
Abstract
Tropical forests are the most carbon (C)-rich ecosystems on Earth, containing 25–40% of global terrestrial C stocks. While large-scale quantification of aboveground biomass in tropical forests has improved recently, soil C dynamics remain one of the largest sources of uncertainty in Earth system models, which inhibits our ability to predict future climate. Globally, soil texture and climate predict ≤ 30% of the variation in soil C stocks, so ecosystem models often predict soil C using measures of aboveground plant growth. However, this approach can underestimate tropical soil C stocks, and has proven inaccurate when compared with data for soil C in data-rich northern ecosystems. By quantifying soil organic C stocks to 1 m depth for 48 humid tropical forest plots across gradients of rainfall and soil fertility in Panama, we show that soil C does not correlate with common predictors used in models, such as plant biomass or litter production. Instead, a structural equation model including base cations, soil clay content, and rainfall as exogenous factors and root biomass as an endogenous factor predicted nearly 50% of the variation in tropical soil C stocks, indicating a strong indirect effect of base cation availability on tropical soil C storage. Including soil base cations in C cycle models, and thus emphasizing mechanistic links among nutrients, root biomass, and soil C stocks, will improve prediction of climate-soil feedbacks in tropical forests.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Tropical forest soils contain some of the largest stocks of carbon (C) on Earth, making them of broad importance in the global C cycle (Field et al. 1998; Jobbagy and Jackson 2000). Nonetheless, our understanding of what drives variation in soil C stocks across tropical landscapes is limited, inhibiting our ability to predict large-scale responses to global change (IPCC 2013). Aboveground C stocks and litterfall in closed-canopy tropical forests can be measured and mapped with precision at large scales using remote sensing as was recently done for Panamanian forests (Asner et al. 2013), so extrapolating soil C stocks from aboveground biomass and plant growth measures with few in situ soil data has formed the basis for many modeling efforts (Luo et al. 2016).
Plants may contribute organic matter to soil C pools either via litterfall, which would be most linked to aboveground biomass measures, or via belowground root turnover and exudation. Large-scale manipulations of plant litterfall, as may happen with shifts in net primary production, have resulted in changes in soil C stocks at the soil surface in tropical forests (Tanner et al. 2016). However, a large proportion of surface litter C is likely respired back to the atmosphere as CO2 rather than transported downward and stored in mineral soils. Also, increased litter may even promote a priming effect and losses of soil C, as suggested by studies in forests on strongly weathered soils in Panama and subtropical China (Liu et al. 2017; Sayer et al. 2011). Rather than originating from aboveground plant tissues, most long-term C storage within mineral soils probably originates from root biomass (Rasse et al. 2005). Detailed studies using isotopically labeled root and shoot tissues have confirmed slower decomposition of buried root material versus surface litter in a ponderosa pine forest (Bird and Torn 2006), and greater retention of material placed into the A horizon versus the O horizon on decadal timescales (Pries et al. 2017), suggesting that both the chemical composition of roots, and their location directly in mineral soil can make root contributions a dominant portion of soil C. Thus, estimates of root biomass, rather than aboveground C stocks, are likely to be more useful for predicting soil C stocks. However, tropical forest root biomass is notoriously difficult to quantify, spatially variable, and is not easily predictable as a proportion of aboveground biomass (Waring and Powers 2017).
An alternative and widely-used approach for predicting soil C stocks at broad scales relies on commonly measured abiotic properties, particularly climate and soil texture, with climate likely most influencing surface soil C stocks, and clay concentrations influencing soil C stocks at depth (Jobbagy and Jackson 2000; Post et al. 1982). For example, global-scale data show that soil C stocks are generally greater in wetter forests (Post et al. 1982). A relationship between rainfall and soil C storage within tropical forests would be of particular interest in the context of regional drying that is expected with climate change (Cusack et al. 2016). Among soil characteristics, clay content has been identified as a relatively strong predictor of soil C stocks across biomes (Jobbagy and Jackson 2000), in part because sorption of organic matter to charged mineral surfaces can suppress degradation by microbial decomposers (Sollins et al. 1996). An advantage of using abiotic properties like clay content to predict soil C stocks is that data for soil texture are widely available, such as from the World Soil Information Service (Batjes et al. 2017), whereas root biomass data are more scarce.
An improved approach for predicting tropical soil C stocks may be to incorporate metrics that are strongly correlated to root biomass into the existing abiotic framework. In particular, soil phosphorus (P) and base cation availability are commonly shown to limit plant growth in highly weathered tropical soils, which are leached of rock-derived nutrients and rich in nitrogen (N) (Crews et al. 1995; Harrington et al. 2001; Vitousek and Sanford 1986). Root biomass in particular has been inversely related to soil P and/or base cation availability across a range of distinct tropical forests (Espeleta and Clark 2007; Maycock and Congdon 2000). Similarly, root biomass and root growth in highly weathered tropical soils have shown consistent declines in response to elevated potassium (K) over 15 years of factorial nutrient fertilizations in Panama (Wurzburger and Wright 2015; Yavitt et al. 2011). These landscape-scale and experimental declines in root biomass in response to increased availability of rock-derived nutrients likely indicate decreased plant C allocation to root biomass when nutrients are plentiful (Bloom et al. 1985), which in turn reduces C inputs to soils. Root exudates are also increasingly recognized as an important contribution of root C to belowground C stocks. Root exudates are produced to promote the release of mineral nutrients via stimulated microbial decomposition and/or direct solubilization (Phillips et al. 2011). Increases in rock-derived nutrient availability in tropical soils appear to suppress root exudates (Nottingham et al. 2012), similar to trends for root biomass, further decreasing root inputs to belowground C stocks. Thus, rock-derived nutrient content in soil might be a useful indicator of plant allocation of biomass to roots in tropical forests, so incorporating base cations into predictive models could improve predictions of soil C stocks.
We hypothesized that soil C stocks in tropical forests correspond to root biomass and soil nutrients, not aboveground plant measures. Specifically, we hypothesized that root biomass and the availability of rock-derived nutrients are the strongest predictors soil C stocks across tropical forests.
Materials and methods
Study sites
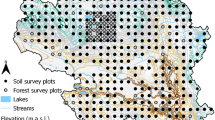
We studied 48 1-ha plots in lowland tropical forest sites across the Isthmus of Panama to assess factors driving soil C stocks in tropical forests (Fig. 1). All plots were in lowland tropical forests (elevation 10–410 meters above sea level) and included secondary, mature secondary, and old growth primary forests (Pyke et al. 2001). The plots spanned a distinct rainfall gradient of ~ 1750 mm year−1 mean annual precipitation (MAP) on the Pacific coast, to ~ 4000 mm year−1 MAP on the Caribbean coast (Engelbrecht et al. 2007; Pyke et al. 2001). The forests across the Isthmus have a tropical monsoon climate. The wetter Caribbean coast has a shorter dry season (~ 115 days), compared with the drier Pacific coast (~ 150 days). The mean annual temperature across sites is 26 °C and mean monthly temperature varies by < 1 °C during the year (Windsor et al. 1990).

A map of the extensive geological variability of central Panama is shown, with plots spanning from the drier southern Pacific coast to the wetter northern Caribbean coast. Colors represent different geological substrates and formations (Fm). The 48 sites used in this study are identified with letters, with site-level geological details, soil chemistry, and within-site variation given in Online Appendix 1. Litterfall data in this study are from sites A, E, F, J, R, NN, SS, and VV. Measured soil C stocks are indicated for each site in bold numbers (kg-C m−2), and site markers indicate sites with low, medium, and high C stocks. Maps modified from Stewart et al. (1980)
The soils at our sites developed on a range of geological substrates across the rainfall gradient (Fig. 1, Online Appendix 1) (Pyke et al. 2001; Stewart et al. 1980; Turner and Engelbrecht 2011; Woodring 1958), including volcanic (basalt, andesite, agglomerate, rhyolitic tuff) and marine sedimentary (limestone, calcareous sandstone, siltstone, mudstone) lithologies. As a result, soils have marked variation in fertility (Condit et al. 2013; Turner and Engelbrecht 2011), which spans a range of taxonomic orders (Turner and Engelbrecht 2011). Soils were classified by digging a profile pit adjacent to the southwest corner of each plot. Profiles were described by genetic horizon and samples of horizons were collected for chemical and physical analysis (Table 1). Soil properties in general across these sites are weakly or not at all correlated with rainfall (Turner and Engelbrecht 2011), providing an opportunity to isolate the influence of rainfall from soil properties.
Soil and root sampling and analyses
We quantified total soil C, N, and P, resin-extractable P, extractable base cations, pH, soil texture and bulk density to 1 m depth in 1-ha plots at each site using sample from within plots and larger soil pits outside the edge of each plot. Each 1 ha plot is marked on a 20 m × 20 m grid. Soil cores were taken to 1 m depths in quadrats in the four corners and the center of each 1 ha plot (subsample n = 5), in increments of 0–10 cm, 10–20 cm, 20–50 cm, and 50–100 cm. The 0–10 cm and 10–20 cm samples were taken with a constant volume corer (5 cm diameter), and deeper samples were taken with a 6.25 cm diameter auger. To account for greater variation in soil roots and C stocks at the surface, 8 additional soil samples from 0–10 cm depths were collected from alternate 20 × 20 m quadrats using the constant volume corer (i.e. 13 surface samples in total per plot). Stocks for fine roots and C were calculated using the average values at each depth, summed to 1 m. This approach yielded 28 individual samples per 1 ha plot, which were each analyzed for soil properties separately. In the two larger plots (BCI—50 ha, Sherman crane site—5.96 ha) additional surface samples were collected, yielding a total of 50 samples on BCI (including samples to 100 cm in eight locations) and 44 samples at Sherman crane (including samples to 100 cm in five locations). Within-plot variation for root biomass and soil chemistry is reported as the standard error of the 5 locations to 1 m depth with plots (Online Appendix 1). The within-plot relative standard error (i.e., standard error as a percentage of the mean) average 19% for root biomass to 1 m, and 13% for soil C stocks to 1 m across plots (Online Appendix 1). Including the additional surface soil samples for 0–10 cm depths decreased these within-plot relative standard errors to 14% for surface roots, and 6% for surface soil C stocks at this depth. These additional measures were included in plot-scale averages for surface soils prior to summing by depth. Larger soil pits (~ 1.5 m3) were dug outside of the edges of each plot for soil classification, bulk density, and texture measures.
Samples from plots were collected during the approximately eight-month wet season to avoid seasonal changes in soil C concentrations (Turner et al. 2015), which could confound comparison among sites. The majority of the plots were sampled during 2008 and 2009, with additional plots sampled between 2010 and 2014. Samples were returned to the laboratory within 2 h of sampling and initially screened (< 9 mm) to break up large aggregates, and stones were removed by hand. Soils were not sieved < 2 mm when fresh, because this is impractical for clay-rich soils.
Fine roots (< 2 mm diameter) was sorted out of soils by hand. Live fine roots were exhaustively removed from each sample (i.e. until no more visible roots remained), washed on a 250 μm sieve, oven-dried for 3 days until weight stabilized at 60 °C, and weighed. Live fine root biomass stocks were then calculated on an area basis (per m2) using the core volume and depth for each sample, averaging depths across the plot and summing the depths to 50 cm and to 100 cm.
Extractable P and base cations were measured on field-fresh soils as an indication of plant-available nutrients. Fresh soils from plots were extracted immediately using resin exchange beads for available P (Turner and Romero 2009). Base cations were extracted on fresh soils within 24 h in Mehlich-3 solution (Mehlich 1984), with detection by inductively coupled plasma optical emission spectrometry (Optima 7300 DV, Perkin Elmer, Inc., Shelton, CT). Total exchangeable bases (TEB) was calculated as the sum of Ca, K, and Mg.
Total nutrient content for C, N and P was then measured on dry soils. Subsamples were air-dried for 10 days at ambient laboratory temperature, sieved < 2 mm to isolate the fine earth fraction, and ground in a ball mill. Total C and N were determined by combustion and gas chromatography using a Thermo Flash NC1112 Soil Analyzer (CE Elantech, Lakewood, NJ), and soil pH was determined in a 1:2 soil to deionized water mixture using a glass electrode. Total P was determined on dried soils by ignition (550 °C, 1 h) and then extraction in 1 M H2SO4 (16 h, 1:50 soil to solution ratio) (Walker and Adams 1958), with P detection by automated neutralization and molybdate colorimetry on a Lachat Quickchem 8500 (Hach Ltd, Loveland, CO, USA).
Soils were classified in profile pits outside of plots according to Soil Taxonomy (Staff 1999), which provides an indication of soil weathering status. The concentrations of sand (0.053–2.0 mm), silt (0.002–0.053 mm), and clay (< 0.002 mm) size particles were determined by the pipette method after pretreatment to remove salts (sodium acetate extraction), organic matter (H2O2 oxidation), and iron oxides (dithionite reduction) (Gee and Or 2002). In profile pits, bulk density was determined by the excavation method (Grossman and Reinsch 2002), removing ~ 1 L of soil and measuring the excavated volume of the plastic-lined hole with water. For the upper 20 cm of the soil, bulk density was calculated by weighing the total fresh soil mass from the constant volume core samples, and converting this to dry mass using oven dry weight (calculated by drying a subsample for 24 h at 105 °C), and accounting for stone content of the samples.
Total C stocks were calculated in kg/m2 using average C concentrations from within-plot samples, bulk density from profile pits, and depth increments, summing to 1 m depths. Bulk densities were measured for the fine earth fraction as a proportion of the total soil volume—i.e. accounting for the stone fraction (in both core samples and profile pits). Where samples were obtained to < 100 cm due to bedrock, the plot values were corrected based on the proportion of cores in the plot reaching a particular depth (i.e. average soil depth for the plot). However, plot averages were not corrected for floating boulders, where augering was prevented in some locations in the plot but soil continued below. Further details on the plots and sampling of vegetation and soils have been reported previously (Condit et al. 2013; Engelbrecht et al. 2007; Pyke et al. 2001; Turner and Engelbrecht 2011).
Aboveground biomass and litterfall production
Aboveground dry biomass (AGB) for all trees > 10 cm diameter at breast height was estimated in each of the plots using allometric equations relating volume to stem diameter along with species-specific wood density. Methodological details and examination of errors have been published (Chave et al. 2004), but most measurements have been repeated and methods updated for this paper between 2000 and 2010. Litterfall biomass was collected at a subset of 8 sites biweekly for 1 year from 2013–2014. Fifty 1 m × 1 m litter traps were established within each of 8 1 ha forest plots. This subset of the 48 plots represented both high and low fertility sites within a given rainfall range. Litter traps were distributed evenly across each plot, with a pair of traps in the center of each of the twenty-five 20 × 20 m quadrats. Litter from each trap was collected biweekly and dried at 65 °C until mass stabilized (~ 48 h), after which dry mass was measured.
Statistical analyses
To identify significant predictors of soil C stocks we used preliminary backward stepwise linear regression analyses including all soil data for order, roots, nutrients, and texture, as well as aboveground biomass, and rainfall as factors. Our stepwise model selection process sought to minimize Bayesian information criterion (BIC) values. This approach identifies significant factors and provides a penalty for adding parameters (Schwarz 1978), resulting in the best minimum adequate general linear model. We then ran the identified model including only potentially significant factors identified by these stepwise regressions in final linear models. Similar general linear model selection was used for predicting fine root biomass. We then ran post hoc means separation Fisher’s Least Significant Difference (LSD) tests to compare soil C, nutrients, texture, and root biomass among soil orders. Where necessary, values were log transformed to meet assumptions of normality (e.g. total P and resin-extractable P).
We also analyzed the above relationships using structural equation modeling (SEM) path analysis to statistically assess the goodness of fit of our hypothesized relationships among factors influencing soil C stocks. We used soil chemical and physical factors, aboveground biomass, and rainfall as exogenous factors, and soil C and root biomass stocks as endogenous factors. We used SEM modification indices to add or remove specific pathways from the model. An initial model included soil P as an exogenous factor, but this was removed because of poor model similarity to the saturated model. The final default model (i.e. our model) was tested for overall similarity to the saturated model (i.e., ideal model) using common metrics of comparison (i.e., no significant difference from saturated model, Chi-square p > 0.05, GFI > 0.9, NFI > 0.9, RMSEA < 0.05). The model used 48 observations.
General linear models were performed using 13.0.0 JMP Pro software (SAS Institute Inc., 2016), and SEM path analysis was performed using SPSS AMOS 23 software (IBM, 2015). Statistical significance for all relationships and models was p < 0.05, and means are reported ± one standard error.
Results
Soil carbon stocks across environmental gradients
Overall, soil C stocks to 1 m depth ranged between 7.3 and 20.3 kg C m−2 across the sites, with an overall average of 13.3 ± 0.5 kg C m−2. The top 50 cm of soil contained an average of 75 ± 1% of soil C across sites, with a greater content of C near the surface in less-weathered versus strongly weathered soil orders (Table 1). There was an even greater content of root biomass in the top 50 cm of soil (93 ± 0.6%), with no variation among soil orders (Table 1).
In addition to the nearly three-fold change in soil C stocks, the sites used here covered five soil orders (Table 1), over 25 geological formations (Fig. 1), a two-fold range in rainfall, a 20-fold range in base cations, and a > 100-fold range in available P (i.e., resin-extractable, Online Appendix 1). Thus, although the data come from a relatively restricted geographic region, the diversity of environmental conditions means that the results are likely to be broadly applicable over much larger geographical ranges. The more strongly weathered soils were poorer in rock-derived nutrients, more acidic, and contained greater fine root biomass and soil C stocks relative to less weathered soil orders (Table 1). However, most soil characteristics were not clearly distinct among the orders (Table 1).
Predictors of tropical soil carbon stocks
Across the 48 diverse tropical forest sites, aboveground biomass and litterfall biomass were not significant factors for predicting soil C stocks using the stepwise general linear model selection process, which initially included all soil, aboveground, and climate data. Neither aboveground biomass nor litterfall biomass had a significant univariate correlation with soil C stocks (Figs. 2a and S2a). Instead, soil clay content, fine root biomass, and precipitation were significant factors in the minimum adequate general linear model for predicting soil C stocks to 1 m (R 2 = 0.45, p < 0.001, BIC = 220, Online Appendix 3a). Soil clay content (Fig. 2b) and fine root biomass (Fig. 2c) had the strongest univariate relationships with soil C stocks to 1 m, and also explained the majority of the variation in the minimum adequate general linear model (Online Appendix 3a). Mean annual precipitation alone was only a weak positive predictor of soil C stocks (R 2 = 0.10, n = 45, p < 0.05; Fig. 3), despite the nearly two-fold difference in rainfall across the sites. Predictors of soil C stocks to 0.5 m were similar, except that soil clay content was not a significant factor, and root biomass explained an even larger portion of the variation in the minimum adequate general linear model (Online Appendix 3b). Soil nutrients did not emerge as significant factors in the general linear model selection process for predicting soil C stocks.

Comparing predictors of soil C stocks, a above ground biomass showed no correlation (R 2 = 0.009, n = 43, p = 0.52), whereas significant correlations were found for b soil clay content (R 2 = 0.20, n = 47, p < 0.05), and c live fine root biomass (R 2 = 0.27, n = 44, p < 0.05). d Live fine roots, in turn, showed a logarithmic decline with extractable base cations (R 2 = 0.21, n = 44, p < 0.05). Excluding the three sites with very low base cations and large fine root biomass still resulted in a significant negative linear relationship between total extractable bases and fine root biomass (dashed line, R 2 = 0.12, n = 41, p < 0.05). Equations for the significant predictors are: b Carbon Stock = 8 + 0.09 * Clay %; c Carbon Stock = 9 + 0.01 * Fine Root Biomass; d Roots biomass = 307– [47 * log (total extractable bases)]

Soil C stocks were weakly but significantly correlated with rainfall across the rainfall and soil fertility gradient in Panama (R 2 = 0.11, n = 48, p < 0.05)
A similar model selection process indicated that live fine root biomass to 1 m depth was most strongly predicted by total extractable base cations alone (BIC = 549), showing a logarithmic decline (Fig. 2d). This relationship between fine root biomass and total base cations was driven by three sites on highly weathered Ultisols with very low base cations. Even excluding those sites, however, there was a significant linear negative relationship between total extractable bases and fine root biomass (Fig. 2d, R 2 = 0.12, n = 41, p < 0.05). There was no significant relationship between root biomass and rainfall. Thus, roots were one of the strongest positive predictors of soil C stocks, and roots in turn were best predicted by total extractable base cations, with a negative logarithmic relationship.
As might be expected from the two sets of regression analyses described above (predictors of soil C, followed by predictors of fine root biomass), there were also significant negative relationships for soil C stocks with extractable calcium (Ca), extractable magnesium (Mg), and total extractable base cations (R 2 = 0.14, 0.14, and 0.16, respectively, p < 0.01), but these relationships were weak compared with the predictors of soil C stocks identified through the stepwise model building process. Adding base cations to the general linear model for predicting soil C stocks raised the BIC value of the model slightly (BIC = 223), indicating no improvement in the overall model.
Structural equation modeling was then used to infer mechanistic and directional relationships among all of the factors, and particularly to explore indirect linkages between soil nutrients and soil C stocks. The most parsimonious structural equation model for the data included soil base cations, clay content, and rainfall as exogenous factors, and fine root biomass as an endogenous factor linking base cations to soil C stocks (Fig. 4). Thus, the influence of base cations on soil C stocks was mediated via effects on root biomass. This model explained nearly 50% of the variation in soil C stocks across the 48 sites.

A structural equation model (SEM) path analysis for the relationships between exogenous factors (total extractable bases to 1m kg/m2 , rainfall MAP mm, and clay content %), and endogenous factors (fine root biomass to 1m g/m2 , and soil organic C stock to 1m kg/m2) to 1 m soil depth is shown. Bold values in italics above each endogenous factor show the squared multiple correlation for that factor, giving the amount of variation in that factor accounted for by all of its predictors (i.e., R 2). The sign below each arrow shows the direction of the relationship, all of which are significant (p < 0.05). Correlations among exogenous factors were not significant in this analysis. Factors in rectangles represent measured values, and circles represent unmeasured error terms (e1 and e2) for each endogenous factor. Our model was not significantly different (p = 0.19) from the ideal saturated model (χ2 = 8.7, df = 6), indicating a good fit for the data
Rainfall was a relatively strong predictor of cumulative annual litterfall (Online Appendix 2b), and particularly of dry season litterfall (Online Appendix 2c), showing significant negative relationships. Thus, rainfall was a strong predictor of aboveground C fluxes from plants to the forest floor, but not of belowground C stocks.
Discussion
In general, our results do not support the use of aboveground plant productivity metrics for predicting tropical soil C stocks. Use of aboveground plant metrics to simulate soil C has been shown to increase variation in model predictions of soil C stocks in general, and there are few modeling cases that have done this for lowland tropical forests (Todd-Brown et al. 2013).
Our average value of 13.3 kg C m−2 for soil C stocks across these Panama sites is between published averages of 11.5 kg C m−2 (Post et al. 1982) and 15.8 kg C m−2 (Jobbagy and Jackson 2000) to 1 m depths for similar tropical forest types globally. Our range in soil C stocks (7.3–20.3 kg C m−2) is also similar to ranges published for Costa Rican forests (5–14 kg C m−2) on volcanic soils across an elevation gradients to 30 cm depth (Powers and Schlesinger 2002) and Amazonia forests (3–14 kg C m−2) across five strongly weathered soils to 2 m depth (Telles et al. 2003).
Rainfall and soil carbon storage
Somewhat surprisingly, soil C stocks were only weakly correlated with precipitation across these sites, despite a nearly two-fold change in rainfall over the gradient, and despite a significant negative relationship between rainfall and litterfall biomass (i.e. aboveground plant inputs to soil C). Trends in soil C stocks across rainfall gradients can be related to how microbial decomposition responds to rainfall. A study of five lowland Neotropical sites with between 508 and 4100 mm MAP showed a relatively linear increase in decomposition rates of standard leaf tissues with moisture, and increased decomposition rates of native leaf litters up to 5500 mm MAP (Cusack et al. 2009). At the very wet end of the rainfall spectrum decomposition may be inhibited by low oxygen availability (McGroddy and Silver 2000). For example, a Hawaiian study across a montane rainfall gradient of 2000–5000 mm MAP showed a linear decline in decomposition rates with rainfall (Schuur 2001). In contrast to these significant relationships between rainfall and decomposition, a study along the Panamanian rainfall gradient sites from 1900 to 2700 mm MAP showed no relationship between rainfall and leaf litter decomposition rates during the wet season (Dale et al. 2015), similar to our weak relationship between soil C stocks and rainfall. Thus, the Panama rainfall gradient may fall within a moderate range of rainfall where decomposition rates, and thus soil C losses, are not negatively affected at either the wet or the dry end of the gradient. These results indicate that surface litterfall dynamics do not have a substantial influence on soil C stocks to 1 m depths in these forests, and this may be the reason that rainfall is decoupled from soil C stocks.
Clay concentrations and soil C storage
Compared with the weak precipitation effect, we found a somewhat stronger univariate relationship between soil clay concentrations and soil C stocks (R 2 = 0.20, p < 0.05), similar to global-scale coefficients of determination (Jobbagy and Jackson 2000). Similarly, clay concentrations were the best predictor of soil C stocks to 2 m depths across five Amazonian lowland forest sites on highly weathered soils (Telles et al. 2003).
While clay concentration appears to be a reliable predictor of soil C stocks across soil types, more detailed mineralogical information can provide much greater predictive power within sites, largely because the formation of organo-mineral associations varies among mineralogies (Oades 1988). For example, aluminum in organo-mineral complexes, as an indicator of Al-humus association, was a strong predictor (R 2 = 0.75) of soil C to 30 cm depths across 26 lowland Costa Rican rainforest sites, whereas higher elevation sites were predicted by other mineralogical properties (Powers and Schlesinger 2002). Data for a Hawaiian soil climosequence showed strong relationships between soil C stocks and soil mineralogy, with short range order (i.e., non-crystalline) mineral content explaining half of the variation in soil C stocks to 1.5 m depth (Torn et al. 1997), and different biomolecular groups of SOC responding similarly positively to short range order mineral content (Cusack et al. 2013). In highly weathered Puerto Rican soils, in contrast, iron minerals were more important than short range order minerals in driving soil C stocks, likely via non-sorptive mechanisms (Coward et al. 2017). Thus, different mineralogical characteristics of tropical clays influence soil C stocks in different sites, such that a broader measure of clay concentration is more likely to be predictive across sites with markedly contrasting soil properties, as in the current study. Given the widespread use and availability of data on soil texture, the relationship shown here for clay concentrations versus tropical soil C stocks across soil orders is broadly useful.
Fine root contributions to soil C storage
Our correlative evidence suggest that root biomass is the main contributors to soil C stocks for these forests, following broader-scale patterns (Rasse et al. 2005). The synthesis by Rasse et al. (2005) suggested that the tough, lignin-rich tissue chemistry of roots, as well as increased physio-chemical protection of organic matter senesced directly into mineral soils, help explain the greater residence time of root-derived C versus shoot-derived C in soils.
For our sites, fine root biomass was insensitive to rainfall, helping explain why soil C stocks were also insensitive to rainfall. Five years of irrigation at a site near the middle of the Panama rainfall gradient did not change root biomass, rooting depth, or root turnover rates (Yavitt and Wright 2001), indicating a general and a site-specific insensitivity to changes in moisture for fine root biomass in these forests. In contrast to our results, a study in seasonal Amazonian forests across soil fertility gradients showed decreased root biomass with declining soil moisture, both between sites due to seasonality, and within sites due to experimental drying (Metcalfe et al. 2008). These contrasting results across observational and manipulative studies indicate that the potential effects of climatic drying on root activity, and links to soil C storage, merit further mechanistic research in tropical forests.
Base cation availability as a control on soil carbon via fine roots
Base cation availability emerged here as the strongest predictor of tropical forest live fine root biomass. These results suggest that there was greater plant investment of C into fine root biomass in cation-poor soils, which would exemplify a common plant strategy of allocating greater growth belowground in nutrient-scarce sites to maximize acquisition (Bloom et al. 1985). Interestingly, no single rock-derived nutrient was strongly correlated with root biomass or soil C stocks across the sites, suggesting that different individual cations may have driven fine root production across the variation in geological substrates and soil types. For example, there was a strong effect of K fertilization on root dynamics at sites near the middle of this rainfall gradient (Wright et al. 2011; Wurzburger and Wright 2015; Yavitt et al. 2011), but detailed studies of cation effects on root dynamics from drier and wetter sites are lacking. It was somewhat surprising that available P did not emerge as an important driver of root biomass or soil C stocks, since resin-extractable P content varied so strongly among these sites, and available P has been shown to limit aboveground plant growth in some tropical forests on highly weathered soils (Harrington et al. 2001; Vitousek and Sanford 1986). Our results, together with the experimental results for K at the middle of the rainfall gradient, indicate that base cations likely provide an important control on root biomass production in these tropical forests, but we need a better understanding of which rock-derived nutrients are linked to soil C storage in different tropical soils.
In addition to influencing fine root biomass, base cations may also affect the transfer of roots into mineral soil C pools via effects on root decomposition rates. For example, root Ca concentrations correlated positively to root decomposition rates in a review of 175 global values (Silver and Miya 2001). In contrast, a recent study at two sites in Panama showed that root decomposition rates correlated negatively with root K and Ca content (Guerrero-Ramirez et al. 2016). The relationships among soil base cation availability, root tissue chemistry, root turnover, and root decomposition rates should be explored further at these sites to better understand the direct and indirect effects of base cations on soil C stocks.
There are some alternate explanations for the linkages we observed among soil base cations, fine roots, and soil C stocks. First, polyvalent cations like exchangeable Ca2+ can directly increase soil C storage via cation bridging to clay particles (Oades 1988). This would represent a direct abiotic effect of cations on soil C stocks, rather than the indirect biological effect posited here. Because we did not see strong direct correlations between any single cation and soil C stocks, however, we do not believe that the importance of cations for C storage in these soils is primarily abiotic or direct. Second, the direction of causality between root biomass and cation availability might be the opposite of that shown in our structural equation model. In a Costa Rican forest, different canopy tree species had varying effects on soil pH, thus altering dispersion of organo-mineral colloids, and thereby releasing cations from pedogenic structures into available pools (Russell et al. 2017). It is possible that across our Panamanian forests, which vary markedly in canopy species composition, there is a site-level effect of root chemistry on soil pH and thus on cation cycling. However, we did not observe a significant univariate relationship between root biomass and soil pH across sites. There were significant differences in soil pH and extractable base cations among soil orders, as would be expected. Thus, we think that our designation of extractable base cations as an exogenous factor in the structural equation model, rather than an endogenous factor affected by root biomass, is correct. While our data support the structural relationships presented in our model, further research is merited to better understand the mechanisms by which base cation availability may drive down root biomass (e.g. changes in plant allocation and/or root turnover rates), and the mechanisms by which root biomass is transferred into soil C stocks (e.g. via exudates, turnover, and/or incorporation into different soil C fractions).
The apparent importance of base cation availability in driving live root biomass stocks, and thus indirectly driving soil C stocks, should be integrated into the tropical ecosystem models predicting future C storage. Data on soil cations, as well as texture, are available for many international sites (Batjes et al. 2017), and can be estimated from geological information and rainfall where data is unavailable (Augusto et al. 2017; Turner and Engelbrecht 2011).
Conclusions
This work illustrates a decoupling between aboveground plant biomass and soil C stocks in these lowland tropical forests. Live fine roots, rather than aboveground plant growth or litterfall, appeared to provide the primary source of C to tropical soil stocks to 1 m depths. Soil clay content and precipitation were also significant predictors of soil C stocks, and base cation availability appeared to provide an important indirect effect on soil C stocks via a relationship with fine root biomass. These results suggest that responses of tropical soil C stocks to global change will depend on how fine root dynamics are altered across fertility gradients.
References
Asner G, Mascaro J, Anderson C, Knapp D, Martin R, Kennedy-Bowdoin T, van Breugel M, Davies S, Hall J, Muller-Landau H, Potvin C, Sousa W, Wright J, Bermingham E (2013) High-fidelity national carbon mapping for resource management and REDD+. Carbon Balance Manage 8(7). https://doi.org/10.1186/1750-0680-1188-1187
Augusto L, Achat D, Jonard M, Vidal D, Ringeval B (2017) Soil parent material—a major driver of plant nutrient limitations in terrestrial ecosystems. Glob Change Biol 23(9):3808–3824. https://doi.org/10.1111/gcb.13691
Batjes NH, Ribeiro E, van Oostrum A, Leenaars J, Hengl T, de Jesus JM (2017) WoSIS: providing standardised soil profile data for the world. Earth Syst Sci Data 9(1):1–14
Bird JA, Torn MS (2006) Fine roots vs. needles: a comparison of C-13 and N-15 dynamics in a ponderosa pine forest soil. Biogeochemistry 79(3):361–382
Bloom AJ, Chapin FS, Mooney HA (1985) Resource limitation in plants—an economic analogy. Annu Rev Ecol Syst 16:363–392
Chave J, Condit R, Aguilar S, Hernandez A, Lao S, Perez R (2004) Error propagation and scaling for tropical forest biomass estimates. Philos Trans R Soc Lond Ser B-Biol Sci 359(1443):409–420
Condit R, Engelbrecht BMJ, Pino D, Perez R, Turner BL (2013) Species distributions in response to individual soil nutrients and seasonal drought across a community of tropical trees. Proc Natl Acad Sci USA 110(13):5064–5068
Coward EK, Thompson AT, Plante AF (2017) Iron-mediated mineralogical control of organic matter accumulation in tropical soils. Geoderma 306:206–216
Crews TE, Kitayama K, Fownes JH, Riley RH, Herbert DA, Mueller-Dombois D, Vitousek PM (1995) Changes in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii. Ecology 76(5):1407
Cusack DF, Chou WW, Yang WH, Harmon ME, Silver WL, Lidet T (2009) Controls on long-term root and leaf litter decomposition in neotropical forests. Glob Change Biol 15(5):1339–1355
Cusack DF, Chadwick OA, Hockaday WC, Vitousek PM (2013) Mineralogical controls on soil black carbon preservation. Glob Biogeochem Cycle 26(2):GB2019
Cusack D, Karpman J, Ashdown D, Cao Q, Ciochina M, Halterman S, Lydon S, Neupane A (2016) Global change effects on humid tropical forests: evidence for biogeochemical and biodiversity shifts at an ecosystem scale. Rev Geophys 54(3):523–610. https://doi.org/10.1002/2015RG000510
Dale SE, Turner BL, Bardgett RD (2015) Isolating the effects of precipitation, soil conditions, and litter quality on leaf litter decomposition in lowland tropical forests. Plant Soil 394(1–2):225–238
Engelbrecht BMJ, Comita LS, Condit R, Kursar TA, Tyree MT, Turner BL, Hubbell SP (2007) Drought sensitivity shapes species distribution patterns in tropical forests. Nature 447(7140):80–U82
Espeleta JF, Clark DA (2007) Multi-scale variation in fine-root biomass in a tropical rain forest: a seven-year study. Ecol Monogr 77(3):377–404
Field CB, Behrenfeld MJ, Randerson JT, Falkowski P (1998) Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281(5374):237–240
Gee G, Or D (2002) Particle size analysis. In: Dane J, Topp C (eds) Methods of soil analysis, part 4—physical methods. Soil Science Society of America, Wisconsin, pp 255–293
Grossman R, Reinsch T (2002) Bulk density. In: Dane J, Topp C (eds) Methods of soil analysis. Part 4. Physical methods. Soil Society of America, Wisconsin, pp 201–293
Guerrero-Ramirez NR, Craven D, Messier C, Potvin C, Turner BL, Handa IT (2016) Root quality and decomposition environment, but not tree species richness, drive root decomposition in tropical forests. Plant Soil 404(1–2):125–139
Harrington RA, Fownes JH, Vitousek PM (2001) Production and resource use efficiencies in N- and P-limited tropical forests: a comparison of responses to long-term fertilization. Ecosystems 4(7):646–657
IPCC (2013) Climate change 2013: the physical science basis. In: Stocker TF, Qin D, Plattner G-K, Tignor M, Allen SK, Boschung J, Nauels A, Xia Y, Bex V, Midgley PM (eds) Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change. Cambridge University Press, Cambridge, UK. http://www.climatechange2013.org/report/full-report/ipcc
Jobbagy EG, Jackson RB (2000) The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecol Appl 10(2):423–436
Liu XF, Lin TC, Yang ZJ, Vadeboncoeur MA, Lin CF, Xiong DC, Lin WS, Chen GS, Xie JS, Li YQ, Yang YS (2017) Increased litter in subtropical forests boosts soil respiration in natural forests but not plantations of Castanopsis carlesii. Plant Soil 418(1–2):141–151
Luo YQ, Ahlstrom A, Allison SD, Batjes NH, Brovkin V, Carvalhais N, Chappell A, Ciais P, Davidson EA, Finzi AC, Georgiou K, Guenet B, Hararuk O, Harden JW, He YJ, Hopkins F, Jiang LF, Koven C, Jackson RB, Jones CD, Lara MJ, Liang JY, McGuire AD, Parton W, Peng CH, Randerson JT, Salazar A, Sierra CA, Smith MJ, Tian HQ, Todd-Brown KEO, Torn M, van Groenigen KJ, Wang YP, West TO, Wei YX, Wieder WR, Xia JY, Xu X, Xu XF, Zhou T (2016) Toward more realistic projections of soil carbon dynamics by earth system models. Glob Biogeochem Cycles 30(1):40–56
Maycock CR, Congdon RA (2000) Fine root biomass and soil N and P in north Queensland rain forests. Biotropica 32(1):185–190
McGroddy M, Silver WL (2000) Variations in belowground carbon storage and soil CO2 flux rates along a wet tropical climate gradient. Biotropica 32:614–624
Mehlich A (1984) Mehlich 3 soil test extractant: a modification of Mehlich 2 extractant. Commun Soil Sci Plant Anal 15:1409–1416
Metcalfe DB, Meir P, Aragao L, da Costa ACL, Braga AP, Goncalves PHL, Silva JD, de Almeida SS, Dawson LA, Malhi Y, Williams M (2008) The effects of water availability on root growth and morphology in an Amazon rainforest. Plant Soil 311(1–2):189–199
Nottingham AT, Turner BL, Chamberlain PM, Stott AW, Tanner EVJ (2012) Priming and microbial nutrient limitation in lowland tropical forest soils of contrasting fertility. Biogeochemistry 111(1–3):219–237
Oades JM (1988) The retention of organic matter in soils. Biogeochemistry 5(1):35–70
Phillips RP, Finzi AC, Bernhardt ES (2011) Enhanced root exudation induces microbial feedbacks to N cycling in a pine forest under long-term CO2 fumigation. Ecol Lett 14(2):187–194
Post WM, Emanuel WR, Zinke PJ, Stangenberger AG (1982) Soil carbon pools and world life zones. Nature 298(5870):156–159
Powers JS, Schlesinger WH (2002) Relationships among soil carbon distributions and biophysical factors at nested spatial scales in rain forests of northeastern Costa Rica. Geoderma 109(3–4):165–190
Pries CEH, Bird JA, Castanha C, Hatton PJ, Torn MS (2017) Long term decomposition: the influence of litter type and soil horizon on retention of plant carbon and nitrogen in soils. Biogeochemistry 134(1–2):5–16
Pyke CR, Condit R, Aguilar S, Lao S (2001) Floristic composition across a climatic gradient in a neotropical lowland forest. J Veg Sci 12(4):553–566
Rasse DP, Rumpel C, Dignac MF (2005) Is soil carbon mostly root carbon? Mechanisms for a specific stabilisation. Plant Soil 269(1–2):341–356
Russell AE, Hall SJ, Raich JW (2017) Tropical tree species traits drive soil cation dynamics via effects on pH: a proposed conceptual framework. Ecol Monogr 87(4):685–701
Sayer EJ, Heard MS, Grant HK, Marthews TR, Tanner EVJ (2011) Soil carbon release enhanced by increased tropical forest litterfall. Nat Clim Chang 1(6):304–307
Schuur EAG (2001) The effect of water on decomposition dynamics in mesic to wet Hawaiian montane forests. Ecosystems 4(3):259–273
Schwarz G (1978) Estimating dimension of a model. Ann Stat 6(2):461–464
Silver WL, Miya RK (2001) Global patterns in root decomposition: comparisons of climate and litter quality effects. Oecologia 129(3):407–419
Sollins P, Homann P, Caldwell BA (1996) Stabilization and destabilization of soil organic matter: mechanisms and controls. Geoderma 74(1–2):65–105
Staff SS (1999) Soil taxonomy: a basic system of soil classification for making and interpreting soil surveys lincoln, NE
Stewart R, Stewart J, Woodring W (1980) Geologic map of the Panama Canal and vicinity, Republic of Panama. Map I-1232. United States Geological Survey, Boulder
Tanner EVJ, Sheldrake M, Turner BL (2016) Changes in soil carbon and nutrients following six years of litter removal and addition in a tropical semi-evergreen rain forest. Biogeosciences 13:6183–6190
Telles EDC, de Camargo PB, Martinelli LA, Trumbore SE, da Costa ES, Santos J, Higuchi N, Oliveira RC (2003) Influence of soil texture on carbon dynamics and storage potential in tropical forest soils of Amazonia. Glob Biogeochem Cycles 17(2):12
Todd-Brown KEO, Randerson JT, Post WM, Hoffman FM, Tarnocai C, Schuur EAG, Allison SD (2013) Causes of variation in soil carbon simulations from CMIP5 Earth system models and comparison with observations. Biogeosciences 10(3):1717–1736
Torn MS, Trumbore SE, Chadwick OA, Vitousek PM, Hendricks DM (1997) Mineral control of soil organic carbon storage and turnover. Nature 389(6647):170–173
Turner BL, Engelbrecht BMJ (2011) Soil organic phosphorus in lowland tropical rain forests. Biogeochemistry 103(1–3):297–315
Turner BL, Romero TE (2009) Short-term changes in extractable inorganic nutrients during storage of tropical rain forest soils. Soil Sci Soc Am J 73(6):1972–1979
Turner BL, Yavitt JB, Harms KE, Garcia MN, Wright SJ (2015) Seasonal changes in soil organic matter after a decade of nutrient addition in a lowland tropical forest. Biogeochemistry 123(1–2):221–235
Vitousek PM, Sanford RL (1986) Nutrient cycling in moist tropical forests. Annu Rev Ecol Syst 17:137–167
Walker T, Adams A (1958) Studies on soil organic matter: I. Influence of phosphorus content of parent materials on accumulations of carbon, nitrogen, sulfur, and organic phosphorus in grassland soils. Soil Sci 85:307–318
Waring BG, Powers JS (2017) Overlooking what is underground: root:shoot ratios and coarse root allometric equations for tropical forests. For Ecol Manage 385:10–15
Windsor D, Rand A, Rand W (1990) Caracteristicas de la Precipitacion de la Isla de Barro Colorado
Woodring W (1958) Geology of Barro Colorado Island, Collection 135:1–39. In: Smithsonian (ed) Panama
Wright SJ, Yavitt JB, Wurzburger N, Turner BL, Tanner EVJ, Sayer EJ, Santiago LS, Kaspari M, Hedin LO, Harms KE, Garcia MN, Corre MD (2011) Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology 92(8):1616–1625
Wurzburger N, Wright SJ (2015) Fine-root responses to fertilization reveal multiple nutrient limitation in a lowland tropical forest. Ecology 96(8):2137–2146
Yavitt JB, Wright SJ (2001) Drought and irrigation effects on fine root dynamics in a tropical moist forest, Panama. Biotropica 33(3):421–434
Yavitt JB, Harms KE, Garcia MN, Mirabello MJ, Wright SJ (2011) Soil fertility and fine root dynamics in response to 4 years of nutrient (N, P, K) fertilization in a lowland tropical moist forest, Panama. Austral Ecol 36(4):433–445
Acknowledgements
Funding was provided by NSF GSS Grant #BCS-1437591 and DOE Grant DE-SC0015898 to D. F. Cusack, and NERC Grant NE/J011169/1 to O. T. Lewis. We thank Julio Rodriguez, Didimo Urena, David Brassfield, Evert Carlos Green, and Christian Harris for field support, and Dayana Agudo, Aleksandra Bielnicka, Dianne de la Cruz, Tania Romero, Irene Torres, Tyler Schappe and Stanley L. Walet for laboratory support. Assistance with map and graphics was provided by Matt Zebrowski, cartographer, at UCLA. Data supporting the conclusions is available in tables, figures, and SI.
Author information
Authors and Affiliations
Corresponding author
Additional information
Responsible Editor: Edward Brzostek.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Cusack, D.F., Markesteijn, L., Condit, R. et al. Soil carbon stocks across tropical forests of Panama regulated by base cation effects on fine roots. Biogeochemistry 137, 253–266 (2018). https://doi.org/10.1007/s10533-017-0416-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-017-0416-8