
Abstract
Chromosomal distribution of the Fat element that was isolated from bacterial artificial chromosome (BAC) end sequences of wheat chromosome 3B was studied in 45 species representing eight genera of Poaceae (Aegilops, Triticum, Agropyron, Elymus, Secale, Hordeum, Avena and Triticale) using fluorescence in situ hybridisation (FISH). The Fat sequence was not present in oats and in two barley species, Hordeum vulgare and Hordeum spontaneum, that we investigated. Only very low amounts of the Fat element were detected on the chromosomes of two other barley species, Hordeum geniculatum and Hordeum chilense, with different genome compositions. The chromosomes of other cereal species exhibited distinct hybridisation patterns with the Fat probe, and labelling intensity varied significantly depending on the species or genome. The highest amount of hybridisation was detected on chromosomes of the D genome of Aegilops and Triticum and on chromosomes of the S genome of Agropyron. Despite the bioinformatics analysis of several BAC clones that revealed the tandem organisation of the Fat element, hybridisation with the Fat probe produces uneven, diffuse signals in the proximal regions of chromosomes. In some of the genomes we investigated, however, it also forms distinct, sharp clusters in chromosome-specific positions, and the brightest fluorescence was always observed on group 4 chromosomes. Thus, the Fat element represents a new family of Triticeae-specific, highly repeated DNA elements with a clustered–dispersed distribution pattern. These elements may have first emerged in cereal genomes at the time of divergence of the genus Hordeum from the last common ancestor. During subsequent evolution, the amount and chromosomal distribution of the Fat element changed due to amplification, elimination and re-distribution of this sequence. Because the labelling patterns that we detected were highly specific, the Fat element can be used as an accessory probe in FISH analysis for chromosome identification and investigation of evolutionary processes at the chromosomal level.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Repetitive DNA sequences are the major component of plant DNA. In cereals, they may comprise up to 90–95% of the nuclear genome (Flavell et al. 1974). Repetitive sequences are highly heterogenic and represent hundreds or even thousands of families that differ with respect to the length of the repeat unit, nucleotide composition, copy number and organisation in the genome (Flavell 1986; Heslop-Harrison 2000; Sharma and Raina 2005). The results of total genome sequencing of two model plant species, rice (Oryza sativa) and Arabidopsis and partial genome sequencing of some important crops such as wheat, barley, maize, sorghum, sugar beet and grapevine have revealed that the most abundant class of repetitive sequences in plants is LTR retrotransposons (The Arabidopsis Genome Initiative 2000; Li et al. 2004; Sequencing Project International Rice G 2005; Paux et al. 2006, 2008; Jaillon et al. 2007; Paterson et al. 2009; Schnable et al. 2009). In plant with large genomes, such as maize, wheat and barley, LTR retrotransposons constitute more than 50% of the genome. From 5% to 11% of wheat DNA is made up of DNA transposons, and the remaining classes of repeats constitute nearly 30% of the wheat genome.
Despite the abundance and outstanding diversity of repetitive DNA sequences in plants, little is known about the causes of their emergence and maintenance in plant genomes. Some authors have hypothesised that these sequences play an important role in chromosome organisation, stabilisation of chromosome structure, recognition and segregation of chromosomes in mitosis and meiosis and regulation of gene activity (Vershinin et al. 1995; Heslop-Harrison 2000; Grewal and Moazed 2003; Sharma and Raina 2005). Changes in the amount and distribution of repetitive DNA are one of the driving forces of plant evolution and speciation (Flavell et al. 1979; Dvořák and Dubkovsky 1996; Cuadrado and Jouve 2002).
It is not surprising, therefore, that repetitive sequences are increasingly attracting the attention of researchers. Over the past several decades, many new families of these sequences have been isolated and characterised (Taketa et al. 2000; Linares et al. 2001; Ananiev et al. 2002; Zhang et al. 2004; Lim et al. 2005). The development of a new vector system bacterial artificial chromosome (BAC) for DNA cloning has played a large role in promoting the discovery of new types of repetitive DNAs (Jiang et al. 1995; Li et al. 2004; Zhang et al. 2004; Charles et al. 2008). However, the number of families of repeats that have been identified and characterised is relatively low, and the number of these sequences that are currently being employed in practical plant cytogenetics is even smaller. Therefore, the search for new markers that can be used for chromosome identification and for the analysis of plant divergence and speciation is an important focus of current studies in plant biology.
Among the Triticeae species, the best studied are the repetitive sequences of rye, Secale cereale (Bedbrook et al. 1980; Jones and Flavell 1982a, b; Vershinin et al. 1995; Cuadrado and Jouve 2002) and Aegilops tauschii, the D genome donor of wheat (Rayburn and Gill 1986; Badaeva et al. 1996, 2002; Nagaki et al. 1999; Taketa et al. 2000). Recently, BAC end sequencing of the 3B chromosome of wheat resulted in the identification of a new category of tandem repeats, Fat, а highly conserved 500-bp element representing nearly 0.6% of the wheat genome (Paux et al. 2006). Fat is clustered in direct tandem repeats that are separated by a consensus 9-bp spacer sequence (CYGRTTTDB where Y is C or T, R is A or G, D is A, T or G and B is C, T or G). Each unit of the Fat element is made up of two perfect direct repeats (GAGAAGCT) at both ends and two putative autonomously replicating consensus sequences, which determine the replication origin in yeast, which suggests that amplification of the Fat element could occur through rolling circle-like replication (Paux et al. 2006). Our preliminary results revealed that this element is present in all wheat chromosomes and is the most abundant sequence in the D genome.
In the study we present here, we used fluorescence in situ hybridisation (FISH) analysis in a broad range of cereal species to trace alterations in the amount and distribution of the Fat sequence over the course of cereal evolution and to assess its possible utility for the purpose of chromosomal analysis.
Materials and methods
Materials
In total, 69 accessions from 45 species belonging to eight genera of Poaceae were studied. Twenty-four of these species belong to the genus Aegilops and nine belong to the genus Triticum. In addition, four barley species (Hordeum), two rye species (Secale), three species of the Avena genus and one accession for each of the Triticale, Agropyron and Elymus genera were examined. The genomic constitutions, chromosome numbers and the origin of the materials that we used are presented in Table 1.
DNA probes
A probe for Fat was synthesised using the 3B_050_N05 BAC clone, following PCR amplification, purification and labelling using a nick translation procedure. Two Fat PCR primers (GGGGAGCTTCTCACAACAAGC and TATTTACCACGGCATGTCGGG) were designed based on the sequence of 3B_050_N05_FM1 (GenBank accession number DX374230) and an approximately 460-bp fragment was amplified by PCR (30 cycles, Tm = 60°C). Additional probes, pSc119.2 (Bedbrook et al. 1980), pAs1 (Rayburn and Gill 1986), pTa71 (Gerlach and Bedbrook 1979) and pTa794 (Gerlach and Dyer 1980), were used for chromosome/genome identification.
Fluorescence in situ hybridisation
Chromosomal preparations and FISH were carried out as described previously (Badaeva et al. 1996) with the following modifications. The Fat sequence was labelled with fluorescein-12-dUTP and detected with anti-fluorescein/Oregon Green rabbit IgG and Alexa Fluor® 488 conjugate (Invitrogen, Carlsbad, CA). The other probes used were labelled with biotin-16-dUTP by nick translocation according to the manufacturer's protocol (Roche, Germany) and detected with streptavidin-Cy3 (Amersham Biosciences, Piscataway, NJ). The slides were counterstained with 2 μg ml−1 4′,6-diamidino-2-phenylindole in 1× phosphate-buffered saline, mounted in Vectashield medium (Vector Laboratories, Peterborough, UK) and examined on an Imager D1 microscope (Carl Zeiss, Germany). From five to 12 metaphase cells from each slide with clear, reproducible signals were selected and captured with an AxioCam HRm black-and-white camera using the AvioVision software, release 4.6.Chromosomes were identified on the basis of labelling patterns produced with the pSc119.2, pAs1, pTa71 and pTa794 probes and classified according to standard nomenclatures developed for each respective species (Cabrera et al. 1995; Badaeva et al. 1996, 2002, 2004; Bardsley et al. 1999; Cuadrado and Jouve 2002).
Results
Wheat
Our previous data showed that hybridisation of the Fat probe on chromosomes of common wheat produces strong, non-uniform fluorescence on 14 chromosomes of the D genome, and the remaining 28 chromosomes exhibit only pale, fuzzy hybridisation signals (Paux et al. 2006). Chromosomes of the A genome show only slightly higher fluorescence levels than B genome chromosomes (Fig. 1a; Supplementary material 1a). In addition to the diffuse signals produced, some D genome chromosomes also contain sharp, brilliantly fluorescing regions. The largest of these is detected in the short arm of chromosome 4D and several additional smaller sites are present on the short and long arms of this chromosome. Two sharp, brilliant signals are seen on the short arm of chromosome 1D. Chromosomes 6D and 7D are both marked by intense, fuzzy labelling of the proximal regions of both arms. Chromosomes 2D, 3D and 5D lacked such brilliant signals, though they were also marked by bright, non-uniform labelling. The distribution of the Fat element on chromosomes of three diploid wheat species (T. urartu, T. boeoticum and T. monococcum) is similar to the pattern of Fat in the A genome of polyploid wheat and consists of unevenly dispersed signals located in the proximal regions of chromosomes (Fig. 2j, k; Supplementary material 2j, k).

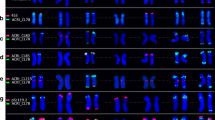
Distribution of the Fat element (green) on chromosomes of polyploid cereal species: a Triticum aestivum (cv. Tumenskaya-80); b Ae. crassa 4x (k-2485); c E. dahuricus; d Ae. ventricosa (SSD32); e Ae. geniculata (IG7761); f A. glaucum; g Ae. juvenalis (i-578009); h Ae. biuncialis (IG47504); i Ae. crassa 6x (k-1005); j Ae. columnaris (TA2084); k Ae. neglecta (Ae. recta, k-2062); l Ae. kotschyi (TA2206); m Ae. cylindrica (k-1319). The chromosomes were co-hybridised with biotinylated probes pSc119.2 (a, h, j, k, l) or pAs1 (b–g, i, m) and detected with streptavidin-Cy3 (red). Scale bar, 10 μm. Chromosomes of polyploid wheat and Aegilops are designated according to (Badaeva et al. 2002, 2004; Schneider et al. 2003); chromosomes of Ae. columnaris with unknown homology and genome affinity are designated with letters (A–J)

Distribution of the Fat element (green) on chromosomes of the diploid Aegilops and rye species and diploid and polyploid barleys: a Ae. bicornis (k-1328); b Ae. caudata (k-1797); c Ae. umbellulata (i-571748); d Ae. longissima (k-907); e Ae. uniaristata (TA2768); f Ae. comosa (i-577976); g Ae. tauschii ssp. tauschii (k-865); h Ae. tauschii ssp. strangulata (TQ27); i Ae. speltoides (#37); j T. urartu (INRA27019); k T. monococcum (cv. Du Pays du Sault); l S. cereale (cv. Dan Nove); m H. chilense (#7); n S. montanum (Israel, El-Kuneitra); p H. geniculatum (2n = 4x = 28). o The distribution of Fat (green) and pAs1 (red) probes on chromosomes of Ae. tauschii ssp. strangulata (TQ27 and i-571716) and ssp. tauschii (k-865). The chromosomes were co-hybridised with biotinylated probes pSc119.2 (a – e, i, l, n), pAs1 (f–h, m, p) and pTa794 (j, k) and detected with streptavidin-Cy3 (red). Scale bar, 10 μm. Chromosomes of rye are classified according to Cuadrado and Jouve (2002), chromosomes of H. chilense are classified according to Cabrera et al. (1995) and chromosomes of Aegilops species are classified according to Badaeva et al. (1996)
Diploid Aegilops
Ae. tauschii (D genome) chromosomes exhibit the strongest Fat element hybridisation intensities that we detected among the diploid Aegilops species. Distribution of the hybridisation signals in this species is similar to what we have seen in the D genome chromosomes of common wheat, though the two subspecies of Ae. tauschii differ in their labelling patterns on chromosomes 1D, 2D and 6D (Fig. 2g, h). In the two studied accessions of the subspecies strangulata, chromosome 1D contains brilliant signals in the distal part of the short arm that are absent in the respective chromosome of the accession k-865 (ssp. tauschii). By contrast, in the latter case, we observe a clear signal in the distal part of the chromosome 6D short arm (Fig. 2o; Supplementary material 2g, h, o). Alteration of pale and brightly fluorescing regions characterises the long and the short arms of chromosome 2D of ssp. tauschii, while in ssp. strangulata, such non-uniform labelling is observed in the long arm only.
Aegilops uniaristata (N genome) and Aegilops comosa (M genome) show a moderate amount of hybridisation with the Fat probe (Fig. 2e, f,), while the intensity of hybridisation to Aegilops umbellulata (U genome) and the Aegilops caudata (C genome) chromosomes is noticeably weaker (Fig. 2b, c; Supplementary material 2b, c, e, f). In species of the Sitopsis (S genome) group, labelling intensity is highest for Aegilops bicornis (Fig. 2a), followed by Aegilops longissima and Aegilops sharonensis (Fig. 2d), and only weak hybridisation is seen in Aegilops speltoides (Fig. 2i; Supplementary material 2a, d, i). For all the genomes we investigated in this group, the Fat element is found in an uneven, diffuse pattern, localised to the proximal regions of all chromosomes, with the brightest fluorescence being seen on group 4 chromosomes.
Polyploid Aegilops of the D genome cluster
FISH analysis of Aegilops cylindrica reveals significant differences in fluorescence intensity of the Fat probe between the C and D genomes (Fig. 1m); hybridisation to chromosomes of the C genome is very poor, but the D genome chromosomes are heavily labelled. The distribution of signals produced by the Fat and pAs1 probes on the D genome chromosomes of Ae. cylindrica is similar to the hybridisation patterns on the chromosomes of diploid Ae. tauschii ssp. tauschii. In another tetraploid species of the D genome cluster, Aegilops ventricosa, all chromosomes hybridise with the Fat element probe, though fluorescence intensity on the Dv genome chromosomes is slightly higher than on the Nv genome chromosomes (Fig. 1d). The hybridisation patterns seen on the D genome chromosomes of both this tetraploid species and the diploid Ae. tauschii ssp. tauschii are virtually the same; however, there are differences in the labelling patterns of some Nv genome chromosomes of Ae. ventricosa compared with Ae. uniaristata (Supplementary material 1d, m; 2e, h, o).
The tetraploid species Aegilops crassa, the most ancient polyploid Aegilops species, originated from hybridisation of Ae. tauschii with an extinct diploid species of the Sitopsis group (Zhang and Dvorak 1992; Badaeva et al. 2002). All Ae. crassa chromosomes hybridise with the Fat probe with approximately the same intensity (Fig. 1b; Supplementary material 1b). However, labelling patterns of the D1 and Xcr genomes differ from their putative diploid ancestors. The overall fluorescence seen on chromosomes 1D1, 3D1, 6D1 and 7D1 is weaker, and chromosome 1D1 does not exhibit any distinct signals in its short arm. Chromosome 4D1 is missing a small distal Fat site that is present in the short arm of the respective chromosome of the ancestral species. Fluorescence intensity of the Xcr genome chromosomes is noticeably higher than in genomes of all species of the Sitopsis group and is comparable with what we have seen on chromosomes in the D1 genome. Thus, large, brilliant signals are present in the short arm of chromosome 3Xcr and the long arms of chromosomes 6Xcr and 7Xcr. Significant changes in the pAs1 and Fat-labelling patterns seen on chromosomes 2D1 and 5D1 and the emergence of large clusters of Fat element on some Xcr genome chromosomes are probably caused by species-specific translocations with the D genome chromosomes. At the same time, intergenomic translocations cannot explain the overall increase of labelling intensity of the Xcr genome chromosomes. This could indicate that the Fat element expanded over the course of Ae. crassa evolution from the D1 to the Xcr genome.
The tetraploid Ae. crassa is the parental form of three hexaploid species—Ae. crassa (genome D1XcrD2), Aegilops vavilovii (genome D1XcrS) and Aegilops juvenalis (genome D1XcrU). FISH analysis reveals that labelling patterns of the hexaploid Ae. crassa (Fig. 1i) and Ae. vavilovii chromosomes are the same to what is seen in the parental species. In contrast, we found significant alterations in the distribution of the Fat probe on the chromosomes of all three genomes of Ae. juvenalis (Fig. 1g; Supplementary material 2b, g, i), some of which are likely to be due to chromosomal rearrangements that have occurred over the course of evolution (Badaeva et al. 2002).
Polyploid Aegilops of the U genome cluster
This group of polyploid Aegilops includes seven tetraploid and one hexaploid species, all of which possess the U genome in combination with the C (Aegilops triuncialis), S (Aegilops peregrina, Aegilops kotschyi), M (Aegilops biuncialis, Aegilops geniculata) or X genome (Aegilops columnaris, Aegilops neglecta 4x (=Aegilops triaristata), Ae. neglecta 6x (=Aegilops recta)). The chromosomes in this group all exhibit moderate, diffuse hybridisation patterns with the Fat probe. Irrespective of the genome constitution of these polyploid species, all of the chromosomes in their karyotypes have similar fluorescence intensities. However, in some species, such as Ae. biuncialis (Fig. 1h), Ae. geniculata (Fig. 1e), Ae. columnaris (Fig. 1j), Ae. neglecta 6x (Fig. 1k), Ae. kotschyi (Fig. 1l) and Ae. peregrina, we did find one to several clusters of Fat probe hybridisation that permitted the identification of at least a few chromosomes in their karyotypes (Supplementary material 1e, h, j, k, l).
Other cereals
We carried out our FISH analysis with the Fat probe on two rye species (Secale cereale and Secale montanum), on hexaploid Triticale, four species of barley (Hordeum vulgare, Hordeum spontaneum, Hordeum geniculatum and Hordeum chilense), three oats with different genome constitutions (Avena prostrata, Avena ventricosa and Avena magna), Elymus dahuricus and Agropyron glaucum (Table 1). No hybridisation with the Fat probe is detected in the genomes of any of the three oat species or in two of the barley species, H. spontaneum and H. vulgare (I genome). In two other barley species, the diploid H. chilense (H genome) and the tetraploid H. geniculatum (XX1 genome), very weak signals are observed (Fig. 2m, p). Distinct hybridisation with the Fat probe is detected in all of the other Triticeae species that we investigated. All chromosomes of S. cereale and S. montanum and the R genome chromosomes in the karyotype of Triticale are labelled in their proximal region, and chromosomes 1R, 4R, 6R and 7R have additional chromosome-specific clusters of Fat probe hybridisation (Fig. 2l, n). In the hexaploid wheatgrass species A. glaucum, 14 chromosomes are heavily labelled, four other chromosomes possess small distinct clusters of Fat probe in pericentromeric regions, and the remaining 24 chromosomes hybridise poorly with this probe (Fig. 1f). In contrast, only eight of 42 chromosomes of E. dahuricus exhibit weak, diffuse Fat signals in their proximal regions (Fig. 1c; Supplementary material 1c, f; 2l, m, n, p).
Discussion
The results we present here demonstrate that the Fat element is a Triticeae-specific sequence. It is absent from the Aveneae species and appears to have arisen first in barley species (H. chilense and H. geniculatum) among the cereal genomes that we investigated in this study. In these barleys, we observe very weak, scarce signals distributed not specifically along all their chromosomes. In two other Hordeum species—H. vulgare and H. spontaneum, no hybridisation with the Fat probe have been detected. At the same time, in all of the other Triticeae species, we observe distinct hybridisation with the Fat probe. Based on these results, it seems likely that this sequence may have first appeared in the genomes of cereal species at the time of divergence of the genus Hordeum, estimated to be ~11 MYA (Huang et al. 2002). During subsequent evolution, the amount and distribution of Fat element changed in a species-specific and genome-specific manner.
The genus Secale is probably diverged from the common ancestor of the Triticeae next after barley (Flavell et al. 1977; Huang et al. 2002). In two rye species, S. montanum and S. cereale, we observe clear, dispersed labelling and several brilliant, clustered signals in chromosome-specific positions. These results indicate that evolution of rye was accompanied by amplification of the Fat sequence. Noteworthy, hybridisation with the Fat probe is completely absent from the regions of telomeric heterochromarin that are known to be formed by the arrays of tandemly repeated DNA families, some of which are found only in rye (Bedbrook et al. 1980; Jones and Flavell 1982b; Vershinin et al. 1995; Cuadrado and Jouve 2002). Based on these observations, we assume that amplification of the Fat sequence occurred independently of the emergence and amplification of the rye-specific repetitive DNAs.
Two other genera of the Triticeae that are Elymus and Agropyron diverged from the common ancestor of cereals prior to wheat/Aegilops complex. These two genera include a broad range of diploid and polyploid, annual and perennial species with different genome compositions (Dewey 1984). In our study, we analysed two hexaploid species representing each of these genera. FISH analysis reveals sharp differences between E. dahuricus and A. glaucum in hybridisation patterns of the Fat sequence on their chromosomes. Fourteen of 42 wheatgrass chromosomes are intensely labelled with the Fat probe and four other chromosomes possess small, but clear signals in their pericentromeric regions. This pattern is similar to what was obtained earlier in A. glaucum using FISH with the genomic DNA of Pseudoroegneria strigosa as a probe (Chen et al. 1998). Based in this observation, we suggest that these are the St genome chromosomes that are labelled with the Fat sequence, and the genome of the putative diploid progenitor of A. glaucum may possess a high amount of the Fat element. On the other hand, a diploid species from the Pseudoroegneria genus is thought to be the St genome to the polyploid Elymus species (Sun et al. 2007). However, only little amount of hybridisation with the Fat probe is detected on a few chromosomes of E. dahuricus. Clear differences in the amount of Fat probe hybridisation between the related genomes in karyotypes of the two hexaploid species can be due to different mechanisms: (1) the ancestral Pseudoroegneria forms that were the St genome donors to Agropyron and Elymus differ substantially in the amount of Fat element or (2) the Fat element was amplified in the St genome of A. glaucum over the course of polyploidisation.
The Fat sequence is found in all species of Triticum and Aegilops; however, it is most abundant in the D genome. Therefore, massive amplification of the Fat element may have occurred in the ancestral form of Ae. tauschii. In other species of Aegilops, the amount of Fat element present is probably similar to the level that was present in the ancient progenitor of Aegilops and wheat or increased slightly or decreased. It is of interest that the fluorescence intensity of Fat probe hybridisation is always higher in those Aegilops species that also show stronger hybridisation with the pAs1 probe. Thus, amplification of the Fat element over the course of the evolution of the Aegilops species is probably connected to amplification of the pAs1 sequence.
Evolution of Ae. comosa and Ae. uniaristata belonging to the Comopyron section was most likely accompanied by an increase in the amount of the Fat element, as their genomes show a relatively high level of hybridisation with the Fat probe. All diploid Aegilops species of the Sitopsis group have similar distribution pattern of the Fat probe; however, lower fluorescence intensities on the chromosomes of more ‘advanced’ species, such as Ae. longissima and Ae. sharonensis, compared with Ae. bicornis that is considered to be more ‘primitive’ (Dvořák and Zhang 1992) is probably indicative of gradual elimination of the Fat sequence in the course of divergence of this group. Diploid wheat species all have the same amount and distribution of the Fat element which is probably similar to what was present in the genome of the putative progenitor of Triticum/Aegilops complex.
Formation of polyploid Triticum and Aegilops species can also lead to alterations in the amount and distribution of the Fat element in one or two parental genomes; however, it is a gradual process that can probably be affected by the genome composition of a polyploid. In this study, we have not revealed any changes in the labelling patterns of parental genomes in such species as common wheat and T. timopheevii, Ae. cylindrica, Ae. triuncialis, Ae. peregrina and Ae. kotschyi, Ae. recta, hexaploid Ae. crassa and Ae. vavilovii, all of which are considered as ‘recent’ polyploids (Zhang et al. 1992; Dubkovsky and Dvořák 1994, 1995; Linc et al. 1999; Feldman 2001; Huang et al. 2002). Only little modifications are found in tetraploid emmer wheat and Ae. ventricosa. The origin of tetraploid wheat is dated back to ~0.5 MYA (Huang et al. 2002) and, although the age of Ae. ventricosa has not been determined, some modifications of the parental genomes detected in this species using different approaches suggest that it is not a recent polyploid (Dubkovsky and Dvořák 1994; Bardsley et al. 1999; Badaeva et al. 2002, 2008). By contrast, the significant changes are found in the tetraploid Ae. crassa, the most ancient Aegilops species (Dubkovsky and Dvořák 1995). Hybridisation of the Fat probe to the Xcr genome chromosomes is significantly higher than in any of the diploid species of the Sitopsis group; what is more, some chromosomes also possess brilliant clusters of the Fat probe hybridisation which are not found in genomes of the putative progenitor species. An increase in the amount of the Fat element in the Xcr genome of Ae. crassa can be caused by the expansion of the Fat sequence from the D to the Xcr genome and its local amplification in certain sites of the Xcr genome chromosomes over the course of long-term co-evolution of two sharply different genomes in a single nucleus.
The significant changes of the Fat probe labelling patterns are also found in some other Aegilops species with the UM (Ae. geniculata and Ae. biuncialis) or UX genome composition (Ae. columnaris and Ae. neglecta) and in hexaploid Ae. juvenalis with the D1XcrU type of nuclear genome. Although some of these changes are likely to be due to chromosomal rearrangements identified earlier in these species (Friebe et al. 1999; Badaeva et al. 2002, 2004; Schneider et al. 2008), the emergence of sharp, brilliant clusters of the Fat probe hybridisation suggests that the formation of these polyploids has promoted local amplification of the Fat sequence in one or several chromosomal sites. As the significant reorganisations of parental genomes in the abovementioned Aegilops species at the molecular and chromosomal levels are also shown by other authors (Abubakar and Kimber 1982; Kimber et al. 1988; Yen and Kimber 1990, 1992; Dubkovsky and Dvořák 1995; Resta et al. 1996), we assume that the alterations of the Fat probe distribution that we observed in our study are probably one of the constituents of these processes. Because all these polyploids share the same genome inherited from diploid Ae. umbellulata, it could be the U genome that promotes the processes of genome changes in the polyploids.
The tandem organisation of the Fat element was first hypothesised on the basis of analysis of 11 Mb of random BES from wheat chromosome 3B (Paux et al. 2006). However, according to FISH analysis, the distribution of the Fat probe is predominantly dispersed within proximal regions of chromosomes. Most families of tandem repeats that have been described so far are usually located in subtelomeric, pericentromeric or interstitial chromosome regions, and in FISH experiments, these elements appear as sharp, distinct signals on chromosomes (Badaeva et al. 1996; Heslop-Harrison 2000; Sharma and Raina 2005). Dispersed labelling is usually seen for mobile elements, in particular LTR retrotransposons (Katsiotis et al. 1996; Belyayev et al. 2001, 2005). Some of these elements can also form clusters in certain chromosome regions (Schmidt et al. 1995; Staton et al. 2009; Thomas et al. 2009). However, the Fat element exhibits no homology to any known retrotransposon families (Paux et al. 2006). Moreover, the labelling intensity that is characteristic of some of the retrotransposon families that have been mapped is usually much weaker than the hybridisation pattern that we see for the Fat element.
Construction of a BAC-based integrated physical map of the largest (995 Mb) wheat chromosome 3B (Paux et al. 2008) also revealed complicated genome organisation of the Fat sequence. Thus, in the BAC clone TAACSPALLhA_1119J14 (accession number AC216572), which is 131 kb long, 264 Fat repeat copies, 465 bp each form long tandems spanning over 129,880 bp. Several other BAC clones are found to carry more than 10 tandemly organised Fat repeats and, in most BACs, two or three copies of the Fat element are interspersed within several kilobase-long stretches of other repetitive sequences. Taking together our molecular and FISH results, we can conclude that, in genomes of cereal species, the Fat element is organised into tandem arrays varying in length (from few to several hundred copies) that are interspersed with arrays of other types of repetitive sequences. Thus, the Fat element represents a new family of Triticeae-specific repeated DNA elements exhibiting a clustered–dispersed distribution and preferential localisation to proximal chromosome regions. Though no classes of repetitive DNA with similarity to Fat have been found in the Triticeae (Sharma and Raina 2005), they have been identified in other plants. In particular, two families of satellite DNA with dispersed distribution have been identified in the Aveneae: pAm1, specific to the C genome (Solano et al. 1992), and pAs120a, specific to the A genome (Irigoyen et al. 2001).
In our analyses, we found that chromosomes of the D genome of Ae. tauschii and the polyploid species that are derived from it and chromosomes of some other species of Aegilops, Secale and Agropyron exhibit rather conservative and specific Fat probe hybridisation. Therefore, the Fat element, like other families of repetitive sequences that have been cloned, can be used in FISH analysis for chromosome identification and investigation of evolutionary processes at the chromosomal level.
Abbreviations
- BAC:
-
bacterial artificial chromosomes
- dUTP:
-
2’-dezoxyuridine-5'-triphosphate
- FISH:
-
Fluorescence in situ hybridization
- LTR:
-
long terminal repeat
- PCR:
-
polymerase chain reaction
References
Abubakar M, Kimber G (1982) Chromosome-pairing regulators in the former genus Aegilops. Z Pflanzenzucht 89:130–138
Ananiev EV, Vales MI, Phillips RL, Rines HW (2002) Isolation of A/D and C genome specific dispersed and clustered repetitive DNA sequences from Avena sativa. Genome 45:431–441
Badaeva ED, Friebe B, Gill BS (1996) Genome differentiation in Aegilops. 1. Distribution of highly repetitive DNA sequences on chromosomes of diploid species. Genome 39:293–306
Badaeva ED, Amosova AV, Muravenko OV et al (2002) Genome differentiation in Aegilops. 3. Evolution of the D-genome cluster. Plant Syst Evol 231:163–190
Badaeva ED, Amosova AV, Samatadze TE et al (2004) Genome differentiation in Aegilops. 4. Evolution of the U-genome cluster. Plant Syst Evol 246:45–76
Badaeva E, Dedkova O, Koenig J, Bernard S, Bernard M (2008) Analysis of introgression of Aegilops ventricosa Tausch. genetic material in a common wheat background using C-banding. Theor Appl Genet 117:803–811
Bardsley D, Cuadrado A, Jack P et al (1999) Chromosome markers in the tetraploid wheat Aegilops ventricosa analysed by in situ hybridization. Theor Appl Genet 99:300–304
Bedbrook RJ, Jones J, O'Dell M, Thompson RJ, Flavell RB (1980) A molecular description of telomeric heterochromatin in Secale species. Cell 19:545–560
Belyayev A, Raskina O, Nevo E (2001) Chromosomal distribution of reverse transcriptase-containing retroelements in two Triticeae species. Chromosome Res 9:129–136
Belyayev A, Raskina O, Nevo E (2005) Variability of the chromosomal distribution of Ty3-gypsy retrotransposons in the populations of two wild Triticeae species. Cytogenet Genome Res 109:43–49
Cabrera A, Friebe B, Jiang J, Gill BS (1995) Characterization of Hordeum chilense chromosomes by C-banding and in situ hybridization using highly repeated DNA probes. Genome 38:435–442
Charles M, Belcram H, Just J et al (2008) Dynamics and differential proliferation of transposable elements during the evolution of the B and A genomes of wheat. Genetics 180:1071–1086
Chen Q, Conner RL, Laroche A, Thomas JB (1998) Genome analysis of Thinopyrum intermedium and Thinoryrum ponticum using genomic in situ hybridization. Genome 41:580–586
Cuadrado A, Jouve N (2002) Evolutionary trends of different repetitive DNA sequences during speciation in the genus Secale. J Hered 93:339–345
Dewey DR (1984) The genomic system of classification as a guide to intergeneric hybridization with the perennial Triticeae. In: Gustafson JP (ed) Gene manipulation in plant improvement. Plenum, New York, pp 209–280
Dubkovsky J, Dvořák J (1994) Genome origin of Triticum cylindricum, Triticum triunciale, and Triticum ventricosum (Poaceae) inferred from variation in repeated nucleotide sequences: A methodological study. Am J Bot 81:1327–1335
Dubkovsky J, Dvořák J (1995) Genome identification of the Triticum crassum complex (Poaceae) with the restriction patterns of repeated nucleotide sequences. Am J Bot 182:131–140
Dvořák J, Dubkovsky J (1996) Genome analysis of polyploid species employing variation in repeated nucleotide sequences. In: Jauhar PP (ed) Methods in genome analysis in plants. CRC, Boca Raton, pp 133–145
Dvořák J, Zhang HB (1992) Reconstruction of the phylogeny of the genus Triticum from variation in repeated nucleotide sequences. Theor Appl Genet 84:419–429
Feldman M (2001) Origin of cultivated wheat. In: Bonjean AP, Angus WJ (eds) The world wheat book: a history of wheat breeding. Intersept, London, pp 3–56
Flavell RB (1986) Repetitive DNA and chromosome evolution in plants. Philos Trans R Soc Lond B Biol Sci 312:227–242
Flavell RB, Bennett MD, Smith JB, Smith DB (1974) Genome size and the proportion of repeated sequence DNA in plants. Biochem Genet 12:257–269
Flavell RB, Rimpau J, Smith DB (1977) Repeated sequence DNA relationships in four cereal genomes. Chromosoma 63:205–222
Flavell R, O'Dell M, Smith D (1979) Repeated sequence DNA comparisons between Triticum and Aegilops species. Heredity 42:309–322
Friebe BR, Tuleen NA, Gill BS (1999) Development and identification of a complete set of Triticum aestivum–Aegilops geniculata chromosome addition lines. Genome 42:374–380
Gerlach WL, Bedbrook JR (1979) Cloning and characterization of ribosomal RNA genes from wheat and barley. Nucleic Acids Res 7:1869–1885
Gerlach WL, Dyer TA (1980) Sequence organization of the repeated units in the nucleus of wheat which contains 5S-rRNA genes. Nucleic Acids Res 8:4851–4865
Grewal SIS, Moazed D (2003) Heterochromatin and epigenetic control of gene expression. Science 301:798–802
Heslop-Harrison JS (2000) Comparative genome organization in plants: from sequence and markers to chromatin and chromosomes. Plant Cell 12:617–636
Huang S, Sirikhachornkit A, Su X et al (2002) Genes encoding plastid acetyl-CoA carboxylase and 3-phosphoglycerate kinase of the Triticum/Aegilops complex and the evolutionary history of polyploid wheat. Proc Natl Acad Sci USA 99:8133–8138
Irigoyen ML, Loarce Y, Linares C et al (2001) Discrimination of the closely related A and B genomes in AABB tetraploid species of Avena. Theor Appl Genet 103:1160–1166
Jaillon O, Aury J-M, Noel B et al (2007) The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 449:463–467
Jiang J, Gill BS, Wang G, Ronald PC, Ward DC (1995) Metaphase and interphase fluorescence in situ hybridization mapping of the rice genome with bacterial artificial chromosomes. Proc Natl Acad Sci USA 92:4487–4491
Jones JDG, Flavell RB (1982a) The mapping of highly-repeated DNA families and their relationship to C-bands in chromosomes of Secale cereale. Chromosoma 86:595–612
Jones JDG, Flavell RB (1982b) The structure, amount and chromosomal localisation of defined repeated DNA sequences in species of the genus Secale. Chromosoma 86:613–641
Katsiotis A, Schmidt T, Heslop-Harrison JS (1996) Chromosomal and genomic organization of Ty1-copia-like retrotransposon sequences in the genus Avena. Genome 39:410–417
Kimber G, Sallee PJ, Feiner MM (1988) The interspecific and evolutionary relationships of Triticum ovatum. Genome 30:218–221
Li W, Zhang P, Fellers JP, Friebe B, Gill BS (2004) Sequence composition, organization, and evolution of the core Triticeae genome. Plant J 40:500–511
Lim KY, Matyasek R, Kovarik A, Fulnecek J, Leitch AR (2005) Molecular cytogenetics and tandem repeat sequence evolution in the allopolyploid Nicotiana rustica compared with diploid progenitors N. paniculata and N. undulata. Cytogenet Genome Res 109:298–309
Linares C, Loarce Y, Serna AAF (2001) Isolation and characterization of two novel retrotransposons of the Ty1-copia group in oat genomes. Chromosoma 110:115–123
Linc G, Friebe BR, Kynast RG et al (1999) Molecular cytogenetic analysis of Aegilops cylindrica Host. Genome 42:497–503
Nagaki K, Kishii M, Tsujimoto H, Sasakuma T (1999) Tandem repetitive Afa-family sequences from Leymus racemosa and Psathyrostachys juncea (Poaceae). Genome 42:1258–1260
Paterson AH, Bowers JE, Bruggmann R et al (2009) The Sorghum bicolor genome and the diversification of grasses. Nature 457:551–556
Paux E, Roger D, Badaeva E et al (2006) Characterizing the composition and evolution of homoeologous genomes in hexaploid wheat through BAC-end sequencing on chromosome 3B. Plant J 48:463–474
Paux E, Sourdille P, Salse J et al (2008) A physical map of the 1-gigabase bread wheat chromosome 3B. Science 322:101–104
Rayburn AL, Gill BS (1986) Isolation of a D-genome specific repeated DNA sequence from Aegilops squarrosa. Plant Mol Biol Rep 4:104–109
Resta P, Zhang HB, Dubkovsky J, Dvořák J (1996) The origin of the genomes of Triticum biunciale, T. ovatum, T. neglectum, T. columnare, and T. rectum based on variation in repeated nucleotide sequences. Am J Bot 83:1556–1565
Schmidt T, Kubis S, Heslop-Harrison JS (1995) Analysis and chromosomal localization of retrotransposons in sugar beet (Beta vulgaris L.): LINEs and Ty1-copia-like elements as major components of the genome. Chromosome Res 3:335–345
Schnable PS, Ware D, Fulton RS et al (2009) The B73 maize genome: complexity, diversity, and dynamics. Science 326:1112–1115
Schneider A, Linc G, Molnar-Lang M (2003) Fluorescence in situ hybridization polymorphism using two repetitive DNA clones in different cultivars of wheat. Plant Breed 122:396–400
Schneider A, Molnár I, Molnár-Láng M (2008) Incorporation of Aegilops biuncialis chromosomes into wheat and their identification using fluorescent in situ hybridization. Acta Biol Szeged 52:133–137
Sequencing Project International Rice G (2005) The map-based sequence of the rice genome. Nature 436:793–800
Sharma S, Raina SN (2005) Organization and evolution of highly repeated satellite DNA sequences in plant chromosomes. Cytogenet Genome Res 109:15–26
Solano R, Hueros G, Fominaya A, Ferrer E (1992) Organization of repeated sequences in species of the genus Avena. Theor Appl Genet 83:602–607
Staton SE, Ungerer MC, Moore RC (2009) The genomic organization of Ty3/gypsy-like retrotransposons in Helianthus (Asteraceae) homoploid hybrid species. Am J Bot 96:1646–1655
Sun GL, Daley T, Ni Y (2007) Molecular evolution and genome divergence at RPB2 gene of the St and H genomes in Elymus species. Plant Mol Biol 64:645–655
Taketa S, Ando H, Takeda K, Harrison GE, Heslop-Harrison JS (2000) The distribution, organization and evolution of two abundant and widespread repetitive DNA sequences in the genus Hordeum. Theor Appl Genet 100:169–176
The Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature 408:796–815
Thomas W, Stefan T, Andreas H et al (2009) A whole-genome snapshot of 454 sequences exposes the composition of the barley genome and provides evidence for parallel evolution of genome size in wheat and barley. Plant J 59:712–722
Vershinin AV, Schwarzacher T, Heslop-Harrison JS (1995) The large-scale genomic organization of repetitive DNA families at the telomeres of rye chromosomes. Plant Cell 7:1823–1833
Yen Y, Kimber G (1990) The U genome in Triticum ovatum from Turkey. Cereal Res Commun 18:13–19
Yen Y, Kimber G (1992) Genomic relationships on N-genome Triticum species. Genome 35:962–966
Zhang HB, Dvorak J (1992) The genome origin and evolution of hexaploid Triticum crassum and Triticum syriacum determined from variation in repeated nucleotide sequences. Genome 35:806–814
Zhang H-B, Dvořák J, Waines JG (1992) Diploid ancestry and evolution of Triticum kotschyi and T. peregrinum examined using variation in repeated nucleotide sequences. Genome 35:182–191
Zhang P, Li W, Fellers J, Friebe B, Gill BS (2004) BAC-FISH in wheat identifies chromosome landmarks consisting of different types of transposable elements. Chromosoma 112:288–299
Acknowledgements
The authors thank Drs. N.N. Chikida and I.G. Loskutov (Vavilov Institute of Plant Industry, VIR, St. Petersburg, Russia); Dr. B.S. Gill (Wheat Genetic and Genomics Resource Center, Kansas State University, USA); Dr. J. Konopka (ICARDA, Aleppo, Syria); Dr. S.N. Sibikeev (Agricultural Research Institute for South-East Region, Saratov, Russia); Drs. V.A. Pukhalskyi and V.P. Upelniek (Vavilov Institute of General Genetics, Moscow, Russia); Dr. S.V. Chebotar (Plant Breeding Institute, Odessa, Ukraine); Dr. L.A. Pershina (Institute of Cytology and Genetics, Novosibirsk, Russia); Dr. I.V. Iordanskaya (Scientific-Research Agricultural Institute of the Central Zone of Non-Chernozem Region, Moscow Region, Nemchinovka); and Dr. Hernandez P. (Institute for Sustainable Agriculture, Cordoba, Spain). This work was partially supported by grant of the Russian State Foundation for Basic Research (project number 08-04-00302).
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Pat Heslop-Harrison.


{kind=link}
{kind=link}
Rights and permissions
About this article
Cite this article
Badaeva, E.D., Zoshchuk, S.A., Paux, E. et al. Fat element—a new marker for chromosome and genome analysis in the Triticeae. Chromosome Res 18, 697–709 (2010). https://doi.org/10.1007/s10577-010-9151-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10577-010-9151-x