 was overexpressed in the tumor specimens when compared with the expression in adjacent tumor-free gastric mucosa. Furthermore, the level of GRP78 expression in both primary tumors and metastatic lymph nodes was inversely correlated with patient survival. Overexpression of GRP78 was directly correlated with Sp1 expression and increased lymph node metastasis. Knocking down GRP78 expression inhibited tumor cell invasion in vitro and growth and metastasis in a xenograft nude mouse model. Therefore, our data imply that dysregulated expression of GRP78 may contribute to the development and progression of gastric cancer.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Gastric cancer is one of the most common malignancies worldwide, with approximately 876,000 new cases diagnosed each year [1]. In Asia and parts of South America, it is the most common epithelial malignancy and leading cause of cancer-related death. Currently, the only curative treatment of gastric cancer is surgical resection when the disease is diagnosed at an early stage. Despite the advances in treatment and research efforts over the past few decades, the outcome of gastric cancer remains poor. The overall 5-year survival rate in the United States and most other western countries ranges from 5 to 15% [2], largely because many gastric cancers are diagnosed at an advanced stage. The aggressive nature of human gastric carcinoma is related to a variety of intracellular events, including activation of various oncogenes, inactivation of tumor suppressor genes, and abnormal expression of growth factors and their receptors [3, 4]. These perturbations result in a tremendous growth advantage for gastric cancer cells.
Glucose-regulated protein 78 (GRP78) is a molecular chaperone that is localized in the endoplasmic reticulum (ER). Expression of it can be enhanced under certain stress conditions, including glucose deprivation, blockade of glycosylation, oxidative stress, acidosis, and hypoxia. As a calcium-binding protein with almost 50% homology with its cytoplasmic counterpart heat shock protein 70, GRP78 is essential for protein folding in the ER lumen, not only for translocation of newly synthesized secretory precursors across the ER membrane but also for presentation of aberrant proteins to the proteolytic machinery by coupling with the proteosome [5]. Other physiological functions of GRP78 include regulation of apoptosis; presentation of antigenic peptides to major histocompatibility complex class I molecules; regulation of cell fate by maintaining the intracellular calcium balance among cytosol, ER, and mitochondria; and maintenance of calcium homeostasis in the cell [6, 7].
Recent studies have indicated a potential role for altered GRP78 expression and function in tumor development and progression. Tumor cells are confronted with oxygen deprivation and nutrient stress often, even with extensive angiogenesis. Such stresses lead to the accumulation of inappropriately processed proteins, which collectively sensitize normal cells to apoptosis more readily when compared with cancer cells with elevated GRP78 expression [6, 8, 9]. Induction of GRP78 expression in tumor cells has been shown to protect them against apoptosis and immune attack [8, 10–12] as well as confer drug resistance [13], thus maintaining the viability of tumor cells that are subjected to such stresses [8]. Moreover, investigators have revealed that GRP78 expression is significantly high in aggressive breast cancer [14] and lung cancer [13, 15]. Overexpression of GRP78 appears to promote tumor growth [8, 16], and inhibition of GRP78 function has been shown to attenuate tumor growth both in vitro and in vivo in B/C10ME fibrosarcoma models [8]. However, the molecular basis for alteration of GRP78 expression and their impact on tumor biology remains unclear. In the present study, we examined whether GRP78 contribute to human gastric cancer development and progression.
Materials and methods
Human tissue specimens and patient information
We used human gastric cancer tissue specimens preserved in the Gastric Cancer Tissue Bank and obtained patient information from The University of Texas M. D. Anderson Cancer Center Upper Gastrointestinal Carcinoma Database with approval from Institutional Review Board. The patients’ primary gastric cancer was diagnosed and treated at M. D. Anderson Cancer Center from 1985 to 1998. They had well-documented clinical histories and follow-up information. None of the patients underwent preoperative chemotherapy or radiation therapy. We randomly selected 86 of these patients for the present study. All of them had undergone gastrectomy with lymph node dissection. Fifty-two patients were positive for lymph node metastasis, whereas 34 were negative for it.
Immunohistochemistry
Sections (5 μm thick) of formalin-fixed, paraffin-embedded tumor specimens were deparaffinized in xylene and rehydrated in graded alcohol. Antigen retrieval was performed with 0.05% saponin for 30 min at room temperature. Endogenous peroxidase was blocked for 12 min with 3% hydrogen peroxide in PBS. The specimens were incubated for 20 min at room temperature with a protein-blocking solution consisting of PBS (pH 7.5) containing 5% normal horse serum and 5% bovine serum albumin and then incubated at 4°C in a 1:100 dilution of a goat polyclonal antibody against human GRP78 (sc-1050; Santa Cruz Biotechnology, Santa Cruz, CA) and a rabbit polyclonal antibody against human Sp1 (clone PEP2). The specimens were then rinsed and incubated for 1 h at room temperature with peroxidase-conjugated antigoat or antirabbit IgG. Next, the slides were rinsed with PBS and incubated for 5 min with diaminobenzidine (Research Genetics, Huntsville, AL). The sections were washed three times with distilled water, counterstained with Mayer’s hematoxylin (BioGenex Laboratories, San Ramon, CA), and washed once each with distilled water and PBS. Afterward, the slides were mounted with the use of a Universal Mount (Research Genetics) and examined under a bright-field microscope. A positive reaction was indicated by a reddish-brown precipitate in the nuclei (Sp1) or cytoplasm (GRP78). Depending on the percentage of positive cells and intensity of the staining, Sp1 and GRP78 staining were classified into one of three groups: negative, weak, and strong. Two independent investigators scored the sections (double blinded) [17].
Cell lines and culture conditions
NCI-N87 is obtained from American Type Culture Collection (ATCC, Manassas, Virginia). SK-GT5 was obtained from Dr. Gary K. Schwartz (Memorial Sloan-Kettering Cancer Center, New York, NY). HF-U251MG astrocytoma cells were obtained from Dr. Suyun Huang (M D Anderson Cancer Center, Houston, TX). The cells were maintained in plastic flasks in minimal essential medium supplemented with 10% fetal bovine serum, sodium pyruvate, nonessential amino acids, l-glutamine, and a vitamin solution (Flow Laboratories, Rockville, MD).
Small interfering RNA and transfection
The small interfering RNA for Sp1 (siSp1) and GRP78 mRNA (siGRP78) were prepared according to previous reports [18–21]. A scrambled sequence unrelated to the siSp1 and siGRP78 was used a control. For transfection, SK-GT5 and NCI-N87 cells were plated in six-well plates in 2 ml of medium supplemented with 10% fetal bovine serum. After 18 h, when cells were 50–60% confluent, siSp1 and siGRP78 duplexes at a final concentration of 50 nM were transfected by using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA); and scrambled duplexes were used as controls. Cells were harvested for 48 h after transfection, and the whole-cell extracts were prepared for Western blot analysis of Sp1 and GRP78 protein expression.
Western blot analysis
Whole-cell lysates were prepared from cell cultures after treatment with the siSp1 and siGRP78. Standard Western blotting was performed with antibodies against human Sp1 and GRP78 as described above. Antirabbit and antigoat IgG, horseradish peroxidase-linked F(ab′)2 fragments obtained from a donkey (Amersham Life Sciences, Arlington Heights, IL), were used as secondary antibodies. Equal protein sample loading was monitored by probing the membranes with an anti-GAPDH antibody. The probe proteins were detected with the Amersham enhanced chemiluminescence system according to the manufacturer’s instructions.
Matrigel invasion assay
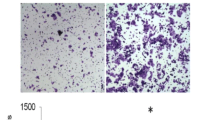
An in vitro invasion assay was performed according to a previously described procedure [22]. BioCoat Matrigel Invasion Chambers (BD, Franklin Lakes, NJ) were primed according to the manufacturer’s directions. Conditioned medium from 1 × 106 HF-U251MG astrocytoma cells cultured for 48 h in modified Eagle’s medium containing 1% fetal bovine serum was placed in the lower well to act as a chemoattractant. Tumor cells (2.5 × 103) in 300 μl of a serum-free medium were placed in the upper chamber of the Matrigel plate and incubated at 37°C for 22 h. Next, the cells on the upper surface of the Matrigel filter were completely removed by using cotton swabs. The migrated cells that penetrated through and remained on the lower surface of the filter were stained with hematoxylin and eosin, counted under microscope at a magnification of 400× in 10 randomly selected fields, and expressed as the mean number of cells per field.
Animals
Female athymic BALB/c nude mice were purchased from Jackson Laboratory (Bar Harbor, ME). The mice were housed in laminar flow cabinets under specific pathogen-free conditions and used when they were 8 weeks old. The animals were maintained in facilities approved by the Association for Assessment and Accreditation of Laboratory Animal Care in accordance with the current regulations and standards of the U.S. Department of Agriculture, Department of Health and Human Services, and National Institutes of Health.
Tumor growth and metastasis
To prepare tumor cells for inoculation, cells in the exponential growth phase were harvested via brief exposure to a 0.25% trypsin/0.02% EDTA solution (wt/vol). Cell viability was determined using trypan blue exclusion, and only single-cell suspensions that were >95% viable were used. Tumor cells (1 × 106 cells/mouse) were then injected into the wall of stomach of nude mice in groups of 10. The animals were killed 60 days after the tumor-cell injection or when they had become moribund. Next, the primary gastric tumors were harvested and weighed. Metastasis to liver and regional lymph nodes was examined by histopathology as described previously [23].
Statistical analysis
The two-tailed χ2-test was performed to determine the significance of the differences between the covariates. Survival durations were calculated by using the Kaplan-Meier method. The log-rank test was used to compare cumulative survival in the patient groups. The patients’ GRP78 and Sp1 expression level, age, sex, Lauren’s classification, American Joint Committee on Cancer stage, and completeness of surgical resection (R0 vs. R1 and R2) were included in the model. The SPSS software program (version 11.05; SPSS Inc., Chicago, IL) was used for the statistical analyses. Student t-test was used to determine the significance of difference in tumor cell invasion in vitro and tumor growth in vivo. In all of the tests, P < 0.05 was defined as statistically significant.
Results
Overexpression of GRP78 in human gastric cancer and its correlation with patient characteristics
We randomly selected the 86 cases and included all stages and histological types of gastric cancer. The median follow-up duration was 25.9 months. At the last follow-up examination, 27 patients were still alive, whereas 59 patients had died. We examined tissue specimens from 86 primary tumors and 52 lymph node metastases. The patients consisted of 56 men and 30 women, and their mean age was 62 years. We observed proximal cancer localization in 20 cases. Regarding the Lauren’s classification, there were 53 intestinal and 33 diffuse-type cancers. Patient characteristics are detailed in Table 1.
We evaluated GRP78 expression in primary cancer tissue specimens obtained from all 86 patients by using immunohistochemistry. We classified the GRP78 expression as negative, weak, or strong. GRP78 was overexpressed in the primary tumors and metastatic lymph nodes as compared with that in the adjacent normal gastric mucosa. Among the primary tumors, we found strong and weak expression in 31 (36%) and 40 (47%) cases, respectively. Fifteen cases (17%) were negative for GRP78 expression. We detected no significant differences in distribution according to gender, residual disease status, or Lauren’s classification among the three GRP78-expression categories (Table 1). Furthermore, there was a significant association between GRP78 expression and pathological type (P = 0.002). However, a statistically significant correlation between the level of GRP78 expression and AJCC stage of gastric cancer was not detected.
Effect of GRP78 expression on survival
A previous study by our group indicated that strong expression of Sp1 is associated with poor outcome of gastric cancer [17]. To determine whether GRP78 expression impacts survival in patients with gastric cancer, we compared the different GRP78 expression levels and survival durations in our patients. The median survival duration in patients who had a tumor with negative, weak, and strong GRP78 expression was 2489, 1242, and 432 days, respectively. The survival duration in patients with weak and strong GRP78 expression was inferior when compared with those with negative expression (Fig. 1A). Likewise, the survival duration in patients with weak and strong GRP78 expression in metastatic lymph nodes was inferior when compared with those with negative expression (Fig. 1B). However, when GRP78 expression, disease stage, completeness of resection, nodal metastasis, Lauren’s classification, and patient age were entered into a Cox proportional hazards model, GRP78 expression was not independently prognostic of poor survival.

Correlation of elevated GRP78 expression with reduced survival in patients with gastric cancer. Shown are Kaplan-Meier plots of overall survival in patients who had a primary tumor (A) or metastatic lymph node (B) with negative, weak, or strong GRP78 expression. The survival curve for the patients who had a primary tumor or metastatic lymph node with negative GRP78 expression was statistically different from that for the patients who had a tumor or metastatic lymph node with weak and strong GRP78 expression
Overexpression of GRP78 and regional lymph node metastasis
Although expression of GRP78 has been shown to be critical to cell survival [12], its role in tumor invasion and metastasis has not been fully defined. Recently, inhibition of GRP78 function in murine fibrosarcoma cells was shown to result in tumor growth impairment [8]. It is also known that patients with a high number of nodal metastases (N2/N3) have a particularly poor prognosis. However, there has been no clinical evidence that GRP78 contributes to human gastric cancer invasion and metastasis. Therefore, we compared the GRP78 expression pattern in patients with N0, N1, and N2/N3 (more than six nodal metastases) disease. In this comparison, N2/N3 disease was associated with strong GRP78 expression. Among patients with N0 and N1 disease, 18 and 35% had strong GRP78 expression, respectively, whereas 52% of the patients with N2/N3 disease had strong GRP78 expression (Table 2). These data suggested that patients with overexpression of GRP78 seemed to have a higher risk of lymph node metastasis than did those without GRP78 overexpression.
Association between GRP78 and Sp1 expression
Transcriptional regulation of GRP78 expression is controlled by several factors, including cellular transcriptional factors, viral proteins, oncogenic proteins, tumor suppressor proteins, and even cytokines [24–28]. Of the transcription factors, Sp1 has been shown to transactivate the GRP78 promoter [29]. Previously, we found that expression of Sp1 in gastric cancer was strongly associated with advanced-stage disease and poor survival [17]. In the present study, we analyzed the correlation between Sp1 and GRP78 expression. Of 69 cases with weak or strong expression of Sp1, 61 (88%) had weak or strong expression of GRP78; this association was statistically significant as analyzed by using Pearson’s χ2-test (P = 0.014) (Table 3). We further confirmed this finding by analyzing consecutive tissue sections. Figure 2 shows representative GRP78 and Sp1 expression in an adjacent tumor-free gastric mucosa (A1 and B1), a primary gastric cancer lesion (A2 and B2), and a metastatic lymph node (A3 and B3). Gastric tumor tissue specimens (both primary and nodal metastases) had a significantly higher level of GRP78 expression than did normal gastric tissue specimens. Moreover, GRP78 expression pattern (Fig. 2C) was consistent with the Sp1 expression pattern (Fig. 2D). Thus, gastric tumors may likely have stronger GRP78 expression in patients with strong Sp1 expression than in those with negative Sp1 expression, indicating that GRP78 is one of the downstream effectors of Sp1 in patients with gastric cancer.

Expression of GRP78 and Sp1 in adjacent tumor-free mucosa, primary gastric cancer, and metastatic lymph nodes. Immunohistochemical staining was performed with polyclonal antibodies against GRP78 (panels A and C) and Sp1 (panels B and D) on tissue sections of primary gastric cancer (panels A1 and B1), adjacent tumor-free mucosa (panels A2 and B2), and metastatic lymph nodes (panels A3 and B3). Two sets of consecutive tissue sections representing strong GRP78 (panels C1 and C2) and Sp1 (panels D1 and D2) expression and negative GRP78 (panels C3 and C4) and Sp1 (panels D3 and D4) expression were also shown
Inhibition of tumor invasion in vitro and growth in vivo by knockdown of GRP78 expression
To directly determine the influence of altered Sp1 expression on GRP78 expression in gastric cancer cells, we transfected SK-GT5 and NCI-N87 gastric cancer cells with the siSp1. As confirmed by Western blot analysis, knockdown of Sp1 expression led to decreased GRP78 expression (Fig. 3A). Our data further confirmed that Sp1 was an important transcription factor for GRP78 expression. Since we have already shown that knockdown of Sp1 retards the growth and metastasis of human gastric cancer in xenograft model, down-regulation of GRP78 might be a contributor to this antitumor activity. However, to provide experimental evidence that GRP78 regulates the growth and metastasis of gastric cancer cells, we determined the impact of decreased expression of GRP78 on tumor cell invasion and metastasis. Both NCI-N87 and SK-GT5 cells were treated with GRP78 siRNA or control. GRP78 expression was substantially suppressed by siGRP78 treatment (Fig. 3B). Their invasive ability was measured using standard Boyden chamber assay and tumorigenicity was determined using orthotopic mouse model. Directly knocking down GRP78 expression inhibited tumor cell invasion in vitro (Fig. 3C) and growth in vivo (Fig. 3D). Additionally, metastasis to liver and regional lymph nodes was also suppressed in tumor cells with reduced GRP78 expression (data not shown).

Inhibition of invasion and growth of human gastric cancer cells by GRP78 knockdown. SK-GT5 and NCI-N87 cells were incubated in a medium alone (Medium) or medium containing a scrambled sequence unrelated to the siSp1 or siGRP78 (Control) or the siSp1 (A) or siGRP78 itself (B) Western blot was performed using total protein lysates. NS: non-specific. Invasion assay was conducted using Matrigel filters. The results shown here were for one representative experiment of three with similar results. Cells on the lower surface of the filter were photographed (C1) The number of migrated cells that penetrated through Matrigel-coated filters was expressed as the mean number of cells in the 10 random fields identified within (C2) Both SK-GT5 and NCI-N87 cells treated as described above (1 × 106/mouse) were injected into the wall of the stomach of nude mice. Tumor sizes were determined as described in Materials and Methods (D) The asterisks indicate statistical significance (P < 0.01) in a comparison between the siGRP78 treated and respective control groups. This was one representative experiment of two with similar results
Discussion
In the present study, we examined the expression of GRP78 protein in human gastric cancer and its relationship with patient characteristics and Sp1 expression. A much higher than normal GRP78 expression level was evident in the resected gastric cancer specimens. GRP78 overexpression in both primary gastric cancer and metastatic lymph node was inversely correlated with patient survival, suggesting that GRP78 serve as a prognostic parameter for gastric cancer. Moreover, the GRP78 expression was strongly correlated with Sp1 expression and lymph node metastasis status. Knockdown GRP78 expression inhibited tumor cell invasion in vitro and growth and metastasis in vivo. Thus, we provided both clinical and experimental evidence that GRP78 plays an important role in gastric cancer progression and metastasis.
Abnormal GRP78 expression and activation may contribute to cancer development and progression. This notion was substantiated by the present study and several previous reports [8, 13–15]. For example, elevated expression of GRP78 in liver tumors was significantly associated with poor prognosis [30]. In the present study, we consistently found that overexpression of GRP78 in human gastric cancer is readily evident, correlates with nodal metastasis, and directly predicts poor survival. However, when GRP78 expression, disease stage, completeness of resection, nodal metastasis, Lauren’s classification, and patient age were entered into a Cox proportional hazards model, GRP78 expression was not independently prognostic of poor survival. Moreover, a statistically significant correlation between the level of GRP78 expression and AJCC stage of gastric cancer was not detected. Since nodal metastasis is one of the factors of AJCC staging, it remains unclear why GRP78 expression affects nodal metastasis but not staging. However, it is likely due to limitations of our small sample size. Alternatively, GRP78 may be more involved in metastasis (N stages) than the growth of local tumors (T stages). In contrast, in patients with neuroblastoma, positive GRP78 expression predicts better survival than does negative GRP78 expression, with a significant difference in the 5-year survival rate. Moreover, a previous multivariate analysis showed that GRP78 expression was an independent prognostic factor in patients with neuroblastoma [31]. Investigators reported a similar observation in patients with lung cancer, in whom GRP78 overexpression appeared to be a favorable prognostic marker [14]. The discrepancy suggests that the role of GRP78 in maintaining the malignant phenotype of various human cancers remains to be defined. However, GRP78 expression appears to be a favorable prognostic factor in tumors that were originally derived from neuroblastic or squamous tissue, whereas GRP78 expression is a poor prognostic factor in adenocarcinoma. Clearly, further investigation should be warranted to determine whether and, if so, how GRP78 plays different roles in the development and progression of tumors from different histological origins and whether GRP78 expression in different pathological types of gastric cancer relates to the infection status of H. Pylori.
The mechanism for the impact of GRP78 alteration on tumor development and progression is still unclear. Several recent studies indicate that GRP78 regulate many aspects of cancer biology. For example, one study correlated GRP78 induction with development of resistance to cytotoxic T lymphocytes and tumor necrosis factor-alpha [32]. Suppression of GRP78 expression with the use of an antisense technique results in increased sensitivity to cytotoxic T lymphocytes and inhibition of the growth of B/C10ME fibrosarcoma cells in vivo [8, 32]. Others believe that elevated GRP78 expression may initiate a tumorigenic pathway through stabilization of proteins with oncogenic potential or resistance to immune response or apoptosis signals [33]. Two reports further supported this notion. First, Chatterjee et al. [34] found that upregulated GRP78 expression in V79 Chinese hamster cells was associated with the development of resistance to clinically useful topoisomerase II-directed agents such as doxorubicin (Adriamycin,) etoposide (VP-16), and m-AMSA. Second, Gomer et al. [35] found that GRP78 expression is associated with cellular resistance to photodynamic therapy. Our current findings may also support this notion, because the majority of the gastric cancer specimens exhibited GRP78 overexpression, which is consistent with the fact that human gastric cancer is commonly refractory to most of the currently available chemotherapeutic agents and has a poor prognosis. However, whether and, if so, how GRP78 is involved in regulation of cell proliferation, apoptosis, and angiogenic property and subsequently leads to poor outcome in patients with gastric cancer warrants further investigation.
Finally, our previous studies showed that Sp1 is overexpressed in gastric cancer and that its high expression level is correlated with advanced disease stage, lymph node metastasis, and predicted poor survival [17]. Sp1 is one of the members of the Sp family of general transcription factors [36]. Interestingly, Abdelrahim et al. [29] found that in Panc-1 pancreatic cancer cells, Sp proteins, including Sp1, Sp3, and Sp4, are constitutively bound to the ER stress response elements, whereas activation of GRP78 protein (or reporter gene) by thapsigargin or tunicamycin is inhibited after co-transfection with small interfering RNAs for Sp1, Sp3, and Sp4, suggesting that Sp1 is critical for GRP78 expression and regulation. In the present study, we analyzed the association between Sp1 and GRP78 expression in clinical gastric cancer specimens and found that strong Sp1 expression has a highly significant association with strong GRP78 expression. Therefore, we have provided first clinical evidence suggesting that GRP78 is a downstream target of Sp1. Our experiments further substantiated this notion by showing that knocking down Sp1 expression led to decreased expression of GRP78. Consistently, altered Sp1 and GRP78 expression have a similar impact on survival of gastric cancer patients. Interestingly, we were the first to show that reduced expression of GRP78 suppressed tumor cell invasion. Although reduced expression of GPR78 rendered the gastric cancer cells more susceptible to apoptosis induction by hypoxia and acidosis (data not shown), the effect on cell invasion did not appear to be due to altered cell survival and proliferation, because GRP78 knockdown did not affect tumor cell viability and proliferation under normal in vitro culture condition (data not shown). Our experimental and clinical findings indicate that elevated expression of GRP78 renders a more malignant phenotype. We are currently investigating the molecular mechanisms by which GRP78 regulates tumor cell migration and invasion.
In summary, we have shown that GRP78 is highly expressed in gastric cancer and appears to be an independent survival predictor in patients with gastric cancer. Our data also indicate a new mechanism; by which altered expression of GRP78 contributes to gastric cancer development and progression. Thus, GRP78 may play an important role in human gastric cancer growth and metastasis and may serve as a cellular target for the development of novel therapeutic approaches.
References
Parkin DM, Bray F, Ferlay J, Pisani P (2005) Global cancer statistics, 2002. CA Cancer J Clin 55:74–108
Berardi R, Scartozzi M, Romagnoli E, Antognoli S, Cascinu S (2004) Gastric cancer treatment: a systematic review. Oncol Rep 11:911–916
Hofler H, Becker KF (2003) Molecular mechanisms of carcinogenesis in gastric cancer. Recent Results Cancer Res 162:65–72
Zheng L, Wang L, Ajani J, Xie K (2004) Molecular basis of gastric cancer development and progression. Gastric Cancer 7:61–77
Gething MJ (1999) Role and regulation of the ER chaperone BiP. Semin Cell Dev Biol 10:465–472
Liu H, Miller E, van de Water B, Stevens JL (1998) Endoplasmic reticulum stress proteins block oxidant-induced Ca2+ increases and cell death. J Biol Chem 273:12858–12862
Little E, Ramakrishnan M, Roy B, Gazit G, Lee AS (1994) The glucose-regulated proteins (GRP78 and GRP94): functions, gene regulation, and applications. Crit Rev Eukaryot Ggene Expr 4:1–18
Jamora C, Dennert G, Lee AS (1996) Inhibition of tumor progression by suppression of stress protein GRP78/BiP induction in fibrosarcoma B/C10ME. Proc Natl Acad Sci USA 93:7690–7694
Reddy RK, Lu J, Lee AS (1999) The endoplasmic reticulum chaperone glycoprotein GRP94 with Ca(2+)-binding and antiapoptotic properties is a novel proteolytic target of calpain during etoposide-induced apoptosis. J Biol Chem 274:28476–28483
Bernstein H, Payne CM, Bernstein C, Schneider J, Beard SE, Crowley CL (1999) Activation of the promoters of genes associated with DNA damage, oxidative stress, ER stress and protein malfolding by the bile salt, deoxycholate. Toxicol Lett 108:37–46
McCormick TS, McColl KS, Distelhorst CW (1997) Mouse lymphoma cells destined to undergo apoptosis in response to thapsigargin treatment fail to generate a calcium-mediated grp78/grp94 stress response. J Biol Chem 272:6087–6092
Reddy RK, Mao C, Baumeister P, Austin RC, Kaufman RJ, Lee AS (2003) Endoplasmic reticulum chaperone protein GRP78 protects cells from apoptosis induced by topoisomerase inhibitors: role of ATP binding site in suppression of caspase-7 activation. J Biol Chem 278:20915–20924
Koomagi R, Mattern J, Volm M (1999) Glucose-related protein (GRP78) and its relationship to the drug-resistance proteins P170, GST-pi, LRP56 and angiogenesis in non-small cell lung carcinomas. Anticancer Res 19:4333–4336
Fernandez PM, Tabbara SO, Jacobs LK, et al (2000) Overexpression of the glucose-regulated stress gene GRP78 in malignant but not benign human breast lesions. Breast Cancer Res Treat 59:15–26
Uramoto H, Sugio K, Oyama T, et al (2005) Expression of endoplasmic reticulum molecular chaperone Grp78 in human lung cancer and its clinical significance. Lung Cancer 49:55–62
Koong AC, Chen EY, Lee AS, Brown JM, Giaccia AJ (1994) Increased cytotoxicity of chronic hypoxic cells by molecular inhibition of GRP78 induction. Int J Radiat Oncol Biol Phys 28:661–666
Wang L, Wei D, Huang S, et al (2003) Transcription factor Sp1 expression is a significant predictor of survival in human gastric cancer. Clin Cancer Res 9:6371–6380
Abdelrahim M, Samudio I, Smith R III, Burghardt R, Safe S (2002) Small inhibitory RNA duplexes for Sp1 mRNA block basal and estrogen-induced gene expression and cell cycle progression in MCF-7 breast cancer cells. J Biol Chem 277:28815–28822
Bond GL, Hu W, Levine A (2005) A single nucleotide polymorphism in the MDM2 gene: from a molecular and cellular explanation to clinical effect. Cancer Res 65:5481–5484
Zu K, Bihani T, Lin A, et al (2006) Enhanced selenium effect on growth arrest by BiP/GRP78 knockdown in p53-null human prostate cancer cells. Oncogene 25:546–554
Dong D, Ko B, Baumeister P, et al (2005) Vascular targeting and antiangiogenesis agents induce drug resistance effector GRP78 within the tumor microenvironment. Cancer Res 65:5785–5791
Huang S, Mills L, Mian B, et al (2002) Fully humanized neutralizing antibodies to interleukin-8 (ABX-IL8) inhibit angiogenesis, tumor growth, and metastasis of human melanoma. Am J Pathol 161:125–134
Wei D, Gong W, Kanai M, et al (2005) Drastic down-regulation of Kruppel-like factor 4 expression is critical in human gastric cancer development and progression. Cancer Res 65:2746–2754
Luo S, Baumeister P, Yang S, Abcouwer SF, Lee AS (2003) Induction of Grp78/BiP by translational block: activation of the Grp78 promoter by ATF4 through and upstream ATF/CRE site independent of the endoplasmic reticulum stress elements. J Biol Chem 278:37375–37385
Cho DY, Yang GH, Ryu CJ, Hong HJ (2003) Molecular chaperone GRP78/BiP interacts with the large surface protein of hepatitis B virus in vitro and in vivo. J Virol 77:2784–2788
Moennikes O, Loeppen S, Buchmann A, et al (2004) A constitutively active dioxin/aryl hydrocarbon receptor promotes hepatocarcinogenesis in mice. Cancer Res 64:4707–4710
Hori O, Matsumoto M, Kuwabara K, et al (1996) Exposure of astrocytes to hypoxia/reoxygenation enhances expression of glucose-regulated protein 78 facilitating astrocyte release of the neuroprotective cytokine interleukin 6. J Neurochem 66:973–979
Jacquier-Sarlin MR, Dreher D, Polla BS (1996) Selective induction of the glucose-regulated protein grp78 in human monocytes by bacterial extracts (OM-85): a role for calcium as second messenger. Biochem Biophys Res Commun 226:166–171
Abdelrahim M, Liu S, Safe S (2005) Induction of endoplasmic reticulum-induced stress genes in Panc-1 pancreatic cancer cells is dependent on Sp proteins. J Biol Chem 280:16508–16513
Lim SO, Park SG, Yoo JH, et al (2005) Expression of heat shock proteins (HSP27, HSP60, HSP70, HSP90, GRP78, GRP94) in hepatitis B virus-related hepatocellular carcinomas and dysplastic nodules. World J Gastroenterol 11:2072–2079
Hsu WM, Hsieh FJ, Jeng YM, et al (2005) GRP78 expression correlates with histologic differentiation and favorable prognosis in neuroblastic tumors. Int J Cancer 113:920–927
Sugawara S, Takeda K, Lee A, Dennert G (1993) Suppression of stress protein GRP78 induction in tumor B/C10ME eliminates resistance to cell mediated cytotoxicity. Cancer Res 53:6001–6005
Dorner AJ, Wasley LC, Kaufman RJ (1992) Overexpression of GRP78 mitigates stress induction of glucose regulated proteins and blocks secretion of selective proteins in Chinese hamster ovary cells. EMBO J 11:1563–1571
Chatterjee S, Cheng MF, Berger SJ, Berger NA (1994) Induction of M(r) 78,000 glucose-regulated stress protein in poly(adenosine diphosphate-ribose) polymerase- and nicotinamide adenine dinucleotide-deficient V79 cell lines and its relation to resistance to the topoisomerase II inhibitor etoposide. Cancer Res 54:4405–4411
Gomer CJ, Ferrario A, Rucker N, Wong S, Lee AS (1991) Glucose regulated protein induction and cellular resistance to oxidative stress mediated by porphyrin photosensitization. Cancer Res 51:6574–6579
Courey AJ, Tjian R (1992) Mechanisms of transcription factor Sp1. In: von Hippel H, Yager TD, Gill SC (eds) Transcription regulation. Cold Spring Harbor, NY, Cold Spring Harbor Laboratory Press, pp 743–769
Acknowledgments
We thank Emily Karotkin for preparation of this manuscript and Don Norwood for editorial comments. This work was supported in part by Research Scholar Grant CSM-106640 from the American Cancer Society and Grant 1R01-CA093829 from the National Cancer Institute (K. Xie) and Grant 1R01-GM69967 from the National Institute of General Medical Sciences (S. Fang), National Institutes of Health.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Zhang, J., Jiang, Y., Jia, Z. et al. Association of elevated GRP78 expression with increased lymph node metastasis and poor prognosis in patients with gastric cancer. Clin Exp Metastasis 23, 401–410 (2006). https://doi.org/10.1007/s10585-006-9051-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10585-006-9051-9