
Abstract
Combining stable carbon isotopic signatures (δ13C-DOC) and optical properties of dissolved organic matter (DOM), we examined spatial variability in the sources and reactivity of DOM from Oregon-Washington coastal waters, with a particular focus on evaluating whether these measurements may reliably trace terrigenous DOM in coastal oceans. We sampled four stations on the continental shelf and four stations on the continental slope near the mouth of the Columbia River, with sampling depths ranging from 0 to 1,678 m. Nitrate and phosphate concentrations were largely controlled by organic matter (OM) regeneration although the river plume may have led to excess nitrates in relation to phosphates near the river mouth and/or the surface. Four fluorescence components (C1 to C4) were identified by using excitation emission matrices-parallel factor analysis. C1 and C2 were assigned as humic-like components which represented degraded DOM rather than OM of allochthonous or autochthonous origin. C3 and C4 were both labile, protein-like components representing autochthonous contributions, while C4 was more sensitive to diagenesis. In the shallow water layer (salinity ≤32.5 and depth ≤50 m), the variation in absorption properties (SUVA254 and ɛ280), fluorescence index, freshness index (β/α), percent fluorescence of C3, and δ13C-DOC revealed that the Columbia River plume exported DOM that was characterized by greater aromaticity, higher molecular weight, and being more decomposed than marine, autochthonous DOM. However, these signatures of terrigenous DOM disappeared rapidly with increasing depth and offshore distance. In the intermediate and deep water layers (salinity >32.5), the DOM indices were most driven by diagenesis, with changes in percent fluorescence components indicating increasing accumulation of humic DOM relative to protein-like DOM with depth. Principal component analysis that collectively assessed the DOM indices suggests that diagenesis was the primary factor driving the spatial variability of DOM properties in the study region, underlining challenges in tracing allochthonous DOM in coastal waters.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
1 Introduction
Dissolved organic matter (DOM) plays a pivotal role in a variety of environmental and ecosystem processes in aquatic systems. DOM attenuates ultraviolet-B penetration and provides protection to aquatic biota (Williamson and Zagarese 1994), influences the forms and mobility of ecotoxins and pollutants (Driscoll et al. 1988; Worrall et al. 1997; Yamashita and Jaffé 2008), as well as provides basal substrate and energy to heterotrophic food webs (Benner 2003; Kirchman 2003). In coastal waters, where terrigenous, allochthonous DOM (i.e., DOM originating from land and transported to oceans via rivers) and marine, autochthonous DOM meet, large spatial variability in DOM properties has been reported (Guo et al. 2007; Para et al. 2010). In particular, coastal environments represent a hotspot where terrigenous DOM is diluted, transformed, or removed (Opsahl and Benner 1997; Para et al. 2010). However, it remains challenging in tracing terrigenous DOM after it enters the ocean, and thus the reactivity and fate of terrigenous DOM in marine environments remain poorly described.
As DOM is a complex mixture of various constituents, multiple indices are usually necessary to trace terrigenous DOM in marine waters. Lignins, as an abundant component in terrestrial vascular plants, have been long considered as the most effective tracer of terrestrial inputs to marine environments (e.g., Opsahl et al. 1999; Walker et al. 2009). However, lignin phenol analysis is not only expensive, time consuming, and labor intensive but also requires large volumes of water samples that may be difficult to obtain in some research cruises. More recent studies have demonstrated that DOM optical measurement, which is a more rapid and less expensive approach requiring only a small volume of water, may generate a suite of indices for tracing terrigenous DOM (Murphy et al. 2008; Walker et al. 2009, 2013; Fichot et al. 2013). This approach evaluates the light-absorbing fraction of DOM (also referred to as “chromophoric DOM” or “CDOM”) through absorption and fluorescence measurements (Mopper and Schultz 1993; Prahl and Coble 1994). More recently, the development of three-dimensional excitation and emission matrix (EEM)–parallel factor analysis (PARAFAC) technique allows differentiating components with dissimilar chemical structures and compositions contributing to CDOM, such as humic-like and protein-like substances (Stedmon et al. 2003). Such information, in combination with the information from more conventional, one-dimensional absorption characterization, allows inferring CDOM sources in relation to its reactivity to microbial and photochemical alterations, yielding invaluable insights into sources and processes responsible for the production, transformation, and degradation of DOM (Kowalczuk et al. 2003; Yamashita and Tanoue 2003; Guo et al. 2007; Murphy et al. 2008). Notably, a few recent studies have reported correlations between EEM-PARAFAC components or absorption indices and more source-specific lignin phenols (Walker et al. 2009, 2013; Osburn and Stedmon 2011; Fichot et al. 2013), illustrating the great potential for developing and applying optical property-based indices for tracing terrigenous OM in oceans.
Using DOM optical properties, the assessment of mixing between terrigenous and marine DOM has been conducted largely in estuaries and surface, near-shore coastal waters, where CDOM signatures usually showed inverse relationships with salinity to various extents (Kowalczuk et al. 2003; Guo et al. 2007; Para et al. 2010). However, what remains insufficiently studied is vertical and lateral transport of terrigenous DOM to deeper and more offshore coastal waters. In these regions, a steep salinity gradient is not found, but terrigenous DOM can be widely distributed through active physical mixing processes such as river plumes and Ekman transport. In the present study, we sampled waters off the Washington-Oregon coasts of various distances and depths, and assessed the suitability of DOM optical properties in tracing terrigenous DOM distribution. Our study site was near the mouth of the Columbia River, which is the largest source of freshwater from the US West Coast to the Eastern Pacific Ocean. Because the spatial extent and amount of terrigenous DOM in coastal environments are often associated with the magnitude of freshwater inputs (Blough et al. 1993; Rochelle-Newall and Fisher 2002), the study area has the potential to be significantly influenced by terrigenous DOM. Our sampling region also included deeper and more offshore waters, where riverine DOM may have travelled through seasonal Ekman transport and associated upwelling/downwelling. In addition to a suite of DOM indices derived from absorption and fluorescence measurements, stable carbon isotopic composition of DOC (δ13C-DOC) was measured as an additional valuable tool to distinguish between terrigenous and marine materials in estuarine and marine environments (Benner et al. 2005; Osburn and Stedmon 2011). Our results provide insights into whether measuring δ13C-DOC values and DOM optical properties may provide effective and reliable tracers of terrigenous DOM in oceans.
2 Methods
2.1 Study site
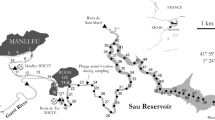
As part of the Early Career Investigator Oceanographic Research Training Cruise (cruise W1106A) organized by the University-National Oceanographic Laboratory System (UNOLS) in July 9–14, 2011, we sampled off the Washington-Oregon coasts, including four stations on the continental shelf and four stations on the continental slope (Fig. 1, Table 1). The major current in the study region is the wind-driven California Current that is derived from the Subarctic Current. The California Current flows southward and is generally strongest in summer, influencing shallow to middle-depth water over the continental shelf and slope (Huyer et al. 1975; Hickey 1979; Huyer 1979; Hill and Wheeler 2002). The dominant alongshore wind stress during our sampling was southward, resulting in offshore Ekman transport that often induces upwelling off the Oregon-Washington coasts in summer. However, during our sampling cruise, the sea surface temperatures and coastal upwelling indices indicated that the sampling stations were not significantly influenced by coastal upwelling, which mostly occurred in the region between 35 and 44°N, south of our sampling stations (Data from NOAA Coast Watch, http://coastwatch.pfel.noaa.gov/elnino.html).

Map showing the eight sampling stations off the Washington-Oregon coasts
All the sampling stations were situated at a region where the Columbia River plume could reach and influence surface water chemistry (http://bragg.coas.oregonstate.edu/ORCoast/Calendars/Calendar_201107; Fiedler and Laurs 1990; Lohan and Bruland 2006; Aguilar-Islas and Bruland 2006). The average discharge of the Columbia River was 4,057 m3/s in 2011, and our sampling captured a period of relatively high discharges, i.e., average 7,098 m3/s for the sampling days. High river discharges are typical from May to June, created by snowmelt, and then usually decrease to minimum flow during August to September (Neal 1972; Thomas and Weatherbee 2006).
2.2 Sample collection and analysis
Sample collection
All containers and sampling equipment that could be in direct contact with sampled water were either combusted at 450 °C for 5 h for glass materials or acid soaked (10 % HCl) and thoroughly rinsed with Milli-Q for plastic materials. Water samples were collected aboard the R/V Wecoma using Niskin bottles on a 12-bottle rosette. Samples were then transferred to polycarbonate bottles and approximately 500 mL water was immediately filtered through pre-combusted GF/F filters (Whatman, nominal size 0.7 μm). The filtrates were stored frozen in the dark prior to subsequent analyses.
DOC concentration and stable carbon isotopes
The water samples were submitted to G. G. Hatch Isotopic Laboratory at the University of Ottawa for the analysis of DOC concentrations and δ13C-DOC. Samples were run on an OI Analytical “TIC-TOC” Combustion Analyzer (Model 1030) interfaced with continuous flow to a Finnigan Mat DeltaPlusXP isotope ratio mass spectrometer. Blank waters were run between every sample. Data were normalized using internal standards (sucrose and potassium phthalate) that were calibrated against the usual international standards (USGS40 and 41). The 2 sigma analytical precision was 2 % for the DOC concentration, and +/− 0.2 ‰ for the isotopes.
Absorption properties
Absorbance of CDOM was measured using a UV–visible Spectrophotometer (Shimadzu UV-1800) from 190 to 700 nm at an interval of 1 nm. All samples were measured in a 1-cm quartz cell. Between each sample, the sample cell was extensively cleaned with Milli-Q water. The baseline of the instrument was checked regularly through Milli-Q water measurements.
Fluorescence properties
Fluorescence measurement was conducted by using a Fluoromax 4 (Horiba), following the procedure detailed in Yamashita et al. (2011). Excitation wavelengths spanned from 240 to 450 nm using 5-nm increments, and the corresponding emission spectrum was collected from 320 to 550 nm at 2-nm increments. Fluorescence spectra were corrected for inner filter corrections using UV–visible absorption spectra according to McKnight et al. (2001). The EEM of Milli-Q water was subtracted from sample EEMs. The excitation and emission correction factors supplied by the manufacturer were applied for correction of instrument-specific responses (Cory et al. 2010). Fluorescence intensity was daily corrected to the area under the water Raman peak (excitation = 350 nm). Following the statistical approach used in Stedmon et al. (2003), the PARAFAC model was constructed in MATLAB using the DOMFluor toolbox from Stedmon and Bro (2008). A four-component EEM-PARAFAC model (C1–C4) was validated by split-half analysis and random initialization. The relative abundance of each of these four fluorescent components (Ci, i = 1 to 4) was calculated as:
where F Ci represented fluorescence intensity of each specific fluorescent component and TF was total fluorescence intensity.
Ancillary parameters
Samples were submitted to Dauphin Island Sea Laboratory (AL, USA) for the analysis of dissolved nutrients (phosphate, nitrate, nitrite, and ammonium). The samples were measured on a Skalar San + continuous flow autoanalyser with wet chemistry modules designed for standard colorimetric techniques for the individual analytes (NO2 − and NO3 − consistent with EPA method 353.2, NH3 consistent with EPA method 350.1, and PO4 3− consistent with EPA method 365.3). Sample absorbance was compared with regression statistics based on a five-point standard curve for each analyte, and the results were baseline and drift corrected throughout the sample run.
In situ physical and biological data, including depth, salinity, temperature, dissolved oxygen, photosynthetically active radiation (PAR), and chlorophyll-a (Chl-a) fluorescence, were obtained from a SeaBird Electronics plus 19 conductivity temperature depth (CTD) profiler equipped with a dissolved oxygen sensor and a Chl-a fluorometer aboard the R/V Wecoma.
Statistical analysis
The significance level, α, was set at 0.05 for all statistical analyses. Non-parametric, Spearman correlation was conducted to assess the correlation of two variables. A subset of DOM parameters was selected for principal components analysis (PCA) based on the assumptions from PCA: (1) sampling adequacy, which was measured with the Kaiser-Meyer-Olkin test (>0.5); (2) intercorrelation between selected variables (Bartlett’s test of sphericity, p < 0.5) without extreme multicollinearity (the determinant of the correlation matrix >0.00001). DOM parameters were standardized prior to the PCA analysis, and PCA components with eigenvalues >1 were extracted.
3 Results and discussion
3.1 Physical and chemical context
Based on the salinity-depth profiles of our sampling stations, the water column was divided into three layers (Fig. 2)—a shallow layer where salinity increased rapidly with depth, an intermediate layer where salinity increased gradually with depth, and a deep layer where salinity remained relatively stable. The salinity value delineating the three layers was the same for all the sampling stations, that is, 32.5 between the shallow and intermediate layer and 33.5 between the intermediate and deep layer (Table 1). The salinity delineating shallow versus intermediate layer was consistent with the traditional delineation boundary for the Columbia River plum water, which was historically defined as waters contiguous to the mouth of Columbia River with salinities less than 32.5 (Barnes et al. 1972). Formed as a result of mixing of river water and coastal seawater, the plume is often manifested as shallow (2–20 m) surface lens of low-salinity water in coastal waters (Hickey et al. 1998) and acts as a conduit for delivering macro- and micro-nutrients from land to coastal oceans (Hill and Wheeler 2002; Aguilar-Islas and Bruland 2006; Lohan and Bruland 2006). In our sampling month, this plume was transported offshore and southward by Ekman transport through the extent of the study region (http://bragg.coas.oregonstate.edu/ORCoast/Calendars/Calendar_201107). Our samples in the shallow layer showed salinities ranging between 23.8 and 32.5, indicating that they were influenced by the plume.

Vertical profiles of salinity for a the four sampling stations on the continental shelf, and b the four sampling stations on the continental slope off the Washington-Oregon coasts
The concentrations of nitrate (NO3 −) and phosphate (PO4 3−) showed an overall increasing trend with depth, and they were positively correlated with each other for the majority of the samples (Fig. 3). Excluding those samples with disproportionally high nitrate concentrations, the slope of the nitrate vs. phosphate regression line was ca. 15 (Fig. 3), which is similar to the Redfield ratio (16:1) and the 14:1–15:1 ratio found in the global ocean waters (Tyrrell and Law 1997). This pattern suggests that organic matter regeneration regulated the variation in the concentrations of nitrate and phosphate in most of our samples. On the other hand, the samples with disproportionately high nitrate values were found in the shallow water layer with salinity <32.5, as well as in the deep layer from the station closest to the Columbia River mouth, i.e., shelf-3, indicating that freshwater input associated with the Columbia River plume may be responsible for the excess nitrate in relation to phosphate. Previous work has shown that the Columbia River plume water was characterized by high ratios of nitrate to phosphate, i.e., 40 μM nitrate and 1 μM phosphate in winter and spring while 10 μM nitrate and 0.25 μM phosphate in summer and autumn (Sullivan et al. 2001).

Nitrate versus phosphate concentrations in the water samples from the eight sampling stations off the Washington-Oregon coasts
3.2 Concentrations and stable carbon isotopic values of DOC
In the shallow layer, the DOC concentrations ranged between 58 and 153 μM and averaged 98 ± 25 μM, comparable to the range previously reported for the surface waters off the Oregon and Washington coasts, i.e., 74 to 105 μM (Hill and Wheeler 2002). The DOC concentrations did not vary as a function of salinity in the shallow layer (Table 2), suggesting similar DOC values between riverine and coastal surface waters in the study area. The DOC values for the intermediate and deep water layers averaged 88 ± 42 μM, and they were overall lower than those in the shallow layer, agreeing with many observations that DOC concentrations decreased with depth in water column due to removal of labile compounds in shallow waters by heterotrophic microbes (Guo et al. 1995; Ogawa and Tanoue 2003).
The δ13C-DOC values ranged between −26.5 and −19.9 ‰ in the study region. Based on a general range for autochthonous and allochthonous materials, i.e., −30 to −18 ‰ for marine, phytoplankton-derived organic carbon and −31 to −27 ‰ for riverine organic carbon (note that riverine signatures incorporate those of terrestrial plants and freshwater phytoplankton ( Montgomery et al. 2000; Retamal et al. 2007; Staal et al. 2007; Lu et al. 2010), the values of our samples suggest the dominance of autochthonous DOC in the study region.
3.3 DOM absorption and fluorescence indices
The absorption properties of DOM provide information about the aromaticity and molecular weight of DOM. Two parameters commonly used are SUVA254 (L mg C−1 m−1), which is defined as the UV absorbance at 254 nm measured in inverse meters normalized to DOC concentration (mg/L) in milligrams per liter (Weishaar et al. 2003), and ɛ 280 (mole C−1 cm−1), commonly defined as molar absorptivity at 280 nm measured in inverse centimeters (Chin et al. 1994). SUVA254 and ɛ 280 have been found to be positively correlated with percent aromatic carbon (Weishaar et al. 2003) and molecular weight (Chin et al. 1994). In general, terrigenous DOM from decomposed higher plants and soils has greater aromaticity and molecular weight than autochthonous DOM (Chin et al. 1994; Sulzberger and Durisch-Kaiser 2009; Lu et al. 2013, 2014), and thus higher SUVA254 and ɛ 280 values may be interpreted as larger contributions of terrigenous DOM.
SUVA254 and ɛ 280 values were strongly correlated with each other in the shallow layer as well as in the intermediate and deep layers (Table 2). SUVA254 averaged 0.8 ± 0.6 L mg C−1 m−1 (n = 47), which was comparable to the reported value for the Pacific Ocean fulvic acid sample (SUVA254 = 0.6 L mg C−1 m−1) (Weishaar et al. 2003). For the shallow layer samples, SUVA254 averaged 1.1 ± 0.8 L mg C−1 m−1, lower than the range observed for freshwater and brackish water samples, e.g., 1.7–5.3 L mg C−1 m−1 for freshwater humic substances from various locations in the US and Antarctica (Weishaar et al. 2003) and 2.6–10.6 L mg C−1 m−1 for samples with salinity between 8 and 26 from the Elizabeth River estuary in the lower Chesapeake Bay (Helms et al. 2008). ɛ 280 values ranged between below detection to 280 mol C−1 cm−1, which were overall lower than the values observed for surface and pore waters from rivers and lakes, e.g., 60–919 mol C−1 cm−1 in Chin et al. (1994). Overall, SUVA254 and ɛ 280 showed that DOM in our samples had lower aromaticity and molecular weight than DOM from terrestrial, freshwater systems, indicating primarily an autochthonous origin.
The fluorescence index (FI) is defined as the ratio of emission intensity (470/520 nm) at 370 nm excitation and has been adjusted since FI was initially introduced as the ratio of fluorescence intensity at 450 and 500 nm (McKnight et al. 2001; Jaffé et al. 2008). FI has been widely used to infer relative contributions of microbial versus terrigenous DOM, on the basis that organic materials from microbial sources generally have greater FI values than those from terrestrial sources (McKnight et al. 2001; Jaffé et al. 2008; Lu et al. 2013, 2014). Although FI and the two absorption indices were all associated with terrigenous versus autochthonous source contributions of DOM, the correlations between FI and SUVA254 and between FI and ɛ 280 were weak and not significant (Table 2), indicating decoupling between these source indicators. In the shallow layer, FI showed a positive correlation with salinity, in agreement with the expectation that the contributions of terrigenous DOM decreased in more saline waters. This correlation, however, was not observed in the intermediate and deep water layers, suggesting a rapid disappearance of source-tracking signals with increasing depth (see more discussion in Section 3.6).
The “freshness index” (β/α), calculated as emission intensity at 380 nm divided by the emission intensity maximum observed between 420 and 435 nm at excitation of 310 nm, has been used to measure the diagenetic status of DOM (Wilson and Xenopoulos 2009; Fellman et al. 2010). Large values of this ratio correspond to less diagenetically altered DOM, as β represents recently microbially produced DOM and α denotes highly decomposed DOM (Wilson and Xenopoulos 2009; Fellman et al. 2010). In our samples, β/α values varied between 0.65 and 1.30, which were overall higher than those values observed in terrestrial systems (e.g., between ca. 0.5 and 0.7 for stream water DOM in Wilson and Xenopoulos 2009). The reliability of β/α as a freshness index is further supported by its positive correlation with the concentrations of dissolved oxygen (DO) in the intermediate and deep layers (Table 2), which is a rough indicator of the relative ages of water masses—older, deeper water not recently in direct contact with shallow, oxygen-rich layers or the atmosphere tends to have low levels of DO. On the other hand, no correlation was found between β/α and the source-tracking parameters, i.e., SUVA254, ɛ 280, and FI (Table 2), suggesting that the DOM diagenetic status was not tightly coupled to its sources in the study region.
3.4 EEM-PARAFAC fluorescence components and their source assignment
Using EEM-PARAFAC, we identified four fluorescence components (Table 3, Fig. 4). Fluorescence components 1 and 2 (C1 and C2) have excitation and emission ranges similar to the “M” and “C” peak, respectively. The “M” and “C” peaks have been found in a diverse range of environments including streams, rivers, and oceans, and they are thought to represent humic-like DOM that is aromatic, highly conjugated, and of high molecular weight (Stedmon and Markager 2005a, b; Coble 2007). Although in some studies the two peaks have been attributed to different sources, i.e., the “M” peak has been related to autochthonous origin while the “C” peak has been associated with terrigenous sources representing riverine OM in marine environments, others have suggested that these two components are ubiquitous products of biological degradation of OM rather than indicating a terrigenous or marine origin (Coble 1996, 2007; Murphy et al. 2008). In our samples, the relative abundances of fluorescence of C1 (%F C1) and C2 (%F C2) were strongly correlated (Table 2), suggesting that these two components were regulated by the same or similar sources/processes. Percent F C2 exhibited a negative correlation with the β/α values as well as with the DO values in the intermediate and deep layers, and percent F C1 negatively correlated with β/α in the shallow layer (note that there was also a weak correlation between %F C2 and β/α, Spearman ρ = −0.48, p = 0.06; Table 2), suggesting that these two fluorescence components represented refractory, more decomposed DOM resulting from OM degradation. C1 and C2 dominated the fluorescent DOM, accounting for 36.1 ± 9.7 and 25.8 ± 7.9 % of total fluorescence, respectively.

Excitation-emission spectra of the four EEM-PARAFAC fluorescence components (C1 –C4)
Fluorescence components 3 and 4 (C3 and C4) showed fluorescence spectra similar to protein-like DOM (Table 3, Fig. 4). Free dissolved tryptophan has an excitation maximum at 278 nm and an emission maximum at 354 nm while free dissolved tyrosine has excitation and emission maximums at 275 and 303 nm, respectively (Stedmon and Markager 2005b). C3 and C4 were assigned as tyrosine-like DOM and tryptophan-like DOM, respectively. In general, protein-like DOM is more enriched in aquatic algae and bacteria than in terrestrial plants and soils (Determann et al. 1998; Stedmon and Markager 2005b), and it is overall more labile to biological utilization than humic and fulvic DOM from higher plants (Fellman et al. 2010; Lu et al. 2013). In our samples, we found that both %F C3 and %F C4 positively correlated with the DO level in the intermediate and deep layers (Table 2), indicating that these components were less abundant in “old” waters and thus supporting that they represented labile components. C3 and C4 comprised 24.2 ± 8.5 and 13.8 ± 14.4 % of total fluorescence, respectively, and they were overall less abundant than degraded, humic-like components represented by C1 and C2.
Despite that C3 and C4 were both assigned as protein-like compounds representing labile, autochthonous sources, their abundances correlated only in the intermediate and deep layers. In the shallow layer, C4 appeared to be driven more by DOM freshness, as evidenced by its correlation to the β/α ratio, while C3 may be regulated more by source inputs as it correlated with salinity and the other source parameters including FI, SUVA254, and ɛ 280 (Table 2). The decoupling of C3 and C4 agrees with the previous observation that these two components may have dissimilar reactivity. For example, Stedmon and Markager (2005b) found that a component similar to C3 was not utilized during bacterial mineralization over a 7-day incubation, while a component similar to C4 can be efficiently removed by both UV irradiation and microbial degradation. These results are similar to our observation that C3 was more a source tracer for autochthonous materials, while C4 was more controlled by DOM diagenetic status.
3.5 Spatial variation in DOM properties
3.5.1 Shallow layer
Salinity is commonly used as a conservative tracer for riverine and marine water mixing in studies of estuarine and coastal systems. In the shallow water layer, neither SUVA254 nor ɛ 280 showed a significant correlation with salinity (Table 2). However, we observed that the two samples with low salinity (∼24) had much larger values for SUVA254 and ɛ 280 than those samples with higher salinity (>28) (Fig. 5a, b). Thus, the weak significance of the correlations may be due to the lack of samples with salinity between 24 and 28, and possibly also the relative small sample size in the shallow layer (n = 17). Similarly, FI and %F C3 exhibited low values for the two fresher samples and clustered values for more saline samples, although both proxies significantly correlated with salinity (Table 2; Fig. 5c, d). The freshness indices showed a similar pattern—β/α and %F C1 had distinct values for the two samples with low salinity (Fig. 5e, f), and both indices varied as a function of salinity (Table 2). These observations demonstrate that the Columbia River plume exported terrigenous DOM of higher aromaticity and molecular weight than in situ DOM to the shallow layer. This terrigenous DOM pool was also more decomposed than autochthonous DOM in the shallow layer, agreeing with the previous observation that terrigenous DOM arriving at coastal waters has usually been altered through the river-to-coastal ocean fluvial transit (Opsahl and Benner 1997; Hernes and Benner 2003; Sulzberger and Durisch-Kaiser 2009). However, the clustered DOM indices at higher salinities demonstrate that riverine DOM signals quickly became indistinguishable with distance from the sources, which may be a combined result of increasing contributions of autochthonous CDOM and further photochemical and microbial alteration of terrigenous CDOM in more offshore environments.

Variation of DOM indices in the shallow layer, including: a SUVA254 vs. salinity; b ɛ280 vs. salinity; c fluorescence index vs. salinity; d %F C3 vs. salinity; e β/α vs. salinity; f %F C1 vs. salinity; g %F C3 vs. δ13C-DOC; and h %FC3 vs. salinity
Another notable pattern is that δ13C-DOC values varied as a function of %F C3 (Table 2, Fig. 5g). Considering %F C3 as an indicator of marine DOM, this correlation indicates that autochthonous DOM was characterized by overall more positive δ13C-DOC values than terrigenous DOM in the study region, which is consistent with previous observations from different geographic regions (Benner et al. 2005; Osburn and Stedmon 2011). However, the lack of a significant correlation between δ13C-DOC and salinity (Table 2), as well as the finding that δ13C-DOC values of the two fresher samples were comparable to those of more saline samples (Fig. 5h), suggests that δ13C-DOC was a less unambiguous tracer of terrigenous DOM than the other source indices in the study region. This is unsurprising as the range for riverine OC (−31 to −27 ‰) and marine OC (−30 to −18 ‰) overlaps (Retamal et al. 2007; Staal et al. 2007). These ranges often vary geographically, and at some regions, δ13C-DOC can be a powerful tracer of terrigenous DOM. For example, Osburn and Stedmon (2011) reported δ13C-DOC values in the Kattega Sea increased with salinity, with an upwardly curving pattern that may represent mixing of terrigenous and marine sources. Overall, only a very limited numbers of studies tracing terrigenous DOM in offshore marine environments have reported δ13C-DOC values, and thus the robustness of δ13C-DOC as a source tracer should be further evaluated.
3.5.2 Intermediate and deep layers
SUVA254, ε 280, and FI showed unsystematic patterns relative to salinity in the intermediate and deep layers (Fig. 6a–d; Table 2), suggesting that these indices cannot trace terrigenous materials in deeper coastal water where terrigenous DOM accounts for only a minor fraction of total OM. In addition, terrigenous compounds may have been significantly altered through the transit from shallow to deep waters. Photochemistry can be an important mechanism in altering and remineralizing terrigenous compounds in euphotic zones (e.g., Sulzberger and Durisch-Kaiser 2009; Para et al. 2010), and terrigenous OM that has survived microbial degradation in rivers and estuaries can become bioavailable to marine microbial communities (e.g., Spencer et al. 2008). It should also be noted that using a longer path length (e.g., 5, 10, or 100 cm) can improve the accuracy/sensitivity of measurements of absorbance indices and thus their suitability as source tracers. For example, using a 5-cm cuvette, Walker et al. (2013) noted that CDOM absorption at 320 nm was better suited to trace riverine OM exported to the Arctic Ocean than EEM-PARAFAC components.

Variation of DOM indices versus salinity or depth in the intermediate and deep layers, including: a SUVA254 vs. salinity; b ɛ280 vs. salinity; c fluorescence index vs. salinity; d β/α vs. salinity; e %F C1 vs. depth; f %F C2 vs. depth; g %F C3 vs. depth; and h %F C4 vs. depth
Positive correlations were found between depth and DOM freshness indices. With increasing depth, %F C1 and %F C2 increased while β/α, %F C3, and %F C4 decreased (Table 2, Fig. 6e–h), indicating that deeper DOM was more diagenetically altered. The values of %F C3 and %F C4 were highly variable above ca. 600 m and remained relative stable and low at depths below this level, while %F C1 and %F C2 remained high and stable below ca. 600 m. These patterns suggest that fresh, labile DOM was rapidly removed in the upper 600 m, resulting in accumulation of humic compounds resistant to microbial degradation in deeper waters. These findings coincide with many coastal and open ocean studies demonstrating that labile and semi-labile DOM is usually removed in the upper approximately 500 m of water columns (Guo et al. 1995; Ogawa and Tanoue 2003). A recent review by Benner and Amon (2014) further highlighted this general trend widely observed in the ocean, noting a shift from high abundance of bioreactive components (i.e., proteins, lipids, carbohydrates) in surface waters to rather uncharacterized, biodegraded compound mixtures in deep waters.
Several patterns that were not found in the shallow layer appeared in the intermediate and deep layers: (1) correlations between DOC concentrations and relative abundance of the fluorescence components—DOC increased with %F C3 and %F C4 but decreased with %F C1 and %F C2 (Table 2), indicating that DOC quantity was regulated by the relative abundance of reactive compounds in deep waters; and (2) significant correlations between the absorption indices (SUVA254 and ɛ 280) and humic fluorescence (%F C1 or %F C2), suggesting that factors controlling the absorption and fluorescence indices were more similar in deep waters. These changes from shallow to deep waters reflect a transition in main factors governing DOM variability with increasing depth. DOM properties were mainly driven by allochthonous versus autochthonous contributions in shallow waters, but more by diagenesis-controlled removal of labile, autochthonous compounds in deep waters. As a result, resistant humic compounds, irrespective of autochthonous or allochthonous origins, became the primary component regulating DOM concentrations and optical properties in the intermediate and deep layers.
3.5.3 PCA analysis integrating shallow, intermediate, and deep water layers
PCA analysis was conducted to integrate the multiple DOM indices generated in order to assessing the variation with depth and salinity. The two samples with the lowest salinity (<25) were not included, as their distinct values would have masked more subtle variation in the other samples. Using sampling adequacy and multicollinearity as selection criteria, we generated a PCA analysis that included β/α, DOC concentrations, %F C2, %F C3, %F C4, and SUVA254.
Two principal components, PC1 and PC2, explained 47 and 28 % of total variance, respectively. PC1 was positively controlled by %F C4 and β/α but negatively by %F C2 (Fig. 7a). Thus, PC1 measured the diagenetic status of DOM, with higher PC1 scores representing fresher DOM and lower scores corresponding to more decomposed DOM. PC1 negatively correlated with depth (Spearman ρ = −0.54, p < 0.001), and PC1 scores were higher and more variable for samples in the upper ca. 100 m (Fig. 7b), suggesting that biological degradation was most active above this depth. PC2 had positive loadings for %F C3 and DOC concentrations and negative loadings for SUVA254. Since source composition mainly influenced SUVA254 and %F C3 in the shallow layer and diagenesis affected the relationships among %F C3, SUVA254, and DOC concentrations (as discussed in the Sections of 3.5.1 and 3.5.2), PC2 was influenced by both the sources and diagentic status of DOM. PC2 did not show a pattern relative to salinity (ρ = −0.24, p = 0.139) or depth (ρ = −0.21, p = 0.178). Samples in the upper ca. 600 m showed more variable scores than those below (Fig. 7c), suggesting that source- and diagenesis-related signatures were largely removed in the upper 600 m, leaving behind deeper DOM with more homogenous properties.

Loadings and scores of principal component analysis of DOM indices: a loadings for principal components 1 and 2 (PC1 and PC2); b PC1 score vs. depth; and c PC2 score vs. depth
3.6 Can CDOM indices effectively trace terrigenous DOM in marine environments?
Although multiple CDOM indices and δ13C-DOC were evaluated in the present study, we did not identify an unambiguous tracer for terrigenous DOM inputs to the intermediate and deep water layers. This observation is in agreement with a number of studies where linear correlations were found between CDOM indices along a steep salinity gradient but became relatively stable in regions with subtle gradients (e.g., Murphy et al. 2008; Walker et al. 2013). As such, it is challenging to reliably trace terrigenous DOM in deeper and more offshore environments solely with CDOM indices.
Results from the PCA analysis revealed part of the reason for the ambiguity of CDOM indices as source tracers—these indices were driven primarily by diagenesis and only secondarily by source contributions. It has been widely reported, from field observations to laboratory incubations, that diagenesis can alter DOM to such an extent that original source signatures are largely removed. Because DOM components reactive to photochemical and microbial degradation appear to share similar isotopic and fluorescence signatures across study regions and systems, the chemical characteristics of residual DOM compounds we observed have been homogenized to various extents (Opsahl and Zepp 2001; Fellman et al. 2009; Lu et al. 2013). For example, 13C-depleted DOM compounds and humic fluorescence components are preferentially degraded photochemically, while protein fluorescence components are preferred by microbial remineralization (Opsahl and Zepp 2001; Fellman et al. 2009; Lu et al. 2013). Aligning with these previous findings, we observed that degraded terrigenous DOM from the two fresher samples in the shallow layer had some indices (e.g., %F C1, β/α) with values more similar to degraded, deep marine DOM than to less-degraded DOM from the shallow and intermediate layers.
It should be pointed out that only a limited number of studies have been conducted to evaluate the suitability of DOM optical properties in tracing terrestrial materials in marine environments (Hernes and Benner 2003; Osburn and Stedmon 2011; Walker et al. 2009). In a study of CDOM in the Canadian Archipelago and coastal Beaufort Sea surface water, Walker et al. (2009) found a positive linear relationship between four EEM-PARAFAC components and dissolved lignin phenols within the riverine plume region, concluding that these components were of a terrestrial origin. Likewise, Osburn and Stedmon (2011) found that the intensity of a fluorescence component correlated significantly with the amount of lignin phenols in the Baltic-North Sea transition zone. Using this component, these researchers were able to estimate the fluxes of allochthonous DOM to seawaters. In most cases, however, the identification of terrestrial fluorescence components did not rely on lignin data. Through characterizing EEM fluorescence in the surface water (4–7 m) of the Atlantic Ocean, Murphy et al. (2008) identified two humic-like fluorescence components representing terrestrial inputs, based on a general distribution that these components had higher intensities in more near-shore waters. These researchers detected these components in the open ocean (up to >200 nautical miles offshore), where the concentrations were estimated to be approximately 1.5 % of the concentrations in rivers. Oftentimes, these source-tracking components are site specific and not transferable to other geographic regions. For instance, the component C2 found in our samples exhibited spectra comparable to one terrestrial component (BERC3; Ex = 370 (265), Em = 488) in Walker et al. (2009), and our component C1 showed similar spectra to the terrestrial component identified by Osburn and Stedmon (2011) (Ex = 240 nm and Em = 424 nm). By comparison, autochthonous production of peak fluorescence that is similar to C2 and BERC 3 has been pointed out by several studies (e.g., Romera-Castillo et al. 2010; Jorgensen et al. 2014; Tanaka et al. 2014). These results strongly indicate that the source of a particular fluorescence component can vary across geographic regions, further demonstrating that CDOM indices may not be reliable as terrestrial source tracer, particularly in environments where autochthonous CDOM is dominating.
Lastly, DOM indices may respond to a suite of interrelated factors and processes, whereby the interpretations of the indices may change even within a specific study region. For example, we observed that SUVA254 and ε 280 were more indicative of source variation in the shallow layer but more of diagenesis in deeper waters. Given that CDOM indices and δ13C-DOC values represent bulk DOM properties susceptible to biases from averaging various components, it is unsurprising that these indices do not always correspond to a specific source or process. Using SUVA254 as an example, the high ratios may be largely due to lignins from rivers but become more related to uncharacterized compound mixtures with increasing depth. Therefore, in order to obtain robust interpretations of sources and degradation processes of DOM, it is necessary that multiple tracers are simultaneously characterized and more sophisticated analytic techniques are incorporated, such as ultra-high-resolution mass spectrometry and source-specific molecular biomarker characterizations.
4 Conclusions
We characterized DOM characteristics from the continental shelf and slope along the Oregon-Washington coasts, and evaluated the usefulness of CDOM indices and δ13C-DOC as tracers of riverine, terrigenous DOM. Results revealed high spatial variability in the sources and reactivity of DOM from the study site. In the shallow layer where riverine inputs were significant, several DOM indices, including SUVA254, ε 280, FI, β/α, δ13C-DOC, and %F C3, showed that the Columbia River plume exported terrigenous DOM of characteristics distinct from marine, autochthonous DOM. However, the terrigenous signatures disappeared rapidly with increasing depth and offshore distance, and the DOM indices did not vary in a predictable fashion in the intermediate and deep layers. Instead, diagenesis became the primary factor controlling DOM properties, as evidenced by the changes in the relative abundance of the fluorescence components relative to depth. PCA analysis further confirmed that the spatial variation in the properties of DOM from the study region was primarily controlled by diagenesis, highlighting challenges in tracing allochthonous DOM in deeper and more offshore coastal environments.
References
Aguilar-Islas AM, Bruland KW (2006) Dissolved manganese and silicic acid in the Columbia River plume: a major source to the California current and coastal waters off Washington and Oregon. Mar Chem 101(3–4):233–247. doi:10.1016/j.marchem.2006.03.005
Barnes CA, Duxburay C, Morse B (1972) Circulation and selected properties of the Columbia River effluent at sea. In: Pruter AT, Alveison DL (eds) The Columbia River estuary and adjacent ocean waters: bioenvironmental studies. University of Washington Press, Seattle, pp 71–80
Benner R (2003) Molecular indicators of the bioavailability of dissolved organic matter. In: Findlay SEG, Sinsabaugh RL (eds) Aquatic ecosystems: interactivity of dissolved organic matter. Academic, San Diego, pp 121–138
Benner R, Amon RMW (2014) The size-reactivity continuum of major bioelements in the ocean. Annual Review of Marine Science, Volume 7. doi: 10.1146/annurev-marine-010213-135126
Benner R, Louchouarn P, Amon RMW (2005) Terrigenous dissolved organic matter in the Arctic Ocean and its transport to surface and deep waters of the North Atlantic. Global Biogeochem Cy 19. doi: 10.1029/2004gb002398
Blough NV, Zafiriou OC, Bonilla J (1993) Optical-absorption spectra of waters from the Orinoco River outflow—terrestrial input of colored organic matter to the Caribbean. J Geophys Res-Oceans 98(C2):2271–2278. doi:10.1029/92jc02763
Chin YP, Aiken G, Oloughlin E (1994) Molecular-weight, polydispersity, and spectroscopic properties of aquatic humic substances. Environ Sci Technol 28(11):1853–1858. doi:10.1021/Es00060a015
Coble PG (1996) Characterization of marine and terrestrial DOM in seawater using excitation emission matrix spectroscopy. Mar Chem 51(4):325–346. doi:10.1016/0304-4203(95)00062-3
Coble PG (2007) Marine optical biogeochemistry: the chemistry of ocean color. Chem Rev 107(2):402–418. doi:10.1021/Cr050350+
Cory RM, McKnight DM (2005) Fluorescence spectroscopy reveals ubiquitous presence of oxidized and reduced quinones in dissolved organic matter. Environ Sci Technol 39(21):8142–8149. doi:10.1021/Es0506962
Cory RM, Miller MP, McKnight DM, Guerard JJ, Miller PL (2010) Effect of instrument-specific response on the analysis of fulvic acid fluorescence spectra. Limnol Oceanogr-Meth 8:67–78
Determann S, Lobbes JM, Reuter R, Rullkotter J (1998) Ultraviolet fluorescence excitation and emission spectroscopy of marine algae and bacteria. Mar Chem 62(1–2):137–156. doi:10.1016/S0304-4203(98)00026-7
Driscoll CT, Fuller RD, Simone DM (1988) Longitudinal variations in trace metal concentrations in a northern forested ecosystem. J Environ Qual 17:101–107
Fellman JB, Hood E, Edwards RT, D'Amore DV (2009) Changes in the concentration, biodegradability, and fluorescent properties of dissolved organic matter during stormflows in coastal temperate watersheds. J Geophys Res-Biogeo 114. doi: 1021 10.1029/2008jg000790
Fellman JB, Hood E, Spencer RGM (2010) Fluorescence spectroscopy opens new windows into dissolved organic matter dynamics in freshwater ecosystems: a review. Limnol Oceanogr 55(6):2452–2462. doi:10.4319/lo.2010.55.6.2452
Fichot CG, Kaiser K, Hooker SB, Amon RMW, Babin M, Belanger S, Walker SA, Benner R (2013) Pan-Arctic distributions of continental runoff in the Arctic Ocean. Sci Rep-Uk 3. doi: Artn 1053 Doi 10.1038/Srep01053
Fiedler PC, Laurs RM (1990) Variability of the Columbia River Plume observed in visible and infrared satellite imagery. Int J Remote Sens 11(6):999–1010
Guo LD, Santschi PH, Warnken KW (1995) Dynamics of dissolved organic carbon (DOC) in oceanic environments. Limnol Oceanogr 40(8):1392–1403
Guo WD, Stedmon CA, Han YC, Wu F, Yu XX, Hu MH (2007) The conservative and non-conservative behavior of chromophoric dissolved organic matter in Chinese estuarine waters. Mar Chem 107(3):357–366. doi:10.1016/j.marchem.2007.03.006
Helms JR, Stubbins A, Ritchie JD, Minor EC, Kieber DJ, Mopper K (2008) Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnol Oceanogr 53(3):955–969. doi:10.4319/lo.2008.53.3.0955
Hernes PJ, Benner R (2003) Photochemical and microbial degradation of dissolved lignin phenols: Implications for the fate of terrigenous dissolved organic matter in marine environments. J Geophys Res-Oceans 108 (C9). doi: Artn 3291 Doi 10.1029/2002jc001421
Hickey BM (1979) The California current system-hypotheses and facts. Prog Oceanogr 8:191–279
Hickey BM, Pietrafesa LJ, Jay DA, Boicourt WC (1998) The Columbia River Plume study: subtidal variability in the velocity and salinity fields. J Geophys Res-Oceans 103(C5):10339–10368. doi:10.1029/97jc03290
Hill JK, Wheeler PA (2002) Organic carbon and nitrogen in the northern California current system: comparison of offshore, river plume, and coastally upwelled waters. Prog Oceanogr 53(2–4):369–387
Huyer A (1979) Seasonal variation in temperature, salinity, and density over the continental shelf off Oregon. Limnol Oceanogr 22:442–453
Huyer A, Pillsburyr D, Smith RL (1975) Seasonal variation of the alongshore velocity field over the continental shelf off Oregon. Limnol Oceanogr 29:90–95
Jaffé R, McKnight D, Maie N, Cory R, McDowell WH, Campbell JL (2008) Spatial and temporal variations in DOM composition in ecosystems: the importance of long-term monitoring of optical properties. J Geophys Res-Biogeo 113 (G4). doi: 10.1029/2008jg000683
Jorgensen L, Stedmon CA, Granskog MA, Middelboe M (2014) Tracing the long-term microbial production of recalcitrant fluorescent dissolved organic matter in seawater. Geophys Res Lett 41(7):2481–2488. doi:10.1002/2014gl059428
Kirchman DL (2003) The contribution of monomers and other low-molecular weight compounds to the flux of dissolved organic material in aquatic ecosystems. In: Findlay SEG, Sinsabaugh RL (eds) Aquatic ecosystems: interactivity of dissolved organic matter. Academic, San Diego, pp 218–243
Kowalczuk P, Cooper WJ, Whitehead RF, Durako MJ, Sheldon W (2003) Characterization of CDOM in an organic-rich river and surrounding coastal ocean in the South Atlantic Bight. Aquat Sci 65(4):384–401. doi:10.1007/s00027-003-0678-1
Lohan MC, Bruland KW (2006) Importance of vertical mixing for additional sources of nitrate and iron to surface waters of the Columbia River plume: implications for biology. Mar Chem 98(2–4):260–273. doi:10.1016/j.marchem.2005.10.003
Lu YH, Meyers PA, Eadie BJ, Robbins JA (2010) Carbon cycling in Lake Erie during cultural eutrophication over the last century inferred from the stable carbon isotope composition of sediments. J Paleolimnol 43(2):261–272. doi:10.1007/s10933-009-9330-y
Lu YH, Bauer JE, Canuel EA, Yamashita Y, Chambers RM, Jaffé R (2013) Photochemical and microbial alteration of dissolved organic matter in temperate headwater streams associated with different land use. J Geophys Res-Biogeo 118(2):566–580. doi:10.1002/Jgrg.20048
Lu YH, Bauer JE, Canuel EA, Chambers RM, Yamashita Y, Jaffé R, Barrett A (2014) Effects of land use on sources and ages of inorganic and organic carbon in temperate headwater streams. Biogeochemistry 119(1–3):275–292. doi:10.1007/s10533-014-9965-2
McKnight DM, Boyer EW, Westerhoff PK, Doran PT, Kulbe T, Andersen DT (2001) Spectrofluorometric characterization of dissolved organic matter for indication of precursor organic material and aromaticity. Limnol Oceanogr 46(1):38–48
Montgomery S, Lucotte M, Cournoyer L (2000) The use of stable carbon isotopes to evaluate the importance of fine suspended particulate matter in the transfer of methylmercury to biota in boreal flooded environments. Sci Total Environ 261(1–3):33–41. doi:10.1016/S0048-9697(00)00593-3
Mopper K, Schultz CA (1993) Fluorescence as a possible tool for studying the nature and water column distribution of DOC components. Mar Chem 41(1–3):229–238. doi:10.1016/0304-4203(93)90124-7
Murphy KR, Stedmon CA, Waite TD, Ruiz GM (2008) Distinguishing between terrestrial and autochthonous organic matter sources in marine environments using fluorescence spectroscopy. Mar Chem 108(1–2):40–58. doi:10.1016/j.marchem.2007.10.003
Neal VT (1972) Physical aspects of the Columbia River and its estuary. In: Pruter AT, Alveison DL (eds) The Columbia River estuary and adjacent ocean waters: bioenvironmental studies. University of Washington Press, Seattle, pp 19–70
Ogawa H, Tanoue E (2003) Dissolved organic matter in oceanic waters. J Oceanogr 59(2):129–147. doi:10.1023/A:1025528919771
Opsahl S, Benner R (1997) Distribution and cycling of terrigenous dissolved organic matter in the ocean. Nature 386(6624):480–482. doi:10.1038/386480a0
Opsahl SP, Zepp RG (2001) Photochemically-induced alteration of stable carbon isotope ratios (delta C-13) in terrigenous dissolved organic carbon. Geophys Res Lett 28(12):2417–2420
Opsahl S, Benner R, Amon RMW (1999) Major flux of terrigenous dissolved organic matter through the Arctic Ocean. Limnol Oceanogr 44(8):2017–2023
Osburn CL, Stedmon CA (2011) Linking the chemical and optical properties of dissolved organic matter in the Baltic-North Sea transition zone to differentiate three allochthonous inputs. Mar Chem 126(1–4):281–294. doi:10.1016/j.marchem.2011.06.007
Para J, Coble PG, Charriere B, Tedetti M, Fontana C, Sempere R (2010) Fluorescence and absorption properties of chromophoric dissolved organic matter (CDOM) in coastal surface waters of the northwestern Mediterranean Sea, influence of the Rhone River. Biogeosciences 7(12):4083–4103. doi:10.5194/bg-7-4083-2010
Prahl FG, Coble PG (1994) Input and behavior of dissolved organic carbon in the Columbia River Estuary. In Dyer K and Orth R (eds.) Changing particle fluxes in estuaries: implications from science to management, ECSAERF22 Symposium, Olsen & Olsen, Friedensborg, pp. 451–457
Retamal L, Vincent WF, Martineau C, Osburn CL (2007) Comparison of the optical properties of dissolved organic matter in two river-influenced coastal regions of the Canadian Arctic. Estuar Coast Shelf Sci 72(1–2):261–272. doi:10.1016/j.ecss.2006.10.022
Rochelle-Newall EJ, Fisher TR (2002) Chromophoric dissolved organic matter and dissolved organic carbon in Chesapeake Bay. Mar Chem 77(1):23–41. doi:10.1016/S0304-4203(01)00073-1
Romera-Castillo C, Sarmento H, Alvarez-Salgado XA, Gasol JM, Marrase C (2010) Production of chromophoric dissolved organic matter by marine phytoplankton. Limnol Oceanogr 55(1):446–454. doi:10.4319/lo.2010.55.1.0446
Spencer RGM, Aiken GR, Wickland KP, Striegl RG, Hernes PJ (2008) Seasonal and spatial variability in dissolved organic matter quantity and composition from the Yukon River basin, Alaska. Global Biogeochem Cy 22 (4). doi: 10.1029/2008gb003231
Staal M, Thar R, Kuhl M, van Loosdrecht MCM, Wolf G, de Brouwer JFC, Rijstenbil JW (2007) Different carbon isotope fractionation patterns during the development of phototrophic freshwater and marine biofilms. Biogeosciences 4(4):613–626
Stedmon CA, Bro R (2008) Characterizing dissolved organic matter fluorescence with parallel factor analysis: a tutorial. Limnol Oceanogr-Meth 6:572–579
Stedmon CA, Markager S (2005a) Resolving the variability in dissolved organic matter fluorescence in a temperate estuary and its catchment using PARAFAC analysis. Limnol Oceanogr 50(2):686–697
Stedmon CA, Markager S (2005b) Tracing the production and degradation of autochthonous fractions of dissolved organic matter by fluorescence analysis. Limnol Oceanogr 50(5):1415–1426
Stedmon CA, Markager S, Bro R (2003) Tracing dissolved organic matter in aquatic environments using a new approach to fluorescence spectroscopy. Mar Chem 82(3–4):239–254. doi:10.1016/S0304-4203(03)00072-0
Sullivan BE, Prahl FG, Small LF, Covert PA (2001) Seasonality of phytoplankton production in the Columbia River: a natural or anthropogenic pattern? Geochim Cosmochim Ac 65(7):1125–1139. doi:10.1016/S0016-7037(00)00565-2
Sulzberger B, Durisch-Kaiser E (2009) Chemical characterization of dissolved organic matter (DOM): a prerequisite for understanding UV-induced changes of DOM absorption properties and bioavailability. Aquat Sci 71(2):104–126. doi:10.1007/s00027-008-8082-5
Tanaka K, Kuma K, Hamasaki K, Yamashita Y (2014) Accumulation of humic-like fluorescent dissolved organic matter in the Japan Sea. Sci Rep-Uk 4. doi:Artn 5292 Doi 10.1038/Srep05292
Thomas AC, Weatherbee RA (2006) Satellite-measured temporal variability of the Columbia River plume. Remote Sens Environ 100(2):167–178. doi:10.1016/j.rse.2005.10.018
Tyrrell T, Law CS (1997) Low nitrate: phosphate ratios in the global ocean. Nature 387(6635):793–796. doi:10.1038/42915
Walker SA, Amon RMW, Stedmon C, Duan SW, Louchouarn P (2009) The use of PARAFAC modeling to trace terrestrial dissolved organic matter and fingerprint water masses in coastal Canadian Arctic surface waters. J Geophys Res-Biogeo 114. doi:Artn G00f06 Doi 10.1029/2009jg000990
Walker SA, Amon RMW, Stedmon CA (2013) Variations in high-latitude riverine fluorescent dissolved organic matter: a comparison of large Arctic rivers. J Geophys Res-Biogeo 118(4):1689–1702. doi:10.1002/2013jg002320
Weishaar JL, Aiken GR, Bergamaschi BA, Fram MS, Fujii R, Mopper K (2003) Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ Sci Technol 37(20):4702–4708. doi:10.1021/Es030360x
Williamson CE, Zagarese HE (1994) The impact of UV-B radiation on pelagic fresh-water ecosystems. Archive fur Hydrobiologie Beiheft 43:9–11
Wilson HF, Xenopoulos MA (2009) Effects of agricultural land use on the composition of fluvial dissolved organic matter. Nat Geosci 2(1):37–41. doi:10.1038/Ngeo391
Worrall F, Parker A, Rae JE, Johnson AC (1997) A study of the sorption kinetics of isoproturon on soil and subsoil, the role of dissolved organic carbon. Chemosphere 34:87–97
Yamashita Y, Jaffé R (2008) Characterizing the interactions between trace metals and dissolved organic matter using excitation-emission matrix and parallel factor analysis. Environ Sci Tech 42(19):7374–7379
Yamashita Y, Tanoue E (2003) Distribution and alteration of amino acids in bulk DOM along a transect from bay to oceanic waters. Mar Chem 82(3–4):145–160. doi:10.1016/S0304-4203(03)00049-5
Yamashita Y, Jaffé R, Maie N, Tanoue E (2008) Assessing the dynamics of dissolved organic matter (DOM) in coastal environments by excitation emission matrix fluorescence and parallel factor analysis (EEM-PARAFAC). Limnol Oceanogr 53(5):1900–1908. doi:10.4319/lo.2008.53.5.1900
Yamashita Y, Panton A, Mahaffey C, Jaffé R (2011) Assessing the spatial and temporal variability of dissolved organic matter in Liverpool Bay using excitation-emission matrix fluorescence and parallel factor analysis. Ocean Dyn 61(5):569–579. doi:10.1007/s10236-010-0365-4
Acknowledgments
We acknowledge funding support from the Research Grant Committee Grant from the University of Alabama (to Y. H. L.) and the Undergraduate Research Grant from the University of Alabama (to Y. H. L. and A. J.), in collaboration with National Science Foundation (NSF) support EAR-1255724 to Y. H. L. We thank participating scientists and supporting crew of UNOLS Chief Scientist Training cruise W1106A on the R/V Wecoma. Special thanks go to Drs. Clare Reimers and Patricia Wheeler for acting as the cruise mentors, Daryl Swensen for providing technical support at sea, and Dr. Alyson Santoro for processing the CTD data. The cruise and training program were supported by the NSF through grant OCE-1125396. We thank the two anonymous reviewers for their thoughtful and helpful comments which helped us to improve this contribution.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Dieter Wolf-Gladrow
Rights and permissions
About this article

Cite this article
Lu, Y., Edmonds, J.W., Yamashita, Y. et al. Spatial variation in the origin and reactivity of dissolved organic matter in Oregon-Washington coastal waters. Ocean Dynamics 65, 17–32 (2015). https://doi.org/10.1007/s10236-014-0793-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10236-014-0793-7