
Abstract
Parkinson’s disease is an incurable progressive neurological condition caused by a degeneration of dopamine-producing cells characterized by motor and non-motor symptoms. The major mechanisms of the antiepileptic actions of ZNS are inhibition of voltage-gated Na+ channel, T-type voltage-sensitive Ca2+ channel, Ca2+-induced Ca2+ releasing system, and neuronal depolarization-induced glutamate release; and enhancement of release of inhibitory neurotransmitters; however, the detailed mechanism of antiparkinsonian effects of ZNS remains to be clarified. We aimed to investigate to the effect of ZNS on the oxidative stress, cell viability, Ca2+ signaling, and caspase activity that induced by the MPP+ model of Parkinson’s in neuronal PC12 cells. Neuronal PC12 cells were divided into four groups namely, control, ZNS, MPP+, and ZNS+MPP+ groups. The dose and duration of ZNS and MPP+ were determined according to cell viability (MTT) analysis which used to assess the cell viability. The cells in ZNS, MPP+, and ZNS+MPP+ groups were incubated for 5 h with 100 μM ZNS, 10 h with 100 μM MPP+, and 10 h with ZNS and MPP+, respectively. Lipid peroxidation and cytosolic free Ca2+ concentrations were higher in the MPP+ group than in control although their levels were lower in ZNS and the ZNS+MPP+ groups than in control. Reduced glutathione and glutathione peroxidase values were lower in the MPP+ group although they were higher in the ZNS and the ZNS+MPP+ groups than in control. Caspase-3 activity was lower in the ZNS group than in the MPP+ group. In conclusion, ZNS induced modulator effects on the oxidative stress, intracellular Ca2+, and the caspase-3 values in an experimental model of Parkinson disease.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Parkinson’s disease is a common progressive neurodegenerative condition associated with significant disability and negative impact on quality of life. Although the cause of Parkinson’s disease is unknown, the pathologic manifestation involves the loss or dysfunction of dopaminergic neurons in the substantia nigra pars compacta (Lew 2007). A large part of the research is focused on the neurodegeneration which plays an important role in the pathogenesis of Parkinson’s disease (Lew 2007; Surmeier et al. 2011). In the pathogenesis of neurodegenerative diseases, there is available information that it may be factor of oxidative stress, excitotoxic mechanisms, mitochondrial energy metabolism, apoptosis, and intracellular Ca2+ balance disorders (Nazıroğlu 2007; Surmeier et al. 2011; Schapira 2011). Oxidative damage in DNA, proteins, and lipids, decrease in glutathione (GSH), and increase in iron was shown on postmortem brain studies of Parkinson’s patients (Baillet et al. 2010; Müller and Muhlack 2011).
In the substantia nigra of Parkinson’s patients, nearly 30–40 % selective reduction was found in the mitochondrial respiratory chain complex I activity (Schapira 2011). Mitochondrial complex I defects contribute to the cell degeneration via the reduction the synthesis of ATP. 1-Methyl-4-phenylpyridinium ion (MPP+)-mediated selective damage to dopaminergic neurons of the nitrostrial damage has been widely used as a model of Parkinson’s disease (Korff et al. 2011). In the synaptosomes of rat brain, complex I inhibition by the MPP+ or MPTP may result in reduced cellular ATP. In addition, MPP+ may induce release of dopamine, which in turn could generate free oxygen radicals upon auto-oxidation (Hasegawa et al. 1990). In vitro evidence for reactive oxygen species (ROS) involvement during MPP+ toxicity has been mostly controversial, with several reports supporting either side of the discussion.
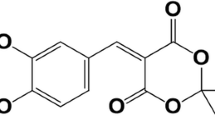
Zonisamide (ZNS, 1,2-benzisoxazole-3-methanesulphonamide) has been developed as anticonvulsant agent in Japan in the 1970s (Okada et al. 2002). Interestingly, several clinical studies have reported the wide clinical spectrum of ZNS against both psychiatric and neurological disorders, including epilepsy mood disorder, essential tremor, and Parkinson’s disease, and protecting against ischemic cerebral damage, such as stroke. The major mechanisms of the antiepileptic actions of ZNS are inhibition of voltage-gated Na+ channel, T-type voltage-sensitive Ca2+ channel, Ca2+ releasing system, and neuronal depolarization-induced glutamate release; enhancement of release of inhibitory neurotransmitters, i.e., GABA, dopamine, and serotonin; and lack of affinity to GABAA receptor (Okada et al. 2002; Zhu et al. 2002). A recent randomized control study demonstrated also that ZNS is effective in Parkinson’s disease at the lower than the therapeutic doses against epilepsy (25–50 mg/day); however, the detailed mechanism of antiparkinsonian effects of ZNS on the molecular pathways remains to be clarified (Mohammadianinejad et al. 2011).
Rat pheochromocytoma (PC12) cells have been used extensively as an experimental system to study various aspects of dopaminergic neurons (Altinkiliç et al. 2010). Although these cells are not true brain dopaminergic neurons, PC12 cells are able to produce dopamine and express dopamine transporters (Kadota et al. 1996). Upon nerve growth factor (NGF) stimulation, PC12 cells not only display abundant neuritic growth, but also adopt a neurochemical dopaminergic phenotype (Fonck and Baudry 2001).
In the current study, we investigated the effect of ZNS on the oxidative stress, Ca2+ signaling, and caspase activity in the experimental model of Parkinson disease that is induced by the MPP+ in neuronal PC12 cells.
Materials and Methods
Chemicals
Cell culture flasks were bought from TPP Company (Switzerland). All chemicals (fish oil, cumene hydroperoxide, KOH, NaOH, thiobarbutiric acid, 1,1,3,3-tetraethoxypropane, 5,5-dithiobis-2 nitrobenzoik, tris-hydroxymethil-aminomethan, 5,5-dithiobis-2 nitrobenzoik asit, cumene hydroperoxide, glutathione, and butylhydroxytoluol) and Caspase-3 (Acetyl-ASp-Glu-Val-Asp-t..amide-4-methylcoumin, “with” catolog no. A-1086) commercial kit were obtained from Sigma-Aldrich Chemical Inc. (St. Louis, MO, USA); and all organic solvents (n-hexane and ethyl alcohol) and RPMI 1640 medium were purchased from Merck Chemicals (Darmstadt, Germany). Fura-2 acetoxymethyl ester (Fura-2/AM) was purchased from Promega Inc (USA). All reagents were analytical grade. All reagents except the phosphate buffers were prepared daily and stored at +4 °C. The reagents were equilibrated at room temperature for half an hour before an analysis was initiated or reagent containers were refilled. Phosphate buffers were stable at +4 °C for 1 month.
Cell Culture
PC12, a rat pheochromocytoma-derived cell line was selected as the neuronal model. The cell line was originally obtained from DSMZ Cell Lines Bank (Germany). Before treatment, PC12 cells were placed in poly-d-lysine-coated cell culture flask at a density of 1 × 105/cm2 and were allowed to attach for 24 h in RPMI 1640 medium supplemented with 10 % horse serum, 5 % fetal bovine serum, 4 mM glutamine, and a mixture of 1 % of penicillin/streptomycin/l-glutamine. Cells were incubated at 37 °C in humid, 5 % CO2, and 95 % air environment (Altinkiliç et al. 2010). Differentiation was induced by addition of (NGF) to the cultures (50 ng/ml) for 2 days.
Study Groups
The PC12 cells were divided into four groups as follows:
-
Group 1
Control group: The cells were incubated with only RPMI medium for 24 h.
-
Group 2
ZNS group: The cells in the group were incubated with ZNS (100 μM) for 5 h (Matar et al. 2009).
-
Group 3
MPP+ group: The cells in the group were incubated with 100 μM MPP+ for 10 h.
-
Group 4
ZNS+MPP+ group: The cells in the group were incubated ZNS (100 μM) for 5 h before MPP+ (100 μM and 10 h) incubation.
At the end of the treatments, half of the cells were washed by the medium and stored at −33 °C. LP and antioxidant analysis were performed in the cells within one week. Remaining cells were immediately used for cytosolic free calcium ion ([Ca2+] i ) concentration and caspase analysis. During the [Ca2+] i analysis, the eight groups were exposed to H2O2 for stimulating [Ca2+] i concentrations.
Measurement of Cytosolic Free Ca2+ ([Ca2+] i ) Concentration
Cells were loaded with fura-2 by incubation with 4 μM fura-2/AM for 30 min at room temperature according to a procedure published elsewhere (Uğuz et al. 2009). Once loaded, the cells were washed and gently re-suspended in Na-HEPES solution containing (in mM): NaCl, 140; KCl, 4.7; CaCl2, 1.2; MgCl2, 1.1; glucose, 10; and HEPES, 10 (pH 7.4). The four groups were exposed to H2O2 for stimulating ([Ca2+] i ) influx. Fluorescence was recorded from 2 ml aliquots of magnetically stirred cellular suspension (2 × 106 cells/ml) at 37 °C using a spectrofluorometer (Carry Eclipsys, Varian Inc, Sydney, Australia) with excitation wavelengths of 340 and 380 nm and emission at 505 nm. Changes in [Ca2+] i were monitored using the fura-2/AM 340/380 nm fluorescence ratio and were calibrated according to the method of Grynkiewicz et al. (1985).
Ca2+ concentrations were estimated using the integral of the rise in [Ca2+] i for 150 s after addition of H2O2 (Uğuz et al. 2009; Espino et al. 2009). Ca2+ release is expressed in nanomolar taking a sample every second (nM/second) as previously described (Heemskerk et al. 1997).
Lipid Peroxidation (LP) Determinations
LP levels in the PC12 cell lines were measured with the thiobarbituric acid reaction by the method of Placer et al. (1966). The quantification of thiobarbituric acid reactive substances was determined by comparing the absorption to the standard curve of malondialdehyde (MDA) equivalents generated by acid catalyzed hydrolysis of 1,1,3,3 tetramethoxypropane. The values of LP in the PC12 cells were expressed as μmol/g protein.
Reduced Glutathione (GSH), Glutathione Peroxidase (GSH-Px), and Protein Assay
The GSH content of the PC12 cells was measured at 412 nm by the method of Sedlak and Lindsay (1968) as described in our previous study (Kutluhan et al. 2009). GSH-Px activities of PC12 cells were measured spectrophotometrically at 37 °C and 412 nm according to (Lawrence and Burk 1976). The protein content in the PC12 cells was measured by method of Lowry et al. (1951) with bovine serum albumin as the standard.
Caspase-3 Assay
The Caspase-3 assay (Acetyl-ASp-Glu-Val-Asp-t..amide-4-methylcoumin, “with” (catolog no. A-1086) commercial kit) was performed according to the instructions provided by Sigma Inc.
Statistical Analysis
Data are expressed as means ± standard deviations (SD) of the numbers of determinations. Analysis of statistical significance was performed using the SPSS program (9.05). In order to compare the different treatments, statistical significance was calculated by Mann–Whitney U test analysis. p < 0.05 was considered to indicate a statistically significant difference.
Results
The Determination of the Toxic Dose of MPP+ ‘s on Cell Viability
In PC12 cells, the effect of MPP+ on cell viability is shown in Fig. 1. Cells were incubated with increasing concentrations of MPP+ (25 μM, 50 μM, 100 μM, 250 μM, 500 μM, 1 mM, 10 mM, 25 mM, 50 mM, 100 mM, 200 mM) seven different time (30 min, 1 h, 2 h, 5 h, 12 h, 24 h,48 h). The toxicity of MPP+ began at 100 mM dose after 24 h of incubation. The toxic dose of MPP+ is defined as 24 h at 100 μM in cell culture (Fig. 1).

The effects of MPP+ on cell viability (MTT) in different doses and times
The Determination of Therapeutic and Toxic Dose of ZNS on Cell Viability
The effect of ZNS on cell viability in PC12 cells is shown in Fig. 2. PC12 cells are incubated at 11 different doses (25 μM, 50 μM, 100 μM, 250 μM, 500 μM, 1 mM, 10 mM, 25 mM, 50 mM, 100 mM, and 200 mM) with ZNS seven different times (30 min, 1 h, 2 h, 5 h, 12 h, 24 h, and 48 h) for determination of toxic doses of ZNS. In the cell viability test, at 100 μM dose and within 10 h 25 % reduction was determined as non-toxic dose. It is determined that the toxic effect of ZNS started at 100 μM dose in 24 h. The highest dose of the therapeutic effect of ZNS is defined as 100 μM dose and 12 h (Fig. 2).

The effects of zonisamide on cell viability (MTT) in different doses and times
Effects of ZNS and MPP+ on LP Levels in PC12 Cells
Lipid peroxidation levels in four groups are shown in Table 1. The LP levels were significantly (p < 0.05) higher in MPP+ group as compared with the control group. The LP levels were also significantly (p < 0.05) lower in ZNS and ZNS+ MPP+ groups than in MPP+. In other words, it was observed that ZNS induced a protective effect on MPP+-induced LP in the PC12 neuronal cells (Table 1).
Effects of ZNS and MPP+ on GSH Levels and GSH-Px Activity in PC12 Cells
The GSH levels and GSH-Px activity in four groups are shown in Table 1. The GSH levels were significantly (p < 0.05) lower in the MPP+ group than in control group. The GSH levels were significantly (p < 0.01) higher in ZNS (p < 0.01 and p < 0.001) and ZNS+MPP+ (p < 0.01) groups than in MPP+ group.
Effects of ZNS and MPP+ on Cytosolic Free Calcium Ion ([Ca2+] i ) Concentrations in PC12 Cells
The effects on ZNS and MPP+ on the [Ca2+] i concentrations in neuronal PC12 cells in Fig. 3. According to control group, the concentrations of [Ca2+] i was found to be increased in MPP+ (p < 0.001) group. The concentrations of [Ca2+] i were significantly (p < 0.001) lower in ZNS and ZNS plus MPP+ groups than in control group. The concentrations of [Ca2+] i were found to be significantly (p < 0.001) lower in ZNS group than in MPP+ group. As compared to ZNS group, the concentrations of [Ca2+] i was found to be significantly (p < 0.01) higher in ZNS plus MPP+ group. In other words, ZNS induced modulator role on MPP+-induced Ca2+ influx in the neuronal PC12 cells (Fig. 3).

The effects of MPP+ and zonisamide (ZNS) on cytosolic Ca2+ [Ca2+] i concentrations in PC12 cell line (n = 8, mean ± SD). a p < 0.001 and versus control group, b p < 0.001 and versus MPP+ group, c p < 0.01 and versus ZNS group (Color figure online)
Effects of ZNS and MPP+ on Caspase-3 Activity in PC12 Cells
The effects on ZNS and MPP+ on caspase-3 activity in PC12 cells are shown in Fig. 4. In comparison with MPP+ group caspase-3 levels were found to be significantly (p < 0.001) lower in ZNS and ZNS plus MPP+ groups. As compared to ZNS group, caspase-3 activity was found to be significantly (p < 0.01) higher in the ZNS plus MPP+ group. As a result, it was observed that ZNS had a protective effect on PC12 cells by reducing caspase-3 activity which is a precursor of apoptosis (Fig. 4).

The effects of MPP+ and zonisamide (ZNS) on caspase-3 activity in neuronal PC12 cell line (n = 8, mean ± SD). a p < 0.001 and versus control group, b p < 0.001 and versus MPP+ group, c p < 0.01 and versus ZNS group
Discussion
Actual pathophysiological mechanism underlying the anti-parkinsonian effect of ZNS remains uncertain. We detected that, ZNS reduced LP in PC12 cells and increased the levels of GSH and GSH-Px. Therefore, we may assume that ZNS is an agent having antioxidant properties. Impaired Ca2+ homeostasis has an important role in neuronal degeneration. We found that ZNS also modulated the MPP+-induced Ca2+ over influx and caspase-3 activity in PC12 cells.
The brain is extremely susceptible to oxidative damage induced by reactive oxygen species (ROS) because it generates very high levels of ROS due to its very high aerobic metabolism and blood perfusion and has relatively poor enzymatic antioxidant defense (Naziroğlu et al. 2009). Many factors play a role in the development of Parkinson’s disease. An imbalance between oxidative stress and antioxidant defense mechanisms in the brain plays an important role in the pathogenesis of Parkinson’s disease. Changes in energy metabolism, excitotoxicity, and impaired Ca2+ homeostasis are the other important factors that may activate apoptosis with different stimuli (Naziroglu et al. 2012). The loss of dopaminergic neurons in Parkinson’s disease results in enhanced metabolism of dopamine, augmenting the formation of H2O2, thus leading to generation of highly neurotoxic hydroxyl radicals (Baillet et al. 2010). The generation of free radicals can also be produced by 6-hydroxydopamine or MPTP which destroys striatal dopaminergic neurons causing parkinsonism in experimental animals as well as human beings (Lee et al. 2012). Studies of the substantia nigra after death in Parkinson’s disease have suggested the presence of oxidative stress and depletion of GSH (Martin and Teismann 2009; Korff et al. 2011); a high level of total iron with reduced level of ferritin; and deficiency of mitochondrial complex I. The biochemical changes due to oxidative stress resulting from tissue iron overload (siderosis) are similar to those now being identified in parkinsonian substantia nigra. These include the reduction of mitochondrial electron transport, complex I and III activities, GSH-Px activity, GSH, ascorbate, calcium-binding protein, and superoxide dismutase and increase of basal LP and deposition of iron (Danielson et al. 2011; Naziroglu et al. 2012). In the current study we observed decreased values of GSH and GSH-Px in MPP+ exposed groups although LP and [Ca2+] i concentrations were increased in MPP+ group.
ZNS acts as a neuroprotectant against oxidative stress and progressive dopaminergic neurodegeneration (Asanuma et al. 2010). ZNS also inhibited monoamine oxidase B and dopaminergic oxidative stress (Binda et al. 2011). In the current study, ZNS increased GSH and GSH-Px values although LP levels were decreased by ZNS incubations. Smilarly, Pileblad et al. (1989) suggested that GSH has an important role in catecholaminergic neurons: protecting against the oxidation of endogenous catechols in rat striatum. Asanuma et al. (2010) reported that ZNS markedly increased GSH levels by enhancing the astroglial cystine transport system and/or astroglial proliferation via S100beta production or secretion. Komatsu et al. (2000) reported the inhibitor effect on the lipid perokside of ZNS and the reducing effect of 8-hydroksi-2V-deoksiguanosin (8-OHdG) level which is the sign of oxidative DNA damage. Dexter et al. (1994) reported that the levels of GSH in substantia nigra were reduced by 35 % in patients with incidental Lewy body disease compared to control subjects. Hence, GSH, GSH-Px, and LP results were supported by results of Dexter et al. (1994), Pileblad et al. (1989), Asanuma et al. (2010), and Komatsu et al. (2000).
The major mechanisms of ZNS are inhibition of voltage-gated Na+ channel, T-type voltage-sensitive Ca2+ channel, and Ca2+ releasing system (Okada et al. 2002; Zhu et al. 2002). The MPP+-induced [Ca2+] i concentrations were modulated by ZNS due to regulator role of ZNS on T-type voltage-sensitive Ca2+ channel, Ca2+ releasing system. Similarly, Kawata et al. (1999) reported that ZNS inhibited the depolarization induced by neuronal excitation, whereas ZNS might enhance the N-type Ca2+ channel activity or N-type Ca2+ channel-related exocytosis mechanisms. Suzuki et al. (1992) reported that ZNS against maximal electroshock seizures in mice reduced T-type Ca2+ current in a dose-dependent manner.
In conclusion, ZNS induced protective effect on MPP+-induced oxidative stress and antioxidant redox system through modulation of cytosolic Ca2+ concentrations in the neuronal PC12 cells. Our current study on PC12 cells taken together with in vitro studies with parkinsonism improved cognition with ZNS suggest a, therefore, unexploited therapeutic potential for this drug in neuronal diseases such as Parkinson and epilepsy diseases that are characterized by oxidant stress.
Abbreviations
- GSH:
-
Reduced glutathione
- GSH-Px:
-
Glutathione peroxidase
- LP:
-
Lipid peroxidation
- MPP+ :
-
1-Methyl-4-phenylpyridinium ion
- NGF:
-
Nerve growth factor
- PC12:
-
Rat pheochromocytoma-derived cell line
- ROS:
-
Reactive oxygen species
- ZNS:
-
Zonisamide
References
Altinkiliç S, Naziroğlu M, Uğuz AC, Ozcankaya R (2010) Fish oil and antipsychotic drug risperidone modulate oxidative stress in PC12 cell membranes through regulation of cytosolic calcium ion release and antioxidant system. J Membr Biol 235:211–218
Asanuma M, Miyazaki I, Diaz-Corrales FJ, Kimoto N, Kikkawa Y, Takeshima M, Miyoshi K, Murata M (2010) Neuroprotective effects of zonisamide target astrocyte. Ann Neurol 67:239–249
Baillet A, Chanteperdrix V, Trocmé C, Casez P, Garrel C, Besson G (2010) The role of oxidative stress in amyotrophic lateral sclerosis and Parkinson’s disease. Neurochem Res 35:1530–1537
Binda C, Aldeco M, Mattevi A, Edmondson DE (2011) Interactions of monoamine oxidases with the antiepileptic drug zonisamide: specificity of inhibition and structure of the human monoamine oxidase B complex. J Med Chem 54:909–912
Danielson SR, Held JM, Oo M, Riley R, Gibson BW, Andersen JK (2011) Quantitative mapping of reversible mitochondrial complex I cysteine oxidation in a Parkinson disease mouse model. J Biol Chem 286:7601–7608
Dexter DT, Sian J, Rose S, Hindmarsh JG, Mann VM, Cooper JM, Wells FR, Daniel SE, Lees AJ, Schapira AH et al (1994) Indices of oxidative stress and mitochondrial function in individuals with incidental Lewy body disease. Ann Neurol 35:38–44
Espino J, Mediero M, Bejarano I, Lozano GM, Ortiz A, García JF, Rodríguez AB, Pariente JA (2009) Reduced levels of intracellular calcium releasing in spermatozoa from asthenozoospermic patients. Rep Biol Endocrinol 7:11
Fonck C, Baudry M (2001) Toxic effects of MPP(+) and MPTP in PC12 cells independent of reactive oxygen species formation. Brain Res 905:199–206
Grynkiewicz C, Poenie M, Tsien RY (1985) A new generation of Ca2+ indicators with greatly improved fluorescence properties. J Biol Chem 260:3440–3450
Hasegawa E, Takeshige K, Oishi T, Murai Y, Minakami S (1990) 1-Methyl-4-phenylpyridinium (MPP+) induces NADH-dependent superoxide formation and enhances NADH-dependent lipid peroxidation in bovine heart submitochondrial particles. Biochem Biophys Res Commun 170:1049–1055
Heemskerk JW, Feijge MA, Henneman L, Rosing J, Hemker HC (1997) The Ca2+-mobilizing potency of alpha-thrombin and thrombin receptor-activating peptide on human platelets concentration and time effects of thrombin-induced Ca2+ signalling. Eur J Biochem 249:547–555
Kadota T, Yamaai T, Saito Y, Akita Y, Kawashima S, Moroi K, Inagaki N, Kadota K (1996) Expression of dopamine transporter at the tips of growing neurites of PC12 cells. J Histochem Cytochem 44:989–996
Kawata Y, Okada M, Murakami T, Mizuno K, Wada K, Kondo T, Kaneko S (1999) Effects of zonisamide on K+ and Ca2+ evoked release of monoamine as well as K+ evoked intracellular Ca2+ mobilization in rat hippocampus. Epilepsy Res 35:173–182
Komatsu M, Hiramatsu M, Willmore LJ (2000) Zonisamide reduces the increase in 8-hydroxy-2′-deoxyguanosine levels formed during iron-induced epileptogenesis in the brains of rats. Epilepsia 41:1091–1094
Korff A, Pfeiffer B, Smeyne M, Kocak M, Pfeiffer RF, Smeyne RJ (2011) Alterations in glutathione S-transferase pi expression following exposure to MPP+-induced oxidative stress in the blood of Parkinson’s disease patients. Parkinsonism Relat Disord 17:765–768
Kutluhan S, Naziroğlu M, Celik O, Yilmaz M (2009) Effects of selenium and topiramate on lipid peroxidation and antioxidant vitamin levels in blood of pentylentetrazol-induced epileptic rats. Biol Trace Elem Res 129:181–189
Lawrence RA, Burk RF (1976) Glutathione peroxidase activity in selenium-deficient rat liver. Biochem Biophys Res Commun 71:952–958
Lee KW, Zhao X, Im JY, Grosso H, Jang WH, Chan TW, Sonsalla PK, German DC, Ichijo H, Junn E, Mouradian MM (2012) Apoptosis signal-regulating kinase 1 mediates MPTP toxicity and regulates glial activation. PLoS One 7:e29935
Lew M (2007) Overview of Parkinson’s disease. Pharmacotherapy 27(12 Pt 2):155S–160S
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the Folin-Phenol reagent. J Biol Chem 193:265–275
Martin HL, Teismann P (2009) Glutathione—a review on its role and significance in Parkinson’s disease. FASEB J 23:3263–3272
Matar N, Jin W, Wrubel H, Hescheler J, Schneider T, Weiergräber M (2009) Zonisamide block of cloned human T-type voltage-gated calcium channels. Epilepsy Res 83:224–234
Mohammadianinejad SE, Abbasi V, Sajedi SA, Majdinasab N, Abdollahi F, Hajmanouchehri R, Faraji A (2011) Zonisamide versus topiramate in migraine prophylaxis: a double-blind randomized clinical trial. Clin Neuropharmacol 34:174–177
Müller T, Muhlack S (2011) Cysteinyl-glycine reduction as marker for levodopa-induced oxidative stress in Parkinson’s disease patients. Mov Disord 26:543–546
Nazıroğlu M (2007) Molecular mechanisms of vitamin E on intracellular signaling pathways in brain. In: Goth Laszlo (ed) Reactive oxygen species and diseases. Research Signpost, Trivandrum, pp 239–256
Naziroğlu M, Kutluhan S, Uğuz AC, Celik O, Bal R, Butterworth PJ (2009) Topiramate and vitamin E modulate the electroencephalographic records, brain microsomal and blood antioxidant redox system in pentylentetrazol-induced seizure of rats. J Membr Biol 229:131–140
Nazıroğlu M, Dikici DM, Dursun S (2012) Role of oxidative stress and Ca(2+) signaling on molecular pathways of neuropathic pain in diabetes: focus on TRP channels. Neurochem Res 37(10):2065–2075
Okada M, Zhu G, Yoshida S, Kanai K, Hirose S, Kaneko S (2002) Exocytosis mechanism as a new targeting site for mechanisms of action of antiepileptic drugs. Life Sci 72:465–473
Pileblad E, Magnusson T, Fornstedt B (1989) Reduction of brain glutathione by l-buthionine sulfoximine potentiates the dopamine-depleting action of 6-hydroxydopamine in rat striatum. J Neurochem 52:978–980
Placer ZA, Cushman L, Johnson BC (1966) Estimation of products of lipid peroxidation (malonyl dialdehyde) in biological fluids. Anal Biochem 16:359–364
Schapira AH (2011) Mitochondrial pathology in Parkinson’s disease. Mt Sinai J Med 78:872–881
Sedlak J, Lindsay RHC (1968) Estimation of total, protein bound and non-protein sulfhydryl groups in tissue with Ellmann’s reagent. Anal Biochem 25:192–205
Surmeier DJ, Guzman JN, Sanchez-Padilla J, Schumacker PT (2011) The role of calcium and mitochondrial oxidant stress in the loss of substantia nigra pars compacta dopaminergic neurons in Parkinson’s disease. Neuroscience 198:221–231
Suzuki S, Kawakami K, Nishimura S, Watanabe Y, Yagi K, Seino M, Miyamoto K (1992) Zonisamide blocks T-type calcium channel in cultured neurons of rat cerebral cortex. Epilepsy Res 12:21–27
Uğuz AC, Nazıroğlu M, Espino J, Bejarano I, González D, Rodríguez AB, Pariente JA (2009) Selenium modulates oxidative stress-induced cell apoptosis in human myeloid HL-60 cells via regulation of caspase-3,-9 and calcium influx. J Membr Biol 232:15–23
Zhu G, Okada M, Murakami T, Kawata Y, Kamata A, Kaneko S (2002) Interaction between carbamazepine, zonisamide and voltage-sensitive Ca2+ channel on acetylcholine release in rat frontal cortex. Epilepsy Res 49:49–60
Acknowledgments
MN and VAY formulated the present hypothesis and they were responsible for writing the report. SG and ACU were responsible for analyses of the data. HRK made critical revision for the manuscript. The study was partially supported by Scientific Research Unit of Suleyman Demirel University (Protocol Number: 1885-TU-09).
Conflict of interest
There is no conflict interest in the current study.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Yürekli, V.A., Gürler, S., Nazıroğlu, M. et al. Zonisamide Attenuates MPP(+)-Induced Oxidative Toxicity Through Modulation of Ca2+ Signaling and Caspase-3 Activity in Neuronal PC12 Cells. Cell Mol Neurobiol 33, 205–212 (2013). https://doi.org/10.1007/s10571-012-9886-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10571-012-9886-3