
Abstract
While the individual effects of climate warming, nitrogen (N) deposition, and native plant diversity on plant invasion success are well studied, little is known about how they interact to influence this success. Here, we select invasive Solidago canadensis and four native plant species (Digitaria sanguinalis, Echinochloa crusgalli, Pennisetum alopecuroides, and Polygonum lapathifolium) to assemble five classes of plant communities, each of which is subject to one of the eight treatments consisting of warming and N-addition. Native plants benefit from warming or N-addition more than invasive S. canadensis, and this asymmetry causes warming or N-addition to decrease S. canadensis relative abundance. The inhibitory effect of N-addition on S. canadensis abundance is substantially enhanced by nighttime warming but not by daytime or diurnal warming. The abundance of S. canadensis varies with native plant identity rather than richness, and native plant identity and richness mediate the effects of warming and N-addition. These findings suggest that temperature- and N-dependent asymmetries in growth between invasive and native plants may comprise a mechanism underlying invasion success. This work implies that climate warming and N deposition both could decrease the success of invasive S. canadensis through enhancing invasion resistance, which is contrary to what is often thought.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Human activities are dramatically altering our environment (e.g., global warming, atmospheric pollution, and biological invasion) (IPCC 2015). Our ability to predict the consequences of these activities may depend upon our understanding of human-induced rapid environmental change (IPCC 2015). Plant invasions are among the major threats to biological diversity and ecosystem services (Lockwood et al. 2013). This threat is positively linked to the success of invasive plants: the more successful invasive plants, the more serious their threats. Plant invasion success is driven by a suite of abiotic and biotic factors, and it is crucial to study them for predicting plant invasion and reducing its threats (Dietz and Edwards 2006; Theoharides and Dukes 2007; Catford et al. 2009; Dai et al. 2020). Here, we focused exclusively on air temperature, soil nitrogen (N), and native plants, because they are usually viewed to be key determinants in the invasion process (Lu et al. 2013; Fernandez-Conradi et al. 2018; Potter and Bowman 2020).
Soil N is one of the most key resources, and air temperature is among the most important conditions for plants (Crawley 1997), so they play a crucial role in driving plant invasions (Baker 1965; Davis et al. 2000; Blumenthal 2010; Peng et al. 2019). Atmospheric N deposition increases soil N availability, and climate warming alters thermal regimes, and thus they are particularly relevant to plant invasions (Dukes and Mooney 1999). The leading hypothesis posits that N deposition and climate warming could increase plant invasion success through lowering the environmental constraints (Dukes and Mooney 1999; Walther et al. 2009; Liu et al. 2017); however, opposing evidence is also continuously accumulating (Sorte et al. 2013; Lu et al. 2016). In fact, climate warming is diurnally asymmetric: nighttime temperatures increase faster than daytime temperatures (Davy et al. 2017). This asymmetry can influence the performance of plants and herbivores (Barton and Schmitz 2018; He and He 2020). Few studies, to date, have examined how asymmetric diurnal warming and N deposition interact to influence plant invasions.
Of all biotic factors influencing plant invasions, the importance of native plant species has long been recognized, so multiple hypotheses have been proposed (Levine and D’Antonio 1999; Catford et al. 2009; Jeschke 2014). For example, as early as 1958, Elton posed that a diverse community is more resistant to invasions by alien species than a simple community (i.e., the biodiversity-invasibility hypothesis). The basic assumption of this hypothesis is that the resource availability to alien plants is lower in species-rich communities than in species-poor communities (Elton 1958). In other words, empty niche space is relatively limited in species-rich communities. Kennedy et al. (2002) also proposed that biodiversity can act as a barrier to biological invasions by increasing crowding and species richness in localized plant neighborhoods. While this perception has been prevailing, the effects of native species richness on invasion success remain ambiguous due to the presence of negative, positive, or no relationships between diversity and invasibility (Levine and D’Antonio 1999; Daneshgar and Jose 2009). Such diverse patterns could be linked to the shifts in interspecific relationships (Levine and D’Antonio 1999; Daneshgar and Jose 2009). On the other hand, native plant identity has also been found to be an important factor driving invasion success (Callaway et al. 2003; Daneshgar and Jose 2009; Fernandez-Conradi et al. 2018). However, comparatively limited studies have addressed the relative importance of native species richness and identity in invasion success (but see Daneshgar and Jose 2009).
Plant invasions are always relevant to particular communities, so addressing the community-level performance of invasive and native plants appears to be more useful for understanding invasion success (Shea and Chesson 2002; Maron and Marler 2007; Pearson et al. 2018; Livingstone et al. 2020; Geng and He 2021). Climate warming and N deposition both influence native and invasive plants at the same time, and thus it is necessary to consider native and invasive plants simultaneously. The central question of this study was whether or not climate warming and N deposition can yield synergistic effects on invasion success in the presence of native plant species. We proposed the following hypotheses. (i) Climate warming and N deposition together could generate stronger facilitation than individual environmental changes, because they are usually viewed to facilitate the performance of invasive plants (Dukes and Mooney 1999; Walther et al. 2009; Liu et al. 2017). (ii) Higher species richness might better offset the facilitation of climate warming and N deposition than lower species richness, because species richness is negatively associated to invasibility (Maron and Marler 2007; Zheng et al. 2018). To test these hypotheses, we performed a field experiment, in which different assemblages of native and invasive plants were subject to experimental warming and N-addition. In addition, we examined the effects of warming and N-addition on the leaf defensive potential (i.e., the potential of plants to defend against multiple stresses) of plant invaders, as stresses are widespread (Nejat and Mantri 2017).
2 Materials and methods
2.1 Study species and precultivation
We selected four native plant species and one invasive plant species (Fig. 1: left panel). Digitaria sanguinalis (L.) Scop. (Gramineae) is an annual grass, Echinochloa crusgalli (L.) Beauv. (Gramineae) is an annual grass, Pennisetum alopecuroides (L.) Spreng. (Gramineae) is a perennial grass, Polygonum lapathifolium L. (Polygonaceae) is an annual forb, and Solidago canadensis L. (Asteraceae) is a perennial forb (He and He 2020). The former four species are native to China, common in regional species pools, and dominant in grassland ecosystems (He and He 2020). Solidago canadensis, native to North America, was introduced into China in the 1930s and is among the most serious invasive plants in China (Wan and Liu 2012). All four native plants are non-invasive in the subtropical region in China; S. canadensis originates from the temperate region in North America and is non-native and invasive in China. For simplicity, we called D. sanguinalis, E. crusgalli, P. alopecuroides, and P. lapathifolium as native plants and S. canadensis as an invasive plant.

An illustration of the field experiment including five plant communities, four warming scenarios, and two nitrogen levels. All individuals per mesocosm shape a synthetic plant community, which was subject to each of the eight environmental treatments
The precultivation of experimental plants, which were from seed or rhizome, was conducted at a common garden in March 2019. The seeds of four native species and rhizomes of S. canadensis were field-collected from multiple wild populations in Jiujiang. We collected native Jiujiang topsoil and sifted it free from rock. A mixture of 1:1 native topsoil and sand (ensuring adequate drainage) was filled into plastic trays (50 cm in length × 30 cm in width × 6 cm in depth; one tray per species). To obtain adequate experimental plants, seeds/rhizomes were planted in the above-mentioned trays before transplanting. The trays were placed on benches at the garden. To ensure all plant species to have comparable sizes at the time of transplanting, the seeds of slow-germinating species were sown up to 1–2 weeks before those of fast-germinating species.
Transplanting of the seedlings/ramets was conducted at the garden. In 5 L mesocosms filled with a mixture of 1:1 native topsoil and sand, four individuals from native plant species were evenly planted around the S. canadensis individual, that is, five individuals per mesocosm (Fig. 1: mid-panel). This design allowed all individuals per mesocosm to shape an experimental plant community (Fig. 1: mid-panel). It is noteworthy that we pre-cultured adequate seedlings/ramets so that the transplanted plants were similar in size, particularly for a given species. After transplanting, all mesocosms were immediately moved to the field experimental site. For 2 weeks after initial transplanting, seedlings/ramets that had died were replaced with similar sized seedlings/ramets that had been maintained under the garden condition.
2.2 Experimental set-up
We conducted a field experiment at the Jiujiang experimental site, Jiangxi Province (29.71 °N, 116.04 °E, and 65 m above sea level), where the mean annual precipitation is 1523 mm and the mean annual air temperature is 17.2 °C (He and He 2020). Two global change components were involved: climate warming (four levels, ambient, daytime warming, nighttime warming, and diurnal warming) and atmospheric N deposition (two levels, control and N-addition) (Fig. 1, right panel). Each plant community was subject to one of the eight treatments consisting of temperature and N (Fig. 1: right panel), thereby generating 40 manipulations (5 communities × 4 temperature levels × 2 N levels). Each manipulation was replicated eight times, and thus there were 320 mesocosms (40 manipulations × 8 replicates per manipulation) at the beginning of the experiment.
We set up ambient (no warming), daytime warming (07:00–19:00 h), nighttime warming (19:00–07:00 h), and diurnal warming (07:00–07:00 h). To control for shading by the radiators, we hung a “heater decoy” made out of aluminum sheeting of the same shape and size as the infrared radiators (Kalglo Electronics, Bethlehem, PA, USA) under the ambient condition. Each warming was implemented with a MSR-2420 infrared radiator suspended 1.6 m above ground level. We recorded air temperatures under four warming conditions with data loggers (HOBO MX2302, Onset, USA) at 5-s intervals. Over the course of the experiment, daytime warming increased the average daytime air temperature by 1.5 °C, nighttime warming increased the average nighttime air temperature by 3.6 °C, and diurnal warming increased the average daytime and nighttime air temperatures by 2.3 °C and 3.5 °C, respectively. These increases simulated the projected climate change and were consistent with the observed phenomena that nighttime temperatures increase faster than daytime temperatures (Karl et al. 1993; Easterling et al. 1997; IPCC 2015).
For N treatments, we set up control and N-addition. Ammonium is the dominant form of N in bulk deposition, and the increase rate in nitrate is largest in China (Liu et al. 2013). Ammonium nitrate (NH4NO3) was selected as the N source because it has been widely used in previous studies simulating N deposition (He et al. 2011). According to the previous predictions for future N deposition rates in southern China (Galloway et al. 2004), we used 5.0 g N m−2 year−1 as the N-addition rate. Specifically, N deposition was simulated by surface applications: 1.0 g N m−2 was added in a wet pulse every 2–3 weeks, and the same amount of water was supplied to the control mesocosms.
All the mesocosms were positioned under the ambient condition for 3 weeks to adapt to field conditions and then subject to each of the eight experimental treatments. We weeded and supplied water as needed. Light, precipitation, and soil nutrients except for N were identical for all experimental plants. As a consequence, these practices ensured that temperature and soil N availability were the primary abiotic factors and native species identity or richness was the primary biotic factor. The experiment ran from March to October 2019.
2.3 Measurements of plant traits
Harvesting of the plants began in early October 2019 and was completed in 3 days. We harvested plants and separated them into shoots and roots. All harvested plants were oven-dried at 65 °C for 48 h and weighed. To examine the stoichiometric response of leaves to experimental warming and N-addition, we determined the foliar carbon (C) and N contents of S. canadensis grown with individuals from multiple native plant species but not from one native plant species. This selection could ensure S. canadensis to share the same neighboring species, thereby excluding the effect of native species identity. The oven-dried leaves were ground for measurements of foliar C and N contents using an elemental analyzer (Vario Micro Select, Elementar, Germany). Here, we used C/N ratios to indicate the defensive potential of plants, because this metric can reflect leaf structure against stresses, particularly herbivores (Hanley et al. 2007).
2.4 Data analysis
The total biomass of an individual was defined as the sum of its root and shoot dry biomass. Based on total biomass, we could determine the following two derived variables. The relative species abundance of S. canadensis was calculated as follows: (the total biomass of S. canadensis in a mesocosm/the total biomass of all species in a mesocosm) × 100%. This metric is a good proxy of invasion success at the plant community level (Zheng et al. 2018; Geng and He 2021). To gauge the effect of warming or N-addition on particular species, we calculated the relative biomass change with warming or N-addition as follows: [(TBm – TBu)/TBu] × 100%, where TBm and TBu are the total biomass of a given species grown under the manipulated condition (i.e., a warmed or N-added condition) and the unmanipulated condition (i.e., the ambient or control). Foliar C/N ratio was calculated as the ratio of foliar C content to foliar N content.
The normality of data was tested using the Shapiro-Wilk test and the homogeneity of variance using Bartlett’s test. A generalized linear model, where warming, N-addition, and native species identity were treated as main factors, was used to test their effects on the relative species abundance of S. canadensis. In this model, the four communities, each with one native plant species, were analyzed, and the community with four native plant species was not involved. A generalized linear mixed-effect model, where warming, N-addition, and species richness (i.e., one versus four native species) were treated as fixed factors and species identity as a random factor, was used to test their effects on the relative species abundance of S. canadensis. Note that there were four communities containing one native plant species and one community containing four native plant species. A generalized linear mixed-effect model, where warming and species origin were treated as main factors and species identity as a random factor, was used to test their effects on the relative biomass change of native and invasive species. A normal linear model, where warming and N-addition were treated as main factors, was used to test their effects on the foliar C, N, and C/N of S. canadensis. In addition, the independent sample t-test was used to test whether the relative biomass change with N-addition differed between native and invasive species. Statistical analyses were performed using the R statistical software v3.5.2 with the “stats” and “lme4” package (R Core Team 2018).
3 Results
We first presented the effects of warming, N-addition, and native species identity on the relative species abundance of invasive S. canadensis (upper section in Table 1). Overall, the average decrease in relative species abundance was 28% due to three warming treatments (i.e., daytime, nighttime, and diurnal warming) (Table 1; Fig. 2). More specifically, relative to the ambient (39.8 ± 2.9%, mean ± 1 SE), daytime warming and diurnal warming significantly decreased the relative species abundance by 31% and 42% (all P < 0.05; Fig. 2); nighttime warming decreased this abundance by 11%, although this decrease was not significant statistically (P > 0.05; Fig. 2). N-addition alone significantly decreased the relative species abundance of S. canadensis from 34.6 ± 1.9 to 28.4 ± 1.8% (Table 1; Fig. 2). This relative species abundance was varied with native species identity, ranging from 39.4 ± 2.5% when growing with P. lapathifolium to 19.7 ± 1.4% when growing with P. alopecuroides: this abundance was suppressed in the order P. alopecuroides > D. sanguinalis > P. lapathifolium (Table 1; Fig. 2). Nighttime warming and N-addition jointly decreased the relative abundance of S. canadensis, but such a decrease did not occur in the other combinations of warming and N-addition (Table 1: interaction between warming and N-addition; Fig. 2). Most importantly, warming treatments, N-addition, and native species identity interactively influenced the relative abundance of S. canadensis, so the effects of warming and N-addition on its abundance varied with native species identity (Table 1; Fig. 2).

Effects of warming, N-addition, and native plant species identity on the relative species abundance of Solidago canadensis. Note that four plant communities, each with one native plant species, were used when analyzing the data. Data are means ±1 SE, with full statistical results provided in Table 1. Ds, Digitaria sanguinalis; Ec, Echinochloa crusgalli; Pa, Pennisetum alopecuroides; Pl, Polygonum lapathifolium
We then showed the effects of warming, N-addition, and native species richness on the relative species abundance of S. canadensis (lower section in Table 1). Three warming treatments or N-addition decreased the relative abundance of S. canadensis, and nighttime warming and N-addition jointly decreased its relative abundance (Table 1; Fig. 3). Unlike the native plant identity, native plant richness (one species versus four species) did not affect the relative abundance of S. canadensis (Table 1; Fig. 3: 31.5 ± 1.3% versus 27.7 ± 2.0%). Despite this, native plant richness did mediate the effects of warming and N-addition on relative species abundance (Table 1: significant interactions; Fig. 3). For example, relative to the mean of each individual native species, four native species substantially decreased the relative species abundance of S. canadensis under the ambient condition with N-addition (Fig. 3).

Effects of warming, N-addition, and native plant species richness on the relative species abundance of Solidago canadensis. Note that there were four plant communities containing one native plant species and one plant community containing four native plant species. Data are means ±1 SE, with statistical results provided in Table 1
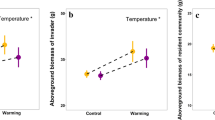
To provide the likely mechanism underlying relative species abundance, we analyzed the effects of warming treatments and N-addition on the growth of native and invasive species. Across two N levels, the warming-induced increase in growth was greater in native plant species (24.1 ± 4.9%) than in S. canadensis (1.9 ± 5.0%) (F species origin = 11.052, P < 0.001; Fig. 4a). This effect depended strongly on warming scenarios (F species origin × warming scenario = 5.563, P = 0.004; Fig. 4a). More specifically, daytime warming enhanced the growth of native and invasive species, but native species benefited from this warming more than S. canadensis (P = 0.078; Fig. 4a). Nighttime warming advantaged the growth of native and invasive species similarly (P = 0.626; Fig. 4a). Diurnal warming increased the growth of native species but decreased the growth of S. canadensis, so the former profited from this warming much more than the latter (P < 0.001; Fig. 4a). While N-addition enhanced the growth of native plant species and S. canadensis across three warming scenarios, this enhancement was much greater in natives than in S. canadensis (F = 12.691, P = 0.001; Fig. 4b). Additionally, there were obvious changes in warming effects among four native species (P < 0.001; Fig. 4a) and in N-addition effects among four native species (P < 0.001; Fig. 4b). This species-specific effect was also varied with warming scenarios (P < 0.001; Fig. 4a).

Effects of warming (a) and N-addition (b) on the growth of native and invasive plant species. Data are means ±1 SE, with statistical results provided in text. Ds, Digitaria sanguinalis; Ec, Echinochloa crusgalli; Pa, Pennisetum alopecuroides; Pl, Polygonum lapathifolium. The green dashed line indicates the mean effect of four native plant species
Additionally, we explored the effects of warming and N-addition on the leaf defensive potential (i.e., C/N ratios) of S. canadensis grown with multiple native plant species. Overall, warming treatments and N-addition independently and opposingly influenced its foliar C, N, and C/N. Warming treatments altered foliar C (F = 3.721, P = 0.019; Fig. 5a), but did not alter foliar N (F = 1.925, P = 0.141; Fig. 5b) and C/N (F = 1.830, P = 0.157; Fig. 5c). We noted that daytime, nighttime, or diurnal warming alone did not alter foliar C, N, and C/N relative to the ambient (all P > 0.05; Fig. 5a–c). Contrary to warming, N-addition did not alter foliar C (F = 0.937, P = 0.339; Fig. 5a), marginally increased foliar N (F = 3.682, P = 0.062; Fig. 5b), and significantly decreased foliar C/N (F = 4.172, P = 0.048; Fig. 5c).

Effects of warming and N-addition on the foliar C content (a), foliar N content (b), and foliar C/N ratio (c) of Solidago canadensis grown with multiple native plant species. Data are means +1 SE, with statistical results provided in test
4 Discussion
Although the effects of climate warming and N deposition on plant invasions have long been detected (Baker 1965; Dukes and Mooney 1999; Walther et al. 2009; Lu et al. 2016), their roles remain to be ambiguous (Sorte et al. 2013; Liu et al. 2017). As opposed to the leading perception that climate warming and N deposition could facilitate plant invasions (Dukes and Mooney 1999; Liu et al. 2017), we found that warming or N-addition per se decreased the relative abundance of invasive S. canadensis, regardless of native plant identity or richness. These findings suggest that climate warming or N deposition alone could suppress plant invasions through enhancing invasion resistance. This pattern could be ascribed to the fact that warming or N-addition differentially advantaged native and invasive species. Specifically, warming and N-addition both increased the growth of native and invasive plants in communities, but S. canadensis benefited from warming and N-addition less than native plants. It is this asymmetry that decreased the relative abundance of the invader but increased the relative abundance of native plants. This work provides evidence for the inhibitory effects of both climate warming and N deposition on plant invasions and also uncovers the related mechanism (i.e., asymmetrically beneficial effects) from a plant community perspective.
Intriguingly, warming treatments interacted with N-addition to decrease the relative abundance of S. canadensis. More specifically, nighttime warming decreased its relative abundance in the presence of N-addition, and daytime and diurnal warming failed to decrease this abundance in the presence of N-addition. As noted above, individual warming or N-addition exhibited negative effects on invasion success. Therefore, it is not surprising that nighttime warming and N-addition acted in concert to decrease invasion success (i.e., an additive effect of the two individual factors). This phenomenon could be attributable partly to asymmetric growth: native plants grew faster under nighttime warming and N-addition than S. canadensis, thereby decreasing its relative abundance.
Warming treatments and N-addition independently and opposingly influenced the leaf structural defensive potential of S. canadensis, as indicated by its foliar C/N, which is a widely used metric in plant defense syndromes and affected by environmental factors (Hanley et al. 2007). Relative to the control treatment, foliar C/N ratios were varied with N-addition, but not with warming treatments. Specifically, individual warming treatments did not alter foliar C and N relative to the ambient, thereby unchanging foliar C/N ratios; N-addition decreased foliar C/N ratios via increasing foliar N and maintaining constant foliar C (i.e., “N dilution” effects, Bidart-Bouzat and Imeh-Nathaniel 2008). These findings support the C/N balance hypothesis (Bryant et al. 1983) and resource availability hypothesis (Coley et al. 1985), but do not support the hypothesis that increased N availability with a warmer climate reduces plant defenses (De Long et al. 2016). Collectively, climate warming exerted neutral effects on the defensive potential of leaves, and N-addition decreased this potential. In nature, abiotic and/or biotic stresses are ubiquitous (Nejat and Mantri 2017). The defensive potential of plants favors them to cope with multiple stresses (Hanley et al. 2007). Therefore, our findings suggest that climate warming might not alter the capacity of plants to tolerate herbivory, but N deposition could decrease this capacity through increasing leaf palatability. Additionally, C/N ratios can also indicate the potential of leaves to photosynthesize: lower C/N ratios mean higher photosynthetic capacity due to higher foliar N contents (Crawley 1997).
We also observed that different warming scenarios exhibited disparate effects on species abundance and foliar stoichiometry. In our warming manipulations, daytime warming increased the daytime air temperature by 1.5 °C, nighttime warming increased the nighttime air temperature by 3.6 °C, and diurnal warming increased the daytime and nighttime air temperatures by 2.3 °C and 3.5 °C, respectively. Therefore, the rising temperatures were diurnally uneven, that is, nighttime temperatures increased faster than daytime temperatures. The relative abundance of S. canadensis was greater under nighttime warming than under diurnal warming, so was foliar C greater under nighttime warming than under diurnal warming. Similar phenomena have been detected in previous studies (He et al. 2012; Li et al. 2012; He and He 2020). For example, daytime, nighttime, and diurnal warming could differentially alter the direction of clonal integration in invasive Alternanthera philoxeroides (Li et al. 2012); diurnal warming caused a lower survival in invasive species than in native species, and daytime and nighttime warming similarly influenced the survival of invasive and native species (He and He 2020). As a result, asymmetric diurnal climate warming might have contrasting effects on native and invasive plants, ultimately influencing the composition and structure of invaded communities.
The relative abundance of S. canadensis was affected by native plant identity, but not by native plant richness. These findings indicate that native plant identity might be more important than native plant richness in driving invasion success, in line with previous findings (Daneshgar and Jose 2009). In fact, the relationships between species richness and community invasibility may be negative, positive, or neutral (Levine and D’Antonio 1999; Daneshgar and Jose 2009). These diverse patterns have contrasting mechanisms and might be attributable primarily to specific settings (cf. Levine and D’Antonio 1999). The role of native plant identity in plant invasions is linked tightly to interspecific interactions such as competition and facilitation (Callaway et al. 2003; Broz et al. 2010; Fernandez-Conradi et al. 2018). We found that the native annual forb P. lapathifolium facilitated the success of invasive S. canadensis, but the native perennial grass P. alopecuroides suppressed its success relative to the native annual grass D. sanguinalis, although these native plant species are sympatric (He and He 2020). Due to the fact that native species were from subtropical areas and invasive S. canadensis was from temperate areas, native and invasive plants had contrasting thermal requirements. These diverse patterns might be linked to contrasting life histories and also highlight the importance of neighboring species per se (i.e., neighborhood effects), supporting the notion that neighbor identity matters (Callaway et al. 2003; Daneshgar and Jose 2009; Fernandez-Conradi et al. 2018).
Native plant identity and richness both strongly modified the joint effects of warming treatments and N-addition. There are a couple of possibilities for explaining these modifications. First, plant identity determines the peculiarity of interspecific interactions (Callaway et al. 2003; Broz et al. 2010; Fernandez-Conradi et al. 2018), and diverse plants can form complex relationship networks (Aschehoug and Callaway 2015). Second, climate warming and N deposition can alter thermal conditions and soil N availability (Dukes and Mooney 1999), which subsequently changes interspecific interactions and their networks (Lu et al. 2013). Finally, warming, N-addition, and native plants could alter soil properties, which can make feedbacks to the performance of native and invasive species (He W.M. unpublished data).
In our long-term experiment with S. canadensis and 16 native plant species under the ambient condition, S. canadensis excluded most of the native plant species after years (He W.M. and He Z.S. unpublished data). Thus, we tentatively speculate that the effects of climate warming and N deposition on S. canadensis could be attenuated over time. In addition, we found an interesting phenomenon that different native plant species (e.g., annual versus perennial plants, or even within annual grasses) exhibited contrasting responses to warming and N addition (Figs. 2, 4). For example, the perennial grass P. alopecuroides had the strongest invasion resistance across all treatments, and the opposite was true for the annual forb P. lapathifolium. These findings suggest that life history might play a key role in determining the responses of plants to climate change. To test the generality of the role of life history under climate change, multiple plant species with the same life history should be selected when conducting the related field experiments.
There are some limitations of our field experiment. More specifically, we only set up two levels of species richness (i.e., 1 versus 4 species) due to limited space and facilities. Thus, our findings cannot be extrapolated to a gradient of species richness. Second, our experiment lasted for one growing season only, so we could not know the multi-year consequences of climate warming and N deposition for invasion success. These limits decrease the general applicability of our findings. Additionally, perennial plants rather than annual plants should be considered in long-term experiments, because plant species with different life histories could respond differentially to experimental manipulations.
In conclusion, the findings from this work might help us to understand the inhibitory effects of global climate warming and atmospheric N deposition on the invasion success of S. canadensis and predict its potential invasion risks under human-induced rapid environmental change. First, climate warming alone or N deposition per se could suppress S. canadensis invasion (i.e., both changes might enhance invasion resistance), contrary to the conventional view that climate warming and N deposition could facilitate plant invasions (Dukes and Mooney 1999; Liu et al. 2017). This suppression might be linked to the asymmetries in growth between native and invasive plants. Second, the inhibitory effects of N deposition on S. canadensis invasion were enhanced by nighttime warming, but not by daytime or diurnal warming. Thus, this inhibitory effect was varied with asymmetric climate warming (i.e., nighttime temperatures increase faster than daytime temperatures), and the timing of climate warming appears to be important for modulating the effects of N deposition. Third, the effects of N deposition on leaf defense were independent of climate warming. N deposition alone decreased the leaf defensive potential of S. canadensis, enabling it to become a weaker defender against stresses (Hanley et al. 2007; Nejat and Mantri 2017). Finally, native plant identity outweighed richness in driving invasions, and both mediated the effects of climate warming and N deposition. It is valuable to consider native plant identity and richness when addressing invasion risks under climate warming and N deposition.
Data availability
The data are available from the corresponding author upon reasonable request.
Code availability
Not applicable.
References
Aschehoug ET, Callaway RM (2015) Diversity increases indirect interactions, attenuates the intensity of competition, and promotes coexistence. Am Nat 186:452–459
Baker HG (1965) Characteristics and modes of origin of weeds. In: Baker HG, Stebbins GL (eds) The genetics of colonizing species. Academic Press, New York, pp 147–168
Barton BT, Schmitz OJ (2018) Opposite effects of daytime and nighttime warming on top-down control of plant diversity. Ecology 99:13–20
Bidart-Bouzat MG, Imeh-Nathaniel A (2008) Global change effects on plant chemical defenses against insect herbivores. J Integr Plant Biol 50:1339–1354
Blumenthal DM (2010) Interactions between resource availability and enemy release in plant invasion. Ecol Lett 9:887–895
Broz AK, Broeckling CD, De-la-Peña C, Lewis MR, Greene E, Callaway RM, Sumner LW, Vivanco JM (2010) Plan neighbor identity influences plant biochemistry and physiology related to defense. BMC Plant Biol 10:115
Bryant JP, Chapin FS, Klein DR (1983) Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 40:357–368
Callaway RM, Mahall BE, Wicks C, Pankey J, Zabinski C (2003) Soil fungi and the effects of an invasive forb on grasses: neighbor identity matters. Ecology 84:129–135
Catford JA, Jansson R, Nilsson C (2009) Reducing redundancy in invasion ecology by integrating hypotheses into a single theoretical framework. Divers Distrib 15:22–40
Coley PD, Bryant JP, Chapin FS (1985) Resource availability and plant antiherbivore defense. Science 230:895–899
Crawley MJ (1997) Plant ecology, 2nd edn. Blackwell Science Ltd, Oxford
Dai ZC, Wan LY, Qi SS, Rutherford S, Ren GQ, Wan JSH, Du DL (2020) Synergy among hypotheses in the invasion process of alien plants: a road map within a timeline. Perspect Plant Ecol Evol Syst 47:125575
Daneshgar P, Jose S (2009) Role of species identity in plant invasions: experimental test using Imperata cylindrica. Biol Invasions 11:1431–1440
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:528–534
Davy R, Esau I, Chernokulsky A, Outten S, Zilitinkevich S (2017) Diurnal asymmetry to the observed global warming. Int J Climatol 37:79–93
De Long JR, Sundqvist MK, Gundale MJ, Giesler R, Wardle DA (2016) Effects of elevation and nitrogen and phosphorus fertilization on plant defence compounds in subarctic tundra health vegetation. Funct Ecol 30:314–325
Dietz H, Edwards PJ (2006) Recognition that causal processes change during plant invasion helps explain conflicts in evidence. Ecology 87:1359–1367
Dukes JS, Mooney HA (1999) Does global change increase the success of biological invaders? Trends Ecol Evol 14:135–139
Easterling DR, Horton B, Jones PD, Peterson TC, Karl TR, Parker DE, Salinger MJ, Rzuvayev V, Plummer N, Jamason P, Folland CK (1997) Maximum and minimum temperature trends for the globe. Science 277:364–367
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London
Fernandez-Conradi P, Borowiec N, Capdevielle X, Castagneyrol B, Maltoni A, Robin C, Selvi F, van Halder I, Vétillard F, Jactel H (2018) Plant neighbour identity and invasive pathogen infection affect associational resistance to an invasive gall wasp. Biol Invasions 20:1459–1473
Galloway JN, Dentener FJ, Capone DG, Boyer EW, Howarth RW, Seitzinger SP, Asner GP, Cleveland CC, Green PA, Holland EA, Karl DM, Michaels AF, Porter JH, Townsend AR, Vörösmarty CJ (2004) Nitrogen cycles: past, present, and future. Biogeochemistry 70:153–226
Geng XM, He WM (2021) Success of native and invasive plant congeners depends on inorganic nitrogen compositions and levels. J Plant Ecol 14:202–212
Hanley ME, Lamont BB, Fairbanks MM, Rafferty CM (2007) Plant structural traits and their role in anti-herbivore defence. Perspect Plant Ecol Evol Syst 8:157–178
He ZS, He WM (2020) Asymmetric climate warming does not benefit plant invaders more than natives. Sci Total Environ 742:140624
He WM, Yu GL, Sun ZK (2011) Nitrogen deposition enhances Bromus tectorum invasions: biogeographic differences in growth and competitive ability between China and North America. Ecography 34:1059–1066
He WM, Li JJ, Peng PH (2012) A congeneric comparison shows that experimental warming enhances the growth of invasive Eupatorium adenophorum. PLoS One 7:e35681
IPCC (2015) Climate change 2014: synthesis report. Cambridge University Press, Cambridge
Jeschke JM (2014) General hypotheses in invasion ecology. Divers Distrib 20:1229–1234
Karl TR, Jones PD, Knight RW, Kukla G, Plummer N, Razuvayev V, Gallo KP, Lindseay J, Charlson RJ, Peterson TC (1993) A new perspective on recent global warming: asymmetric trends of daily maximum and minimum temperature. Bull Am Meteorol Soc 74:1007–1023
Kennedy TA, Naeem S, Howe KM, Knops JMH, Tilman D, Reich P (2002) Biodiversity as a barrier to ecological invasion. Nature 417:636–638
Levine JM, D’Antonio CM (1999) Elton revisited: a review of evidence linking diversity and invasibility. Oikos 87:15–26
Li JJ, Peng PH, He WM (2012) Physical connection decreases benefits of clonal integration in Alternanthera philoxeroides under three warming scenarios. Plant Biol 14:265–270
Liu XJ, Zhang Y, Han WX, Tang A, Shen JL, Cui ZL, Vitousek P, Erisman JW, Goulding K, Christie P, Fangmeier A, Zhang F (2013) Enhanced nitrogen deposition over China. Nature 494:459–463
Liu Y, Oduor AMO, Zhang Z, Manea A, Tooth IM, Leishman MR, van Kleunen M (2017) Do invasive alien plants benefit more from global environmental change than native plants? Glob Chang Biol 23:3363–3370
Livingstone SW, Isaac ME, Cadotte MW (2020) Invasive dominance and resident diversity: unpacking the impact of plant invasion on biodiversity and ecosystem function. Ecol Monogr 90:e01425
Lockwood JL, Hoopes MF, Marchetti MP (2013) Invasion ecology. John Wiley & Sons, New York
Lu XM, Siemann E, Shao X, Wei H, Ding JQ (2013) Climate warming affects biological invasions by shifting interactions of plants and herbivores. Glob Chang Biol 19:2339–2347
Lu XM, Siemann E, He MY, Wei H, Shao X, Ding JQ (2016) Warming benefits a native species competing with an invasive congener in the presence of a biocontrol beetle. New Phytol 211:1371–1381
Maron JL, Marler M (2007) Native plant diversity resists invasion at both low and high resource levels. Ecology 88:2651–2661
Nejat N, Mantri N (2017) Plant immune system: crosstalk between responses to biotic and abiotic stresses the missing link in understanding plant defence. Curr Issues Mol Biol 23:1–16
Pearson DE, Ortega YK, Eren Ö, Hierro JL (2018) Community assembly theory as a framework for biological invasions. Trends Ecol Evol 33:313–325
Peng Y, Yang JX, Zhou XH, Peng PH, Li JJ, Zhang SM, He WM (2019) An invasive population of Solidago canadensis is less sensitive to warming and nitrogen-addition than its native population in an invaded range. Biol Invasions 21:151–162
Potter TS, Bowman WD (2020) Testing invasion filters for the alpine: the roles of temperature, nitrogen deposition and soil. Biol Invasions 22:1889–1901
R Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. URL: http://www.R-project.org
Shea K, Chesson P (2002) Community ecology theory as a framework for biological invasions. Trends Ecol Evol 17:170–176
Sorte CJB, Ibáñez I, Blumenthal DM, Molinari NA, Miller LP, Grosholz ED, Dukes JS (2013) Poised to prosper? A cross-system comparison of climate change effects on native and non-native species performance. Ecol Lett 16:261–270
Theoharides KA, Dukes JS (2007) Plant invasion across space and time: factors affecting nonindigenous species success during four stages of invasion. New Phytol 176:256–273
Walther GR, Roques A, Hulme PE, Sykes MT, Pysek P, Kuhn I, Zobel M, Bacher S, Botta-Dukat Z, Bugmann H (2009) Alien species in a warmer world: risks and opportunities. Trends Ecol Evol 24:686–693
Wan FH, Liu QR (2012) Biological invasions: color illustrations of invasive alien plants in China. Science Press, Beijing
Zheng YL, Burns JH, Liao ZY, Li YP, Yang J, Chen YJ, Zhang JL, Zheng YG (2018) Species composition, functional and phylogenetic distances correlate with success of invasive Chromolaena odorata in an experiment text. Ecol Lett 21:1211–1220
Funding
This work was supported by the National Natural Science Foundation of China (31971552).
Author information
Authors and Affiliations
Contributions
Wei-Ming He and Zheng-Sheng He designed the experiment. Zu-Xing Wang and Zheng-Sheng He performed the experiment. Zu-Xing Wang and Wei-Ming He analyzed the data. Wei-Ming He, Zu-Xing Wang, and Zheng-Sheng He wrote the manuscript.
Corresponding author
Ethics declarations
Ethics approval
Not applicable.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Wang, ZX., He, ZS. & He, WM. Nighttime climate warming enhances inhibitory effects of atmospheric nitrogen deposition on the success of invasive Solidago canadensis. Climatic Change 167, 20 (2021). https://doi.org/10.1007/s10584-021-03175-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10584-021-03175-0