
Abstract
River ecosystem dynamics are strongly regulated by the surrounding watershed. The availability and sources of energy and nutrient resources that drive these systems are controlled by topography, climate, geology, and position in the watershed. Here we examined particulate and dissolved organic matter (POM and DOM, respectively) and nutrient concentrations across an entire watershed during multiple flow regimes in an unregulated, low gradient river with extensive floodplain forests. The objectives of the study were to: (1) determine the influence of watershed position and floodplain connectivity on POM, DOM, and nutrient concentrations; (2) examine the influence of flow variability on POM, DOM, and nutrient concentrations; and (3) develop an empirical rating curve to predict POM, DOM, and nutrient transport and flux. We sampled POM, DOM, and nutrient concentrations (ammonia, nitrate, nitrite and soluble reactive phosphorus) at ten sites across the Sipsey River watershed that varied in their degree of connectivity to the floodplain over a two-year period. Both watershed position and flow regime influenced POM, DOM, and nutrient concentrations. In particular, a large floodplain swamp in the middle of the watershed, the Sipsey Swamp, strongly controlled the relative availability of particulate and dissolved materials in the water. In the headwaters, there was a greater proportion of particulate material in suspension relative to dissolved carbon. While in the downstream reaches, both within and downstream of the Sipsey Swamp, DOC accompanied by greater molecular mass and more aromatic DOM was in greater quantity than particulate materials at high flows. Nutrient concentrations in the stream water tended to decline through the floodplain swamp across all flow conditions and tended to be lower in high flows. We demonstrate that floodplains can disrupt the upstream–downstream continuum by supplying a large quantity of allochthonous organic matter. Using long-term data we estimated the total annual flux of DOC and nitrate to range between 1221–6500 and 24–35 tonnes per year, respectively, between 2007 and 2017 with the highest flux rates occurring during high flow periods. Our study shows the complex dynamics of a natural floodplain river system and generally supports the flood pulse concept by highlighting the importance of wetland complexes and floodplain connectivity on material and nutrient transport. Description of organic matter and nutrient dynamics in natural low gradient rivers is critical to understanding production of organisms, food webs and ecosystem processes in the face of climate and land use changes.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Resource and nutrient availability for producers and consumers is a primary constraint driving food webs across ecosystems. River ecosystems are complex and autochthonous (in-stream) energy produced and allochthonous (out-of-stream) energy and nutrient sources that enter these systems varies with underlying geology, gradient, climate, and position within the watershed (Hynes 1975; Gregory et al. 1991; Golladay et al. 2000; Minshall 1978). The geomorphic template is well known to govern abiotic habitat conditions in streams, influencing light penetration and allochthonous inputs and consequential composition of biological communities (Vannote et al. 1980; Frissell et al. 1986; Wallace et al. 1997; Atkinson and Cooper 2016). River ecosystems process organic matter inputs from the surrounding watershed and produce new organic matter in dynamic ways varying with hydrologic regime, lateral floodplain connections, as well as environmental factors influencing organic matter decomposition and assimilation (Vannote et al. 1980, Junk et al. 1989; Naiman et al. 1994). The availability and transport of organic matter and nutrients from the terrestrial landscape strongly controls the productivity of stream ecosystems as most stream systems are net heterotrophic, in that microbial processing of terrestrial carbon (C) is a strong driver of whole-system metabolism (Mulholland et al. 2001). The transport and transformation of particulate and dissolved materials within rivers links upstream and downstream communities into an integrated ecosystem.
Several conceptual models have emerged to explain how downstream movement and lateral exchanges between the channel and riparian corridor alters resource availability and quality and the implications for stream organisms. The river continuum concept (RCC) posits that allochthonous detritus dominates energy sources in shaded headwaters with autochthonous primary production becoming more prevalent in wider downstream reaches, which consequently influences aquatic community functional feeding group composition (Vannote et al. 1980). The RCC has been a central tenet in stream ecology for decades. The Flood Pulse Concept (Junk, Bayley, and Sparks 1989) further characterized stream and floodplain systems as being strongly linked with transport of organic material being a function of temporal changes in discharge. Recent conceptualizations (e.g. hierarchical patch dynamics; Poole 2002) include downstream transport and lateral stream connections as being important as well as hyporheic flow in controlling resource supplies to stream biota (i.e., Hyporheic Corridor Concept; Stanford and Ward 1993). The Pulse Shunt Concept (Raymond et al. 2016) further highlights the importance of hydrologic regime, that is, a large fraction of dissolved organic matter (DOM) is processed, recycled, or retained within river reaches during low flows, but then shunted to downstream systems during high flows. Collectively, downstream transport along fluvial continuums, lateral river-floodplain exchanges, and flow regime regulate fluxes and retention of organic material and nutrients, but the relative role of longitudinal versus lateral linkages on nutrient concentrations and organic matter transport in rivers is understudied, particularly across flow regimes and in systems with intact floodplain forests.
The relative quantity and quality of the materials and nutrients exported and transformed are driven by the interactions among availability of material to be transported, stream power, and the efficiency of instream retention (Atkinson et al. 2009; Ensign and Doyle 2006; Gregory et al. 1991; Naiman et al. 1994; Webster et al. 1987). Most organic matter transported in streams is fine particulate organic material (POM; < 1 mm) or dissolved organic matter (DOM) (Webster et al. 1999; Creed et al. 2015). The transport of these materials originates from the erosion and leaching of soil organic matter (Shang et al. 2018; Hedges et al. 1986), processing of leaves and wood (Webster 1983), and sloughing or grazing of periphyton and phytoplankton (Creed et al. 2015; Lamberti et al. 1987; Biggs, Smith and Duncan 1999). The DOM pool is hypothesized to become increasingly uniform with reduced molecular weight and structural complexity during downstream transport, due to increasing contributions of autochthonous compounds and removal of reactive compounds (Vannote et al. 1980; Creed et al. 2015; Raymond et al. 2016). The associated empirical evidence, however, remains scarce, as the majority of relevant studies focused on a limited section of fluvial continuums (e.g., Williams et al. 2010; Lu et al. 2014). Substantial amounts of POM are transported during both base (Webster 1983; Golladay et al. 2000) and high flows (Golladay 1997; Golladay et al. 1987; Webster et al. 1990), while DOC export occurs primarily during large high flow events (Creed et al. 2015; Hu et al. 2016), yet this can vary as a function of stream gradient, watershed position (e.g., the relative distance to headwaters), and antecedent conditions (Raymond et al. 2016; Shang et al. 2018). In addition, nutrient flux and transport also vary due to these similar factors as streams are corridors for nutrient transport while also acting as critical regions where nutrients are removed or transformed (Ensign and Doyle 2006; Peterson et al. 2001). Understanding the drivers and timing of material availability and nutrient concentrations across the stream network is essential for better understanding productivity dynamics, biological communities, longitudinal, and latitudinal linkages at a basin scale.
Frequently flooded low gradient streams with extensive riparian forests are a common feature of the Coastal Plain of the southeastern US (Benke et al. 2000). Fringing riparian forests contribute large quantities of organic material and strongly regulate stream hydrology and biogeochemical cycling (Cuffney 1988; Meyer and Edwards 1990; Benke et al. 2000; Atkinson et al. 2017) making these ecosystems ideal for studying stream-floodplain connectivity. Seasonally pulsed floods create spatial and temporal variation in basal resources (Poff et al. 1997; Atkinson et al. 2009), yet the concentrations of nutrients and how the relative quantities of dissolved versus particulate matter is altered by both fringing floodplains and multiple, complex channels that are characteristic of these systems is still unknown. Previous studies have demonstrated the importance of hydrological regimes in determining the amount, form, and composition of organic materials transported to stream channels (Meyer and Edwards 1990; Golladay et al. 2000; Atkinson et al. 2009; Raymond and Saiers 2010). However, the role of river-floodplain connectivity is often not taken into consideration simultaneously with flow variation and dowstream tranpsort. Shang et al. (2018) showed in a southeastern watershed without extensive floodplain forests that temperature and soil-stream hydrological connectivity both drove the concentrations and the composition of in-stream DOM, but the study was restricted to baseflow conditions. Raymond and Saiers (2010), in a meta-analysis of small eastern streams, demonstrated that 86% of annual DOC export occurred during large hydrologic events, but the analysis was restricted to watersheds with no wetland component. In a Coastal Plain stream with extensive floodplain forests, Golladay et al. (2000) showed that 55-85% of POM transport occurred during seasonal high flows. Yet, the relative strengths of flow regimes, upstream–downstream linkages, and floodplain connectivity as drivers of particulate and dissolved organic material and nutrient fluxes and their spatial patterns in low-gradient stream networks is not fully understood.
Here we examined organic matter and nutrient dynamics across a watershed in an undammed, low-gradient stream across multiple seasons and flow regimes in the southeastern Coastal Plain. In particular, we examined the proportion of particulate and dissolved organic matter and nutrient concentrations across flow regimes at sites having varying lateral connectivity to the floodplain. The objectives of our study were to: (1) determine the influence of watershed position, especially connectivity with an extensive floodplain swamp, on particulate and dissolved organic matter and nutrient concentrations; (2) examine the influence of flow variability on POM, DOC, and nutrient concentrations; and (3) develop an empirical rating curve to predict POM, DOC, and nutrient transport and flux. This study gives insight into the longitudinal versus lateral controls of energy and nutrient availability in a low gradient Coastal Plain stream with intact forested floodplain wetlands and a natural flow regime allowing the ecosystem to still harbor an exceptionally diverse biological community (Haag and Warren 2010). The increasing likelihood of more extreme climatic events (e.g. tropical storms) in the Coastal Plain (Golladay et al. 2016) makes these questions particularly relevant due to the strong impacts on stream flow.
Methods
Study area
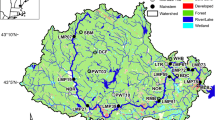
We sampled ten sites along the Sipsey River, Alabama (Fig. 1a) approximately monthly within a single day across a variety of flow regimes and seasons between August 27, 2015 and May 23, 2017 for a total of 18 sampling events (Fig. 1b). The Sipsey is a 2044 km2 alluvial river with extensive forested floodplain wetlands (McGregor and O’Neil 1992). The headwaters begin in the Cumberland Plateau physiographic province and flow through constrained river segments in sandstone lithology. Downstream of site three (see Fig. 1a), the stream flows into the East Gulf Coastal Plain physiographic province before it enters the Tombigbee River (Mobile Basin; Haag and Warren 2010). Within the East Gulf Coastal Plain, the Sipsey flows through mostly Cretaceous gravels of the Fall Line Hills and has an extensive floodplain wetland that includes secondary channels and oxbow lakes. Given this physiography, the river has lower gradient and high floodplain connectivity in the Gulf Coastal Plain section of the watershed (Fig. 2) with the greatest floodplain connectivity (i.e., frequency and extent of overbank flows) occurring in the Sipsey Swamp (floodplain width is 2.5–8 km wide; measured from 1:24,000 US Geological Survey topographic maps). In the southeastern US, streamflow, evapotranspiration and temperature variations are strongly seasonal with periods of low flow occurring during summer and autumn and periods of higher flow and flooding occurring during winter and spring (Benke et al. 2000). Much of the Sipsey River is relatively unmodified by human activities, and major impacts are restricted to the upper and extreme lower sections. Because of the relatively pristine conditions, the Sipsey River supports one of most diverse and abundant biological communities in the region (McGregor and O’Neil 1992), including most of its historical native mussel fauna (Haag and Warren 2010).

a Map of the study area and the sample sites. b Discharge and temperature at Site 8 (USGS gauge 02446500) during the study period. The dashed line represents the annual median flow at the USGS gauge. Arrows at the bottom of the figure indicate the time periods in which sampling was conducted

Physical characteristics of the Sipsey River and its floodplain. Elevation above sea-level is based on measurements obtained from a Garmin GPS. Floodplain width was estimated from U.S. Geological Survey 1:25,000 topographic maps in ArcMap GIS 10.4. Areas between the dotted lines indicate the area that comprises the Sipsey Swamp
Flow characterization
We obtained discharge data from a long term USGS gauge station (74 years; USGS 02446500 located at site 8) to characterize flow patterns in the river. We deployed a Hobo temperature sensor at the gauge location to obtain concurrent temperature data. Additionally, we deployed temperature and pressure transducers at a site upstream (Site 4) and downstream (Site 9) of the USGS gauge. We measured discharge at both Site 4 and 9 when it was possible resulting in ten discharge measurements at each site to develop a rating curve between depth and discharge over our study period for these two sites. At each site we selected a straight and uniform section of the stream and measured depth and velocity (m s−1) every 0.5 meters using a Hach FH950 flow meter (Hach Company, Loveland, CO). Discharge was calculated by multiplying the subsection area by the measured velocity and summing across the section. We also divided our flows into two data categories to help examine patterns across the whole watershed due to flow conditions based on the annual median flow (18.6 m3s−1) at the USGS gauge station (site 8): low flow (flow below the annual median) and high flow (flow above the annual median).
Sampling and lab analyses
Prior to collecting water samples, we used a calibrated YSI multiparameter probe (Yellow Springs, OH) to measure temperature, pH, conductivity, and dissolved oxygen during each sampling trip. We field filtered triplicate samples (100–250 ml) onto pre-ashed 25-mm glass-fiber filters diameter filters (pore size = 0.7 μm; MilliporeSigma, Burlington, MA) for chlorophyll analysis at each site. Filters for chlorophyll were immediately placed on ice and were frozen until being analyzed on a UV–Vis spectrophotometer (Genesys 10S, Thermo Scientific, Waltham, MA) following a 12-h acetone extraction using standard methods (APHA 1995). Water samples were collected in a well-mixed area (i.e. flowing) with acid-washed bottles at each site. We filtered each sample onto pre-ashed and pre-weighed 47-mm pre-weighed glass-fiber filters (pore size = 0.7 μm; MilliporeSigma, Burlington, MA) for gravimetric particulate matter analyses. Filters were dried for 48 h at 50 °C and weighed to the nearest 0.00001 g to determine dry mass (DM). Filters were then ashed in a muffle furnace for 2 h at 500 °C, wetted, and dried for 48 h at 50 °C, reweighed, and ash-free dry mass (AFDM) was determined. Filtrate water samples were frozen and analyzed within 28 days of collection for NH4–N, NO3–N, NO2–N, and soluble reactive phosphorus (PO4–P) analyses on a Lachat Quickchem FIA + 8000 Series flow injection analyzer (Hach Company, Loveland, Colorado, USA). Additional subsamples (230-ml amber bottles per sample) were retained for DOC and DOM analyses. The DOC samples were stored frozen and the DOM samples stored in the dark at 4 °C (in a refrigerator) prior to the analysis. DOM samples were analyzed as soon as possible. While immediate analysis after sample collection yields the most realistic results, it is often not logistically possible. Selecting the optimal storage method for generating accurate DOC and DOM results remains a challenge for biogeochemists. Freezing and acidification are the most commonly used methods to prevent biodegradation. Several studies that have assessed these storage methods reported varied, inconsistent results (Spencer et al. 2007; Peacock et al. 2015; Walker et al. 2016). The direction and magnitude of changes in DOC/DOM during storage are difficult to predict from the initial property and type of the samples (Spencer et al. 2007; Peacock et al. 2015; Walker et al. 2016), but one study showed that the changes may be more significant at high DOC concentrations (Fellman et al. 2008). However, a common pattern found was that freezing/thawing led to large, significant changes in DOM optical properties while usually small changes in DOC concentration. Based on these results, we did not freeze the samples for DOM optical property analysis to avoid potential large shifts in spectroscopic parameters, but we froze the DOC samples to prevent DOC biodegradation loss, given that changes in DOC due to freezing/thawing was usually less than 5% (Peacock et al. 2015; Walker et al. 2016). Detailed description of DOC and DOM measurements can be found in Lu et al. (2015) and Shang et al. (2018). Briefly, DOC concentrations were analyzed using a Shimadzu TOC-V total organic carbon analyzer. Six known concentrations of potassium hydrogen phthalate (KHP) solutions were used to construct calibration curves and carbon-free ultrapure water was regularly measured to assess instrumental baseline. The sample cell was extensively cleaned with ultrapure water and rinsed with sample water at least once before the measuring the sample. The baseline of instrument was evaluated daily through measuring ultrapure water. An external reference standard (Hansell laboratory http://yyy.rsmas.miami.edu/groups/ biogeochem/CRM.html) was analyzed to confirm accuracy. For DOM absorbance, samples were analyzed on a UV-1800 Shimadzu spectrophotometer under the scanning wavelength of from 190 to 670 nm at an interval of 1 nm. We characterized DOM character by specific ultraviolet absorbance at 254 nm (SUVA254; L mg−1 m−1), a photometric measure of DOC aromacity (Weishaar et al. 2003). SUVA254 was calculated as normalized absorption coefficient at 254 nm relative to DOC concentration. Spectral slope ratio (SR) was calculated as the spectral slope gradient (S) of 275–295 nm over 350–400 nm (SR = S275–295 nm/S350–400 nm). SR and SUVA254 provide measures of DOM chemical compositions and quality that are often correlated with the biological source of DOM, that is, terrestrial DOM with higher molecular weight and aromaticity tend to have lower SR and higher SUVA254 values (Weishaar et al. 2003; Helms et al. 2008).
Data analysis
Our goal was to determine how watershed position and stream flow patterns influenced dry mass (DM), ash-free dry mass (AFDM), and chlorophyll concentrations of POM in suspension, as well as the concentration of DOC and nutrients. We used a two-way ANOVA to determine if there were significant differences in dry-mass (DM), ash-free dry mass (AFDM), and chlorophyll-a concentrations of the suspended particulate matter across our two flow categories (high vs. low) and sites. We also used a two-way ANOVA to determine if DOC, SUVA254, SR, any of the water chemistry parameters (NH4–N, NO3–N, NO2–N, PO4), or our molar DOC: nutrient ratios (DOC: NH4–N + NO3–N + NO2–N and DOC: PO4) varied across the sites and the two flow categories.
Further, using the sites in which we had continuous discharge or depth data (Sites 4, 8, and 9), we aimed to understand how discharge altered the availability of organic materials and nutrients in various reaches of the watershed. The U.S. Geological Survey maintains a stream gauge station at Site 8 in the Sipsey River and the data for the other two sites are from rating curves developed from depth data from our Hobo pressure transducers. We were not able to measure discharge during extremely high flows at our sites associated with the pressure transducers (Sites 4 and 9), thus there is error associated with the upper bounds. We fit linear or quadratic regressions to examine the relationships between discharge and POM, DOC, and water chemistry data to examine how these parameters varied across flow within the sites. Data were transformed (natural logarithm) to meet the assumptions of normal distribution. Significant linear or quadratic fit indicated that concentrations could be estimated from discharge on days when sampling did not occur. We used significant relationships between discharge and DOC and NO3 transport from the USGS gauge site (Site 8) to estimate total transport (see below). We were also interested in the availability of particulate organic matter relative to dissolved organic matter within the sites across flow regimes. We calculated the %POM similar to Golladay et al. (1999) as:
This is a metric of the relative availability of AFDM and DOC resources. We used linear or quadratic equations to examine the relationship between %POM and discharge at sites 4, 8, and 9 to determine under what flow regimes particulate or dissolved materials are more prevalent across this watershed. Lastly, we estimated total annual and seasonal flux at Site 8 (USGS gauge) between 2007 and 2017 based on significant relationships between flow and concentrations from that site. NO3 concentrations between our sampling period and McGregor and O’Neil (1992) coincided, so our estimates are a reasonable approximation. All statistical analyses were conducted using the R statistical environment (R Core Team 2016).
Results
Physiochemical parameters
Temperature was lowest at our most upstream site and tended to increase in a downstream direction (Table 1). There was not a distinct pattern in pH or dissolved oxygen with latitudinal position, but conductivity increased between sites 1 and 3, peaking at 3 and then decreased in the downstream samples sites (Table 1).
Particulate and dissolved organic matter
We found that dry mass (DM) of suspended matter varied significantly across sites (F9,519 = 6.59, P < 0.00001), the two flow categories (F1,519 = 5.33, P < 0.05), and with the interaction of site and flow (F9,519 = 3.72, P < 0.001). Tukey’s HSD indicated that flow category led to significant differences in the total amount of DM in suspension and that the headwater sites were different from sites in the mid-reaches of the basin (Fig. 3a; Table 2). AFDM of particulate matter varied due to site (F9,519 = 6.76, P < 0.00001) and due to the interaction between site and flow (F9,519 = 3.23, P < 0.001), but did not vary due between low and high flows (F1,519 = 1.56, P = 0.21). Tukey’s HSD indicated there were differences in the concentration of AFDM in suspension between the headwater sites and sites in the middle and lower section of the basin (Fig. 3b; Table 2). Chlorophyll-a concentrations of particulate matter varied significantly across sites (F9, 361 = 5.73, P < 0.00001) with the most upstream and downstream sites being different from all others (Fig. 3c; Table 2). Chlorophyll-a concentrations of suspended particulate matter did not vary between flow categories (F1,361 = 0.95, P = 0.33), but did as a result of the interaction between site and flow condition (Fig. 3c; F9,361 = 2.65, P < 0.01). DOC varied across sites (F9,503 = 19.11, P < 0.00001) and tended to be greater in the downstream reaches (Fig. 3; Table 2). DOC also varied due to the interaction between flow conditions and site (F9,503 = 4.83, P < 0.00001), but not due to flow alone (Fig. 3d; F1,503 = 0.09, P = 0.76). SUVA254 of DOM varied significantly across sites (F9,493 = 9.06, P < 0.00001), between flow categories (F1,493 = 16.95, P < 0.00001), and as a result of the interaction (F9,493 = 4.60, P < 0.00001) with higher values being typical of lower reaches of the river and high flows (Fig. 3e; Table 2). DOM SR varied significantly across sites (Fig. 3f; F9,493 = 3.97, P < 0.00001) with SR being lower in the lower reaches and varied as a result of the interaction between site and flow (F9,493 = 2.77, P < 0.01), but not between low and high flows (F1,493 = 1.32, P = 0.25).

Measures of particulate matter, particulate matter quality and DOM parameters going from upstream to downstream across the various flow periods. Measurements made during low flow periods represent flow categories below the median flow and high flows represent measurements made above the median flow (mean ± SE). The area between the dotted lines running parallel to the y-axis indicate the portion of the watershed that flows through the Sipsey Swamp
Dissolved nutrients and ratios
Sites in the middle of the watershed just above the Sipsey swamp tended to have the highest dissolved concentrations of nitrogen. At all sites, nutrient concentrations tended to be greater during low flow periods in comparison to high flows (Fig. 4; Table 2). Specifically, NH4–N concentrations in the water varied significantly across sites (Fig. 4a; F9,519 = 8.23, P < 0.00001), flow categories (F1,519 = 44.39, P < 0.00001), and due to the interaction (F9,519 = 3.18, P < 0.001). NO3–N also varied significantly across sites (Fig. 4b; F9,519 = 24.64, P < 0.00001), flow categories (F1,519 = 25.69, P < 0.00001), and as a result of the interaction (F9,519 = 16.35, P < 0.001), while NO2–N varied as a function of both site (Fig. 4c; F9, 519 = 34.41, P < 0.00001) and flow categories (F1, 519 = 41.02, P < 0.00001), but the interaction between site and flow category was not significant (F9,519 = 1.51, P = 0.25). PO4–P varied across sites (Fig. 4d; F9,519 = 3.7, P < 0.001) and flow categories (F1,519 = 11.63, P < 0.001), but the interaction between site and flow was not significant (F9,519 = 1.51, P = 0.38). Molar DOC:N varied as a result of the sampling site (F9,519 = 2.49, P < 0.01) and the interaction between site and flow category (F9,519 = 3.98, P < 0.00001), but not due to flow category alone (F1,519 = 3.24, P = 0.07). Molar DOC:N tended to be higher during low flows in the upper portion of the watershed and highest in the area of the watershed below the Sipsey Swamp during high flow periods (Fig. 4e). Molar DOC:P tended to be greatest below the Sipsey Swamp during high flow periods and varied significantly between sample sites (F9,519 = 2.85, P < 0.01) and as a result of the interaction between site and flow category (F9,519 = 2.05, P = 0.03), but did not differ due to flow category alone (Fig. 4f; F1,519 = 1.38, P = 0.24).

Water chemistry parameters from our most upstream to our most downstream sampling station across all sampling periods and during low and high flows only. Measurements made during low flow periods represent flow categories below the median flow and high flows representing measurements made above the median flow (mean ± SE). The area between the dotted lines running parallel to the y-axis indicates the portion of the watershed that flows through the Sipsey Swamp
Discharge relationships
At the three sites with daily discharge available, the quantity of suspended particulate matter AFDM was only well predicted by discharge at site 4 (y = 0.42x + 0.07, r2 = 0.38, P < 0.01), but not at sites 8 and 9 (Fig. 5). Discharge was a good predictor of DOC concentrations at sites 8 (y = 0.99x + 0.14, r2 = 0.31, P = 0.02) and 9 (y = 0.24x + 0.93, r2 = 0.29, P = 0.02), but not site 4 (Fig. 5). When examining the relative availability of suspended particulate organic matter to dissolved carbon (%POM), discharge was a good predictor across all three sites (Fig. 6). Water chemistry parameters were not well explained by discharge (Supplemental Fig. 1). Overall, only NO3–N concentrations could be predicted by a quadratic function at sites 8 (y = − 0.40x2 + 1.21x + 4.31, r2 = 0.60, P = 0.001) and 9 (y = − 0.67x2 + 1.65x + 4.05, r2 = 0.59, P = 0.001), while other nutrient concentrations were not well predicted by discharge. This suggests other mechanisms are controlling the availability and processing of nutrients in this watershed.

AFDM and DOC 96 km from the headwaters (Site 4; upstream of the Sipsey Swamp), 194 km from the headwaters (Site 8; within the Sipsey Swamp), and 227 km from the headwaters (Site 9; downstream of the Sipsey Swamp) across all the flow periods

The %POM, indicating the relative availability of particulate matter to dissolved organic carbon, at 96 km from the headwaters (Site 4; upstream of the Sipsey Swamp), 194 km from the headwaters (Site 8; within the Sipsey Swamp), and 227 km from the headwaters Site 9; downstream of the Sipsey Swamp) across all the flow periods
Total flux
The long-term gauge data indicated a great deal of variation in average annual discharge (range: 15.8–47.6 cms) across the ten-year period (Fig. 7a). Using the long-term gauge data and the rating curve developed from our collected samples (Fig. 5) at the USGS gauge, we estimated the total annual flux of DOC at Site 8 to range between 1221 and 6500 tonnes per year between 2007 and 2017 with the months of January through June contributing 58–93% of the flux (Fig. 7b). These are the months in which the Sipsey Swamp is typically inundated. The total flux of NO3-N from Site 8 (curve shown in Supplemental Fig. 1) ranged between 24 and 35 tonnes per year with the each half of the year contributing a near equal portion of the flux (Fig. 7b).

a Average annual discharge at the USGS Gauge (USGS 02446500) on the Sipsey River at Elrod, AL. The flow data were used to estimate total DOC (b) and NO3–N flux over a 10 year period (2007–2017) with flux between the typical high flow period (Jan–June) separated from the typical low flow period (July–Dec)
Discussion
Ecologists have long been fascinated by interfaces and the exchange of materials across aquatic-terrestrial boundaries (Polis et al. 1997). We found that the areas within the Sipsey with broad floodplains act as important source areas that supply abundant DOC during high flow periods and tend to trap and process particulate materials. Riparian forests are a substantial source of organic inputs to streams at all times, but our data highlight that high flow periods are important, particularly in the headwaters and mid-reaches. The Sipsey floodplain swamp occurs in the mid-reaches of this watershed and seems to act as a sponge in that it reduces the downstream flux of particulate material during high flows, but exports aromatic, high-molecular-weight dissolved organic matter to downstream reaches during high flow periods. This area of the watershed strongly controls the composition of material fluxes through processing of particulates and release of dissolved materials to more constrained reaches downstream.
In general, our stream met the expectations of the RCC in that the quantity of suspended POM increased in the downstream reaches. However, the significant changes in POM quantity and the source and quality of DOM (more plant-derived compounds with higher aromaticity and structural complexity) corresponding to the presence of Sipsey Swamp demonstrates that floodplains can disrupt the upstream–downstream continuum via supplying a large quantity of allochthonous organic matter to nearby and downstream reaches. This influence is particularly evident at higher flows, as shown by that SUVA254 was greater and SR was lower within and downstream of the swamp during high flow periods indicating continued influx of allochthonous materials. Furthermore, the DOC to nutrient ratios were typically higher than the Redfield Ratio (106:16:1; Redfield 1958), suggesting the significance of allochthonous organic materials to the river. DOC: TN and DOC:SRP ratios were higher within and downstream of the Sipsey Swamp section, particularly during high flows, indicating that the swamp acts as a source for DOM and a sink for nutrients or dilutes nutrients. In particular, there was dilution of all N compounds with high flows within the swamp and downstream, but little change in PO4 with flow suggesting that N is getting diluted during high flows while there is little change in P. Similar to the findings of They et al. (2017) in floodplain lakes, we also found that DOC:N and DOC:P tended to be higher during higher flows, which shows greater river-floodplain connectivity led to larger amounts of swamp sourced organic materials being exported to the main stream channel. Our observations support the Flood Pulse Concept (Junk et al. 1989). Indeed, floodplain inundation is known to support productivity and maintain biological diversity in large rivers and low gradient streams (Flood Pulse Concept; Amoros and Bornette 2002; Junk et al. 1989). Our results indicate that our wetland floodplain zones contribute food resources to the stream and shift the stream segments to being a source or a sink of particulate and dissolved organic carbon based on flow condition, yet upstream reaches are important for bringing in and transporting these materials downstream. This highlights the importance of both longitudinal and lateral connectivity particularly in Coastal Plain river ecosystems. The system studied here is uncommon in North America, as it is unimpounded, and is a relatively undisturbed floodplain stream (e.g., Benke 1990). Thus, our results reveal the natural processing of organic materials and nutrients and the importance of the natural floodplains and flow regime. The natural seasonal flood pulse of this river is likely important for not only the processing of these materials and nutrients, but also community and food web structure (Poff et al. 1997; Freeman et al. 2001; Taylor et al. 2008). Riparian forests, particularly those on reaches with broad floodplains, should be maintained or restored to support instream food webs.
The Sipsey Swamp floodplain appears to function as a sink for nutrients, retaining or taking up nutrients, in particular inorganic N, as nutrient concentrations declined as the stream flowed through the swamp complex. This supports the idea that riverine wetlands serve as areas of high nutrient uptake (Ward 1989). Wetlands have been recognized as hot spots for denitrification due to increased water residence time, low oxygen conditions, and providing additional supplies of organic carbon (Czuba et al. 2018). In an Orinoco River floodplain lake, Hamilton and Lewis (1987) observed that the lake had a net retention of carbon, nitrogen and total phosphorus, and was a net source of soluble reactive phosphorus. The fringing floodplain Sipsey Swamp also seemed to retain or take up particulate organic matter and nutrients. The full mechanisms underlying the reduction in nutrient concentrations as the river flowed through the Sipsey Swamp are not understood but could be due to physical sorption, denitrification and anaerobic ammonium oxidation activity and direct bacterial and primary producer uptake (Burt et al. 1999; Naiman and Decamps 1997; Zedler 2003). Future studies should examine the relative contributions of these mechanisms to nutrient dynamics.
In contrast, Sipsey Swamp floodplains act as a significant source of allochtonous DOM particularly during high river flows. The importance of wetlands as a watershed source of DOM has been recognized in previous studies (Yamashita et al. 2010; Shafiquzzaman et al. 2014). Organic matter accumulates in wetlands due to a combination of high plant productivity and slow decomposition rates; flood inundation mobilizes this pool of organic matter to receiving streams. Wetland sourced DOM is often characterized by high structure complexity and molecular weight ((Shafiquzzaman et al. 2014; Yamashita et al. 2010); also shown in the SUVA254 and SR values in the present study), implying that they are resistant to bacterial decomposition and assimilation. However, these DOM compounds are highly susceptible to photochemical alterations and can be broken down into smaller compounds that are readily available to microbes (Lu et al. 2013). Discharge and light penetration to stream reaches are likely the most important factors determining when and how far these potentially bioavailable compounds are transported downstream and stimulate heterotrophic respiration therein. In the Sipsey River, however, DOM downstream of the swamp largely retained the wetland-related characters (i.e., high SUVA254 and low SR values, Fig. 4), indicating that photochemistry had not significantly altered DOM flushed out floodplains.
Natural high flow pulses are essential for maintaining these systems (Poff et al. 1997; Lambert et al. 2016). The natural flow regime has been shown to be essential in the establishment of diverse ecological communities within this ecosystem (Starr et al. 2014; Tronstad et al. 2005b; Tronstad et al. 2005a; Kennedy et al. 2007). In the Sipsey River, high flow pulses are essential in maintaining production of invertebrate communities (Tronstad et al. 2005b, 2007) and riparian tree growth (Rypel et al. 2009). Base flow conditions are also essential for maintaining the development of biological communities (Poff and Zimmerman 2010; Poff et al. 1997), and in the Sipsey have been shown to be an important condition for maintaining the growth of the diverse freshwater unionid mussel communities found in the downstream segment of this river (Rypel et al. 2009; Haag and Warren 2010). The timing of flows is also an essential component to material transport (Dhillon and Inamdar 2014). We acknowledge that we were not able to sample across the rising and falling limbs of flow periods, thus our results do not capture rapid changes during a high flow event, which may lead to biases in estimating material fluxes during floods. Further work during the rising and falling limbs would more accurately determine the timing of material transport in these dynamic wetland-riverine ecosystems.
Our results do indicate that high flows might be particularly important for supplying food resources to the stream. Here, particulate material was transported in greater concentrations during high flows in the upper and lower portions of the watershed during high flows and the highly connected swamp in the mid-reaches exported higher concentrations of DOC during high flows. Reductions in the surface area of active (natural) floodplains could have significant implications for floodplain–river food webs (Edwards 1987; Zeug and Winemiller 2008). As shown here, these fringing floodplain wetlands act as potentially active area for nutrient cycling and uptake and as an important source area for DOM. Furthermore, inundated floodplains are important habitat for young fish and allow movement of organisms between the floodplains and main channel (Zeug and Winemiller 2008; Junk et al. 1989; Poff et al. 1997). Unconstrained stream reaches appear to be important source areas for material scavenged during floods (Golladay et al. 2000). Our results indicate that the spatial and temporal complexity of stream ecosystems, including flooding regimes and distribution of floodplain forests, interact to determine the input of basal food resources that are available to stream consumers. The production of aquatic insects has been shown to be supported by microbially enriched detritus from floodplains in other Coastal Plain streams (Edwards et al. 1990; Hall et al. 2001) and our results show that both watershed position and flow dictate material transport in this system.
Floodplains are a significant source of high quality POM (i.e., high organic matter and low C : nutrient ratios) in unconstrained streams (Golladay et al. 2000; Atkinson et al. 2009). Our results show that lowland floodplain swamps alter resource availability in dynamic ways through exporting abundant dissolved organic carbon from the swamp to downstream reaches during high flows and trapping particulate matter during these periods. As larger intensity storm events are to increase in frequency in time, there is an expectation that there may be more frequent high pulse events in headwaters leading to greater transport of dissolved organic matter into large rivers and coastal systems (Raymond et al. 2016). As changes in hydrology are anticipated we may see shifts in the controls on energy and nutrient availability in fluvial ecosystems. Most climate change projections for the southeastern US predict increasing water scarcity and that the frequency of both above- and below-median flows will increase (Golladay et al. 2016). Our work provides a framework for understanding material and nutrient fluxes within a complex riverine network and highlights the important role that broad floodplain wetlands play in altering flux. The flooding regime and the distribution of fringing wetland floodplain forests need to be taken into account to determine the retention, processing and release of energy and nutrients in drainage networks. Future projections suggest long‐term alterations in the interactions between streams and their floodplains with unknown consequences on stream biota and food webs.
References
Amoros C, Bornette G (2002) Connectivity and biocomplexity in waterbodies of riverine floodplains. Freshw Biol 47:761–776
APHA (1995) Standard methods for the examination of water and sewage. American Public Health Association, Washington, DC
Atkinson CL, Cooper JT (2016) Benthic algal community composition across a watershed: coupling processes between land and water. Aquat Ecol 50:315–326
Atkinson CL, Golladay SW, Opsahl SP, Covich AP (2009) Stream discharge and floodplain connections affect seston quality and stable isotopic signatures in a coastal plain stream. J N Am Benthol Soc 28:360–370
Atkinson CL, Allen DC, Davis L, Nickerson ZL (2017) Incorporating ecogeomorphic feedbacks to better understand resiliency in streams: a review and directions forward. Geomorphology 305:123–140
Benke AC (1990) A perspective on America’s vanishing streams. J N Am Benthol Soc 9:77–88
Benke AC, Chaubey I, Ward GM, Dunn EL (2000) Flood pulse dynamics of an unregulated river floodplain in the Southeastern U.S. Coastal Plain. Ecology 81:2730–2741
Biggs BJ, Smith RA, Duncan MJ (1999) Velocity and sediment disturbance of periphyton in headwater streams: biomass and metabolism. J N Am Benthol Soc 18:222–241
Burt T, Matchett L, Goulding K, Webster C, Haycock N (1999) Denitrification in riparian buffer zones: the role of floodplain hydrology. Hydrol Process 13:1451–1463
Creed IF, Mcknight DM, Pellerin BA, Green MB, Bergamaschi BA, Aiken GR, Burns DA, Findlay SE, Shanley JB, Striegl RG (2015) The river as a chemostat: fresh perspectives on dissolved organic matter flowing down the river continuum. Can J Fish Aquat Sci 72:1272–1285
Cuffney TF (1988) Input, movement and exchange of organic-matter within a sub-tropical coastal blackwater river floodplain system. Freshw Biol 19:305–320
Czuba JA, Hansen AT, Foufoula-Georgiou E, Finlay JC (2018) Contextualizing wetlands within a river network to assess nitrate removal and inform watershed management. Water Resour Res 54:1312–1337
Dhillon GS, Inamdar S (2014) Storm event patterns of particulate organic carbon (POC) for large storms and differences with dissolved organic carbon (DOC). Biogeochemistry 118:61–81
Edwards RT (1987) Sestonic bacteria as a food source for filtering invertebrates in 2 southeastern blackwater rivers. Limnol Oceanogr 32:221–234
Edwards RT, Meyer JL, Findlay SEG (1990) The relative contribution of benthic and suspended bacteria to system biomass, production, and metabolism in a low-gradient blackwater river. J N Am Benthol Soc 9:216–228
Ensign SH, Doyle MW (2006) Nutrient spiraling in streams and river networks. J Geophys Res 111:G04009
Fellman JB, D’Arnoreb DV, Hood E (2008) An evaluation of freezing as a preservation technique for analyzing dissolved organic C, N and P in surface water samples. Sci Total Environ 392:305–312
Freeman MC, Bowen ZH, Bovee KD, Irwin ER (2001) Flow and habitat effects on juvenile fish abundance in natural and altered flow regimes. Ecol Appl 11:179–190
Frissell CA, Liss WJ, Warren CE, Hurley MD (1986) A hierarchical framework for stream habitat classification-viewing streams in a watershed context. Environ Manag 10:199–214
Golladay SW (1997) Suspended particulate organic matter concentration and export in streams. J N Am Benthol Soc 16:122–131
Golladay S, Webster J, Benfield E (1987) Changes in stream morphology and storm transport of seston following watershed disturbance. Journal of the North American Benthological Society 6:1–11
Golladay SW, Watt K, Entrekin S, Battle J (2000) Hydrologic and geomorphic controls on suspended particulate organic matter concentration and transport in Ichawaynochaway Creek, Georgia, USA. Arch Hydrobiol 149:655–678
Golladay S, Martin K, Vose J, Wear D, Covich A, Hobbs R, Klepzig K, Likens G, Naiman R, Shearer A (2016) Achievable future conditions as a framework for guiding forest conservation and management. For Ecol Manag 360:80–96
Gregory SV, Swanson FJ, Mckee WA, Cummins KW (1991) An ecosystem perspective of riparian zones. Bioscience 41:540–551
Haag WR, Warren ML (2010) Diversity, abundance, and size structure of bivalve assemblages in the Sipsey River, Alabama. Aquat Conserv 20:655–667
Hall RO, Likens GE, Malcom HM (2001) Trophic basis of invertebrate production in 2 streams at the Hubbard Brook Experimental Forest. J N Am Benthol Soc 20:432–447
Hamilton SK, Lewis WM (1987) Causes of seasonality in the chemistry of a lake on the Orinoco River floodplain, Venezuela. Limnol Oceanogr 32:1277–1290
Hedges JI, Clark WA, Quay PD, Richey JE, Devol AH, Santos M (1986) Compositions and fluxes of particulate organic material in the Amazon River. Limnol Oceanogr 31:717–738
Helms JR, Stubbins A, Ritchie JD, Minor EC, Kieber DJ, Mopper K (2008) Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol Oceanogr 53:955–969
Hu Y, Lu YH, Edmonds JW, Liu CK, Wang S, Das O, Liu J, Zheng CM (2016) Hydrological and land use control of watershed exports of dissolved organic matter in a large arid river basin in northwestern China. J Geophys Res Biogeosci 121:466–478
Hynes HBN (1975) The stream and its valley. Verh Int Ver Limnol 19:1–15
Junk WJ, Bayley PB, Sparks RE (1989) The flood pulse concept in river-floodplain systems. Can Spec Publ Fish Aquat Sci 106:110–127
Kennedy TB, Pugh SA, Culp JJ, Benke A (2007) Quantifying and identifying unionid larvae in drift and on fishes of the Sipsey River, Alabama. Alabama Department of Conservation and Natural Resources, Montgomery
Lambert T, Teodoru CR, Nyoni FC, Bouillon S, Darchambeau F, Massicotte P, Borges AV (2016) Along-stream transport and transformation of dissolved organic matter in a large tropical river. Biogeosciences 13:2727–2741
Lamberti GA, Ashkenas LR, Gregory SV, Steinman AD (1987) Effects of three herbivores on periphyton communities in laboratory streams. J N Am Benthol Soc 6:92–104
Lu YH, Bauer JE, Canuel EA, Yamashita Y, Chambers RM, Jaffe R (2013) Photochemical and microbial alteration of dissolved organic matter in temperate headwater streams associated with different land use. J Geophys Res Biogeosci 118:566–580
Lu YH, Bauer JE, Canuel EA, Chambers RM, Yamashita Y, Jaffé R, Barrett A (2014) Effects of land use on sources and ages of inorganic and organic carbon in temperate headwater streams. Biogeochemistry 119:275–292
Lu YH, Edmonds JW, Yamashita Y, Zhou B, Jaegge A, Baxley M (2015) Spatial variation in the origin and reactivity of dissolved organic matter in Oregon-Washington coastal waters. Ocean Dyn 65:17–32
McGregor S, O’Neil P (1992) The biology and water quality monitoring of the Sipsey River and Lubbub and Bear Creeks, Alabama 1990–1991. Geological Survey of Alabama 169
Meyer JL, Edwards RT (1990) Ecosystem metabolism and turnover of organic carbon along a blackwater river continuum. Ecology 71:668–677
Minshall GW (1978) Autotrophy in stream ecosystems. Bioscience 28:767–771
Mulholland P, Fellows CS, Tank J, Grimm N, Webster J, Hamilton S, Martí E, Ashkenas L, Bowden W, Dodds W (2001) Inter-biome comparison of factors controlling stream metabolism. Freshw Biol 46:1503–1517
Naiman RJ, Decamps H (1997) The ecology of interfaces: riparian zones. Annu Rev Ecol Syst 28:621–658
Naiman RJ, Pinay G, Johnston CA, Pastor J (1994) Beaver influences on the long-term biogeochemical characteristics of boreal forest drainage networks. Ecology 75:905–921
Peacock M, Freeman C, Gauci V, Lebron I, Evans CD (2015) Investigations of freezing and cold storage for the analysis of peatland dissolved organic carbon (DOC) and absorbance properties. Environ Sci 17:1290–1301
Peterson BJ, Wollheim WM, Mulholland PJ, Webster JR, Meyer JL, Tank JL, Martí E, Bowden WB, Valett HM, Hershey AE (2001) Control of nitrogen export from watersheds by headwater streams. Science 292:86–90
Poff NL, Zimmerman JK (2010) Ecological responses to altered flow regimes: a literature review to inform the science and management of environmental flows. Freshw Biol 55:194–205
Poff NL, Allan JD, Bain MB, Karr JR, Prestegaard KL, Richter BD, Sparks RE, Stromberg JC (1997) The natural flow regime. Bioscience 47:769–784
Polis GA, Anderson WB, Holt RD (1997) Toward an integration of landscape and food web ecology: the dynamics of spatially subsidized food webs. Annu Rev Ecol Syst 28:289–316
Poole GC (2002) Fluvial landscape ecology: addressing uniqueness within the river discontinuum. Freshw Biol 47:641–660
R Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Raymond PA, Saiers JE (2010) Event controlled DOC export from forested watersheds. Biogeochemistry 100:197–209
Raymond PA, Saiers JE, Sobczak WV (2016) Hydrological and biogeochemical controls on watershed dissolved organic matter transport: pulse-shunt concept. Ecology 97:5–16
Redfield AC (1958) The biological control of chemical factors in the environment. Am Sci 46:205–221
Rypel AL, Haag WR, Findlay RH (2009) Pervasive hydrologic effects on freshwater mussels and riparian tree in southeastern floodplain ecosystems. Wetlands 29:497–504
Shafiquzzaman M, Ahmed AT, Azam MS, Razzak A, Askri B, Hassan HF, Ravikumar BN, Okuda T (2014) Identification and characterization of dissolved organic matter sources in Kushiro river impacted by a wetland. Ecol Eng 70:459–464
Shang P, Lu Y, Du Y, Jaffé R, Findlay RH, Wynn A (2018) Climatic and watershed controls of dissolved organic matter variation in streams across a gradient of agricultural land use. Sci Total Environ 612:1442–1453
Spencer RG, Bolton L, Baker A (2007) Freeze/thaw and pH effects on freshwater dissolved organic matter fluorescence and absorbance properties from a number of UK locations. Water Res 41:2941–2950
Stanford JA, Ward J (1993) An ecosystem perspective of alluvial rivers: connectivity and the hyporheic corridor. J N Am Benthol Soc 12:48–60
Starr SM, Benstead JP, Sponseller RA (2014) Spatial and temporal organization of macroinvertebrate assemblages in a lowland floodplain ecosystem. Landsc Ecol 29:1017–1031
Taylor CM, Millican DS, Roberts ME, Slack WT (2008) Long-term change to fish assemblages and the flow regime in a southeastern US river system after extensive aquatic ecosystem fragmentation. Ecography 31:787–797
They NH, Amado AM, Cotne JB (2017) Redfield ratios in inland waters: Higher biological control of C: N: P ratios in tropical semi-arid high water residence time lakes. Front Microbiol 8:1505
Tronstad LM, Tronstad BP, Benke AC (2005a) Invertebrate responses to decreasing water levels in a subtropical river floodplain wetland. Wetlands 25:583–593
Tronstad LM, Tronstad BP, Benke AC (2005b) Invertebrate seedbanks: rehydration of soil from an unregulated river floodplain in the southeastern US. Freshw Biol 50:646–655
Tronstad LM, Tronstad BP, Benke AC (2007) Aerial colonization and growth: rapid invertebrate responses to temporary aquatic habitats in a river floodplain. J N Am Benthol Soc 26:460–471
Vannote RL, Minshall GW, Cummins KW, Sedell JR, Cushing CE (1980) The river continuum concept. Can J Fish Aquat Sci 37:130–137
Walker BD, Griffin S, Druffel ERM (2016) Effect of acidified versus frozen storage on marine dissolved organic carbon concentration and isotopic composition. Radiocarbon 2016:1–15
Wallace JB, Eggert SL, Meyer JL, Webster JR (1997) Multiple trophic levels of a forest stream linked to terrestrial litter inputs. Science 277:102–104
Ward JV (1989) The 4-dimensional nature of lotic ecosystems. J N Am Benthol Soc 8:2–8
Webster JR (1983) The role of benthic macroinvertebrates in detritus dynamics of streams: a computer simulation. Ecol Monogr 53:383–404
Webster JR, Benfield EF, Golladay SW, Hill BH, Hornick LE, Kazmierczak RF Jr, Perry WB (1987) Experimental studies of physical factors affecting seston transport in streams. Limnol Oceanogr 32:848–863
Webster J, Golladay S, Benfield E, D’angelo D, Peters G (1990) Effects of forest disturbance on particulate organic matter budgets of small streams. J N Am Benthol Soc 9:120–140
Webster J, Benfield E, Ehrman T, Schaeffer M, Tank J, Hutchens J, D’angelo D (1999) What happens to allochthonous material that falls into streams? A synthesis of new and published information from Coweeta. Freshw Biol 41:687–705
Weishaar JL, Aiken GR, Bergamaschi BA, Fram MS, Fujii R, Mopper K (2003) Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ Sci Technol 37:4702–4708
Williams CJ, Yamashita Y, Wilson HF, Jaffe´ R, Xenopoulos MA (2010) Unraveling the role of land use and microbial activity in shaping dissolved organic matter characteristics in stream ecosystems. Limnol Oceanogr 55:1159–1171
Yamashita Y, Scinto LJ, Maie N, Jaffe R (2010) Dissolved organic matter characteristics across a subtropical wetland’s landscape: Application of optical properties in the assessment of environmental dynamics. Ecosystems 13:1006–1019
Zedler JB (2003) Wetlands at your service: reducing impacts of agriculture at the watershed scale. Front Ecol Environ 1:65–72
Zeug SC, Winemiller KO (2008) Evidence supporting the importance of terrestrial carbon in a large-river food web. Ecology 89:1733–1743
Acknowledgements
This work would not have been possible without field assistance from Mark Dedmon, Zach Nickerson, and Monica Winebarger. Anne Belle was responsible for all the water chemistry analyses. The University of Alabama Center for Freshwater Studies, the Weyerhaueser Corporation, and the National Science Foundation DEB-1831512 provided support for this this project. Stephen Golladay provided helpful comments and suggestions on an earlier version of this manuscript. We appreciate the comments of two anonymous reviewers on a previous version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Responsible Editor: Jack Brookshire.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Atkinson, C.L., van Ee, B.C., Lu, Y. et al. Wetland floodplain flux: temporal and spatial availability of organic matter and dissolved nutrients in an unmodified river. Biogeochemistry 142, 395–411 (2019). https://doi.org/10.1007/s10533-019-00542-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-019-00542-z