
Abstract
Recent measurements have demonstrated unprecedented increase in atmospheric deposition of nutrients in many parts of India. To determine whether atmospheric nutrient inputs would increase phytoplankton growth and catchment dissolved organic carbon (DOC) flushing to constrain benthic algae, we analyzed NO3 − and PO −34 in atmospheric deposits; nutrients and DOC in runoff and lake water and standing crop biomass of phytoplankton and periphyton at Jaisamand Lake of Rajasthan, India. Atmospheric deposition of NO3 − (7.18–29.95 kg ha−1 year−1) and PO −34 (0.56–2.15 kg ha−1 year−1) showed a consistently rising trend across the year. Microbial biomass and activity in catchment increased in response to atmospheric deposition. Lake DOC and nutrients showed strong coherence with their terrestrial and atmospheric fluxes. Phytoplankton development showed significant linearity with atmospheric input of nutrients. Air-driven input appeared to have compensated the nutrient constraints to phytoplankton during drought. The N:P stoichiometry of deposition and that of lake water indicated that, although there was a seasonal switchover to N- or P-limitation, phytoplankton were mainly co-limited by N and P due probably to the synergistic effects of combined N + P enrichment in the pelagic zone of the lake. Periphyton standing crop showed inverse relationship with phytoplankton and lake DOC. The study indicated that enhanced phytoplankton development and terrestrial DOC flushing in response to atmospheric nutrient input attenuated light penetration to constrain algal periphyton. We suggests that data on these issues may be considered in developing aquatic ecosystem models to establish future links between changing air–water–land interactions and associated shifts in lake ecosystem functioning for more accurately predicting climate change drivers and designing integrated lake basin management strategies.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
During recent years, anthropogenic activities have dramatically increased the atmospheric deposition of pollutant aerosols in many parts of India. Much research efforts in our country, on atmospheric deposition-linked changes in ecosystem functioning have been devoted towards terrestrial ecosystems (Singh and Tripathi 2000). Surface water bodies have received less attention on these issues. Air-borne nutrients can be transported long distances and affect aquatic environments far from their sources (Bergstrom et al. 2008). This may lead to long-term changes in ecosystem functioning and may constrain restoration of surface water resources.
The catchment characteristics together with human modifications and hydrology regulate the water quality of lakes and rivers. Forest clearing along with agricultural and industrial activities in the catchment could substantially modify the amount of carbon and nutrients being flushed to receiving waters (Tilman et al. 2001). Deposition of air-borne pollutant elements modifies watershed related properties which may accelerate nutrient and carbon flushing to receiving waters (Monteith et al. 2007). Studies have indicated atmospheric-N deposition as high as 20–60 kg N ha−1 year−1 in many ecosystems in Europe (Bobbink and Lamers 2002) and from 12 to 35 kg N ha−1 year−1 in many parts of India (Pandey and Pandey 2009). Climatic extremes such as drought could change vegetation cover and pedo-atmospheric interfacial interactions in the lake catchments and consequently the runoff-linked changes in the chemistry of receiving waters (Krivtsov et al. 2001). Reduced precipitation in the drainage basin reduces stream flow, terrestrial carbon pool (productivity) and dissolved organic carbon (DOC) loading to lakes and streams. Drought restricts growth of natural vegetation especially those of annuals. As a result, nutrient extraction and flushing towards receiving water is reduced. Eimers et al. (2008) observed that trends in DOC concentration were well associated with terrestrial carbon pools. The C:N and C:P ratios of allochthonous organic matter rich in tree foliage are more than threefold higher than autochthonous particulates generally dominated by phytoplankton (Elser et al. 2000). Atmospheric deposition of N and P may reduce C:N and C:P ratios of terrestrial organic matter enhancing microbial colonization (Pandey and Pandey 2001). Furthermore, air-borne nutrients increase productivity (C-pool) of terrestrial (Singh and Tripathi 2000) and aquatic ecosystems (Bergstrom et al. 2005; Bergstrom and Jansson 2006), as well as microbial biomass and activity in catchment soils (Pandey 2011). DOC of terrestrial origin is extensively regulated by microbial metabolism in soils, streams and at land–water interfaces as it flows towards recipient lakes (Jansson et al. 2000). These processes together may significantly enhance the magnitude of allo- and autochthonous-C inputs in receiving waters. Studies on run-off waters from different catchments have indicated that the regional variations in climate, soil and vegetation significantly affect the chemistry and biology of aquatic ecosystems (Zade et al. 2005).
Phytoplankton productivity in tropical lakes, where light and temperature regimes are generally optimal, is affected mainly by nutrients (Yang et al. 2008). On the other hand, the growth of algal periphyton, the submerged micro-floral community living attached to the substrate, can be primarily light limited (Carey et al. 2007). Thus, in oligotrophic lakes, elevated levels of DOC can effectively constrain light penetration and growth of benthic primary producers (Karlsson et al. 2009). Similarly, in eutrophic lakes, high phytoplankton chlorophyll associated with high nutrient input can reduce light penetration and consequently the periphyton production (Vadeboncoeur et al. 2003). In particular, colored humic substances of terrestrial origin more effectively attenuate water column light penetration (Karlsson et al. 2009). Algal periphyton supply oxygen to benthic organisms, regulate nutrient cycling and contribute up to 39 % of biomass of primary producers in oligotrophic lakes (Wetzel 2001). Further, benthic algae help reducing nutrient release from bottom sediments (Christensen et al. 1997) and recovering the water body from eutrophication (Luijn et al. 1995). However, despite high contribution to aquatic ecosystem functioning, the periphyton community has received little attention for tropical lakes of India.
DOC enrichment in oligotrophic lakes is a derived character, which may be accelerated through atmospheric deposition-linked catchment DOC flushing and pulsed phytoplankton growth as has been illustrated for Lake Baghdara (Pandey 2011). Lake Jaisamand, considered in the present study, is a large freshwater reservoir with heterogeneous habitat conditions to support rich algal diversity including nutritionally rich benthic Rhodophycean Batrachospermum moniliformae, an important algal food supplement of Bhil tribe. From the last 15 years, Udaipur region of India has been witnessing high frequency of drought and consequently, increased pressure on surface water resources. Further, this period has also witnessed an unprecedented urban-industrial growth and as a result, the atmospheric deposition increased rapidly from 1997 onward. However, there is a general dearth of studies explicitly analyzing the relationships in the dynamics of phytoplankton, periphyton and DOC and their linkages with changing climatic extremes and atmospheric deposition in tropical lakes of India. We hypothesized that nutrient inputs through atmospheric deposition would increase phytoplankton pulse and catchment DOC flushing through runoff to constrain growth of algal periphyton in Lake Jaisamand. To address this hypothesis, we tested whether patterns of microbial biomass in catchment soils, DOC flushing to the lake, and phytoplankton growth varied over time in relation to atmospheric-N and -P deposition or drought. To examine possible effects of catchment processes on partitioning of productivity within the lake, we also tested whether productivity by benthic algae was negatively related to DOC and phytoplankton productivity.
Materials and methods
Study area
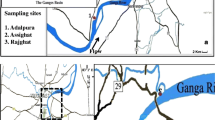
The data presented here are the results of seven consecutive years of study (2003–2009) conducted at Lake Jaisamand (catchment area: 180,974 ha; surface area 8,700 ha; maximum depth: 36 m) situated about 52 km SE of Udaipur city on Udaipur–Banswara road (240 16′N latitude; 73°54′E longitude; 560 m above msl) in the Udaipur District of Rajasthan, India. This largest artificial lake of Asia was constructed during 1711–1730 AD primarily for conservation of wildlife. When first constructed, it was the largest artificial lake of the world. The lake has three islands measuring about 160,000 m2 each. It receives water from six small rivers, where Gomati River regulates the main inflow. Climate of the region is tropical with distinct seasonality. The year is divisible into three distinct seasons, a hot and dry summer (April–June), a warm and wet rainy season (July–September) and, a cool and dry winter (November–February). March and October represent transition months. During the study period, mean annual rainfall varied between 360 and 815 mm, relative humidity ranged between 17–81 % (summer) and 44–96 % (rainy season). During summer, day temperature varied between 25.3 and 46.3 °C. During winter, night temperature sometimes drops below 4 °C. In the last two decades, for more than half of the period, this region was affected by drought, which was almost continuous between 1999 and 2004 (Lobo 2008).
Catchment characteristics that were hypothesized to influence links between terrestrial flushing and water quality are shown in Table 1. A detailed description of catchment characteristics is available in Lobo (2008). Of the four catchment landscape categories considered in this study, two (tussocked rocky terrains, RT and woodland, WL) were in a relatively natural state and the other two (agricultural land, AL and pavement, PT) were invariably human disturbed. Samples were collected for all the seven consecutive years. However, for data comparisons, in cases where inter-annual variations were less marked, time series were divided into an initial study year (2003) and a final study year (2009). Climatic conditions can be justified to be uniform since all the catchment landscapes are situated in a uniform geographical location and it is unlikely that there would be large differences in temperature–humidity variables. Soil of the region is sandy to clay loam. At some places, skeletal rocky and hilly soils are also found. Catchments differed in terms of percentage of fine soils (silt and clay), water soluble organic carbon, sedentary soil on rocky terrains and percentage of artificially fertilized areas. The catchment land use comprises of 16 % forest cover, 6 % irrigated land, 16 % un-irrigated land, 25 % culturable wasteland and 37 % not available for cultivation (Lobo 2008).
Measurements
The experimental design consisted of three tiers of study that include, measurements of atmospheric deposition, chemistry of catchment runoff and lake and stream water and influence on standing crop biomass dynamics of phytoplankton and periphyton.
Atmospheric deposition
Atmospheric deposition of pollutant aerosols were measured using bulk samplers made up of a 5 l high density polyethylene bottle connected to a Teflon funnel and devised with PVC needles on top to avoid bird nesting. Collectors were maintained at a height of 2 m to avoid collection of re-suspended particulates. Samples were collected monthly and at the end of the sampling period the funnels were rinsed with double distilled water to collect particles deposited on funnel walls. As soon as the samples were brought to the laboratory, a 50 ml of sub sample of atmospheric deposition collected in the bottle and a 50 ml of subsample of the rinsing water were filtered through pre-combusted Whatman glass fiber filter (0.7 μm) and analyzed for nitrate and phosphate spectrophotometrically. Samples were screened for contamination using high soluble reactive-P (>0.2 mg) as an indicator (Lohse et al. 2008).
Water chemistry
For analyses of DOC, dissolved inorganic nitrogen (DIN), dissolved organic nitrogen (DON) and dissolved reactive phosphorus (DRP), the runoff waters were collected in pre-sterilized polythene bottles during rain events for the entire study period along the topographic gradients from different catchment landscapes. The topographic gradients representing catchment sampling area ranged from a gentle slope (3.5 %) of agricultural catchment to moderately high slope (8.6 %) of tussocked rocky terrains. Composite water samples were collected monthly at 15–20 m reach in the lake during the study period. The distance between replicate sampling (n = 3) was about 50 m. Water samples were also collected from the stream inlet on the same occasions as from the lake. Water was collected from each site, directly below the surface (6–10 inch depth), in acid-rinsed 5 l plastic containers, for analysis of DOC, DIN, DON and DRP. DOC was quantified using a KMNO4 digestion procedure (Michel 1984). A known volume of water sample was mixed with acidified N/80 potassium permanganate and incubated at 37° C. Organic carbon was estimated by titrating to quantify oxygen after 4 h of incubation (American Public Health Association 1998). Nitrate–N was quantified using a brucine sulphuric acid method (Voghe 1971) and ammonia–N was quantified using Nessler’s reagent method (Maiti 2001). Total dissolved nitrogen (TDN) was estimated following high temperature persulphate digestion. DON was computed as TDN minus NO3–N and NH4–N (Perakis and Hedin 2002). Orthophosphate (DRP) was quantified using Olsen′s ammonium molybdate method (Mackereth 1963). Depth of light penetration in the lake was measured using Secchi disk at all sites at each sampling date during the study.
Microbial biomass and activity
For measurement of microbial biomass (Cmic) and activity, surface soil samples (0–10 cm depth) were collected at monthly interval during the study period from four randomly selected blocks (plot size, 50 × 50 m) in each catchment landscape category. To ensure possible uniformity in soil moisture regime, sampling plots were selected in riparian zone of the respective catchment. Triplicate samples (n = 3) collected randomly from each plot were pooled and considered as one composite sample for further analysis. Cmic in catchment soil was determined using chloroform fumigation extraction procedure (Tate et al. 1998). An extraction efficiency coefficient of 0.38 was used to convert soluble-C into microbial biomass-C. For substrate induced respiration (SIR, a measure of active microbial biomass), 20 g of soil sample (moisture content, 55 %) was taken in a 250 ml container and amended with 120 mg of glucose at the beginning of incubation (Anderson and Domsch 1978). The container with soil sample and 25 ml of 0.5 N NaOH perched in it was made air tight. CO2–C flux was determined by titrating excess of NaOH with 0.5 N HCl. Metabolic quotient (qCO2) was expressed as CO2–C flux per unit of microbial biomass.
Algal biomass
For phytoplankton biomass, measured in terms of chlorophyll a, water samples were collected in acid-rinsed 5 l plastic containers from each replication point (n = 3), directly below the surface (6–10 inch depth). From this sample, subsamples taken for phytoplankton were preserved in Lugol’s iodine and concentrated by centrifugation at 3,500 rpm (American Public Health Association 1998). Chlorophyll a, extracted from this concentrate using acetone, was measured spectrophotometrically. Phytoplankton standing crop biomass was expressed in terms of chlorophyll a (chlorophyll a constitutes approximately 1.5 % of algal dry organic matter, American Public Health Association 1998). For periphyton, 35 mm plastic slides were laid in triplicate over the rock surface fixed in closed wire cage (basket sampler) of 15 × 15 × 15 cm size at 3 m depth for 1 month. Each replicate represents a composite sample of three independent slides placed at each collection point at a distance of 2 m. The material within the slide area (scrub area = 2.3 cm × 3.5 cm = 8 cm2) was pulled by vacuum onto a pre-ashed (free of organic matter) glass fiber filter (Whatman GF/A) for wet and dry weights. Enough care was taken to ensure complete recovery of algal assemblages attached to the surface. Chlorophyll a biomass in phytoplankton and periphyton was determined in fresh material following acetone extraction procedure (Maiti 2001). Periphyton chlorophyll a values were expressed as mg m−2 by relating to exposed surface area (8 cm2). Phycoerythrin pigment in freshwater red alga, B. moniliformae (Florideophyceae) was extracted in 0.1 mol phosphate buffer (pH 6.8) and quantified following spectrophotometric technique of Beer and Eshel (1985).
Statistical analysis
Significant effects of site (catchment landscape) and time series (year) were determined using analysis of variance (ANOVA). Sampling locations were sampled repeatedly over time and all repeated measurements, except a few that were excluded for not following a normal distribution pattern, were considered in ANOVA model. Significant differences in stream flow data also were tested using ANOVA. Where necessary, values were log transformed. Interaction terms were always included in the analysis. To justify variances, means were supported with standard errors. Coefficients of variation (CV) across time were computed for expressing data variability. Correlation coefficient (R) and regression analyses were used to test relationships between atmospheric deposition variables (N and P) and microbial biomass, lake DIN, DRP, DOC and phytoplankton (Chl a). For light related relationships, the variables tested against light penetration (Secchi disk transparency) were lake DOC and periphyton. Relationship between periphyton and phytoplankton was also tested since the latter can potentially influence light penetration in lakes. Confidence limits above 95 (p < 0.05) were considered as significant. SPSS package was used for statistical analysis.
Results
Atmospheric deposition of nutrient ions increased consistently over time (Fig. 1a, b). The N:P stoichiometry of atmospheric deposition (15:1) did not change over time from 2003 to 2009 but there was over 1.5-fold increase in N and P deposition in 2009 compared to 2003. At RT, during 2009, more than 80 % of atmospheric deposition samples contained PO −34 above 1.75 kg ha−1 year−1 and NO3 − above 26 kg ha−1 year−1. Similar were the trends for WL, AL and PT (Fig. 1a, b). Atmospheric deposition did not differ significantly between sites (Table 2).

Year-wise trends in atmospheric NO3 − (a), atmospheric PO4 3− (b), microbial biomass (c), lake dissolved organic carbon (DOC, d) and runoff fluxes of dissolved inorganic nitrogen (DIN, e), dissolved organic nitrogen (DON, f), dissolved reactive phosphorus (DRP, g) and dissolved organic carbon (DOC, h) at four sites of Lake Jaisamand. Values are mean (n = 12) ± 1SE. RT tussocked rocky terrains, WL woodland catchment, AL agricultural catchment, PT pavement
Microbial biomass (Cmic) in the catchment increased consistently over time from 2003 to 2009 (Fig. 1c). Coefficient of variations in Cmic and activity increased marginally over time. Cmic and atmospheric-N (R = 0.41, p < 0.02, n = 28) and Cmic and atmospheric-P (R = 0.44, p < 0.02, n = 28) were correlated; when catchment influences were removed, the relationships were stronger (R = 0.98, p < 0.0001, n = 7 for Cmic–N and R = 0.97, p < 0.0001, n = 7 for Cmic–P). Microbial biomass and activity both were highest in WL and lowest in PT. During 2009 summer, relative to 2003, Cmic at WL increased by 46 %. Similar was the trend for microbial activity, measured as SIR and metabolic quotient (qCO2) where the corresponding increases in WL were 40 and 44 % respectively. On seasonal scale, Cmic was highest (412–1050 μg g−1) in summer and lowest (280–784 μg g−1) in rainy season. On the other hand, microbial activity remained highest in rainy season (Table 3).
Lake- and runoff DOC and nutrients increased over time (Fig. 1d–h) and the increases over time were highest for WL. The lake received maximum runoff DOC from AL (189.22–212.10 kg ha−1 year−1) followed by WL (162.40–208.75 kg ha−1 year−1), RT (120.45–154.82 kg ha−1 year−1) and PT (54.08–82.67 kg ha−1 year−1). Similar was the trend for DIN (1.86–13.40 kg ha−1 year−1), DON (0.63–6.76 kg ha−1 year−1) and DRP (0.39–1.94 kg ha−1 year−1) flushed to lake through runoff (Fig. 1e–g). Spatial and temporal differences in lake DOC as well as in runoff DOC and nutrients were significant (p < 0.001, Table 2). As stream inflow, Gomati River adds 3.20–6.25 mg l−1 DOC; 0.17–0.67 mg l−1 DIN; 0.06–0.18 mg l−1 DON and 0.07–0.29 mg l−1 DRP (Table 4). River inflow of nutrients was highest during rainy season.
Concentrations of DIN, DRP and phytoplankton Chl a in the lake showed seasonal trends similar to that of atmospheric deposition (Fig. 2a–e). With respect to the year, similar to DOC, lake DIN and DRP increased over time from 2003 to 2009 (Fig. 3). During 2009, relative to 2003, lake water contained over threefold higher nutrients and over fourfold higher DOC. In an annual cycle, lake DOC showed two peaks opposite to Secchi disk transparency (Fig. 4a, b). The dip in Secchi disk transparency (measured as Secchi depth) as well as in periphyton biomass did appear after terrestrially derived DOC input in July (Fig. 4b, d). Periphyton Chl a showed two peaks almost opposite to that of phytoplankton (Fig. 4c, d). Similar was the case of phycoerythrin pigment of a Rhodophycean benthic alga B. moniliformae. Atmospheric-N showed significant positive relationship with lake DIN (R = 0.93, p < 0.0001, n = 28), phytoplankton Chl a (R = 0.92, p < 0.0001, n = 28) and lake DOC (R = 0.92, p < 0.0001, n = 28). Similarly, atmospheric-P showed significant positive relationship with lake DRP (R = 0.97, p < 0.0001, n = 28), phytoplankton Chl a (R = 0.96, p < 0.0001, n = 28) and lake DOC (R = 0.97, p < 0.0001, n = 28; Fig. 5a–f).

Seasonal patterns in atmospheric NO3 − (a), atmospheric PO4 3− (b), lake DIN (c), DRP (d) and phytoplankton Chl a (e) in Lake Jaisamand. Phytoplankton Chl a is expressed as biomass per unit water volume. Values are mean (n = 7) ± 1SE

Year-wise trend in the concentrations of dissolved inorganic nitrogen (DIN), dissolved organic nitrogen (DON) and dissolved reactive phosphorus (DRP) in Lake Jaisamand. Values are mean (n = 12) ± 1SE

Month-wise trends in lake DOC (a), Secchi disk transparency (b), phytoplankton Chl a (c) and periphyton Chl a (d) in Lake Jaisamand. Seechi disk transparency presents depth (m) of light penetration. Phytoplankton Chl a is expressed as biomass per unit water volume and periphyton Chl a is expressed as biomass per unit surface area. Data (mean (n = 7) ± 1SE) comparisons are made between initial (2003) and final (2009) study year

Significant relationships between a atmospheric-N and lake DIN (R = 0.93, p < 0.0001, n = 28), b atmospheric-N and phytoplankton Chl a (R = 0.92, p < 0.0001, n = 28), c atmospheric-N and Lake DOC (R = 0.92, p < 0.0001, n = 28), and d between atmospheric P and lake DRP (R = 0.97, p < 0.0001, n = 28), e atmospheric P and phytoplankton Chl a (R = 0.96, p < 0.0001, n = 28) and atmospheric P and lake DOC (R = 0.97, p < 0.0001, n = 28)
Variations in phytoplankton and periphyton growth over time from 2003 to 2009 showed almost opposite trends (Fig. 6a, b). Periphyton Chl a biomass declined over time by approximately 77 %. Conversely, phytoplankton Chl a biomass increased over time (Fig. 6a) and in 2009, it increased by about three orders of magnitude higher than the values recorded in 2003. Secchi disk transparency declined over time and showed significant negative relationship with lake DOC (R = 0.97, p < 0.0001, n = 24). Periphyton Chl a biomass that showed significant negative relationship with lake DOC (R = 0.76, p < 0.0001, n = 24) and phytoplankton Chl a (R = 0.69, p < 0.0002, n = 24), showed significant positive relationship with Secchi disk transparency (R = 0.84, p < 0.0001, n = 24; Fig. 7).

Annual trends in phytoplankton Chl a (a) and periphyton Chl a (b) in Lake Jaisamand. Phytoplankton Chl a is expressed as biomass per unit water volume and periphyton Chl a is expressed as biomass per unit surface area. Values are mean (n = 12) ± 1SE

Significant relationships between a Lake DOC and Secchi depth (R = 0.97, p < 0.0001, n = 24) and b lake DOC and periphyton Chl a (R = 0.76, p < 0.0001, n = 24), c Secchi depth and periphyton Chl a (R = 0.84, p < 0.0001, n = 24) and d between phytoplankton Chl a and periphyton Chl a (R = 0.69, p = 0.0002, n = 24)
Discussion
Nutrient enrichment
Chemistry of lake water depends on constituents added through surface discharge as well as through atmospheric deposition and data on these issues is important in developing integrated lake basin management strategies. Our results indicate that the concentrations of nutrients in lake increased in response to atmospheric deposition. From 2003 to 2009, atmospheric inputs of nutrients increased by 1.5-fold but the concentration of nutrients in lake increased by over threefold. Substantially high increase in lake nutrient concentration possibly indicates the effect of direct atmospheric deposition onto the lake water together with indirect addition through runoff flushing (Bergstrom et al. 2005). Correlation analyses showed that the nutrients in lake as well as in runoff waters responded positively to atmospheric inputs. Hydrological release of nutrients from the catchment depends also on atmospheric deposition-linked changes in uptake or leaching of nutrients by plants (McEachern et al. 2000), changes in soil chemistry and, microbial consumption or release of nutrients (Pandey and Pandey 2009). Catchment-wise differences in runoff DIN:DRP ratios indicate one possible result of such effects. The DIN:DRP ratios in runoff added to the lake were generally high for vegetal catchment due probably to faster uptake of air-driven P than N by catchment vegetation (McEachern et al. 2000). Additionally, high runoff flushing of DIN and DRP from agricultural catchment shows hydrological and anthropogenic controls on N- and P-export (Chen and Hong 2011).
A close correspondence between stream and lake concentrations of nutrients, both within and across seasons, is important evidence that stream water inputs also are driving changes over time. Changes in nutrient concentrations of our study lake were large and we attempted to explore possible drivers of such changes. Our data indicate a highly dynamic and tightly coupled linkage between atmospheric input and stream water input of nutrients. Stream water transport during monsoon was strongly tied with atmospheric fluxes indicating the role of principal state factors (geologic parent materials, topography and biota) coupled with atmospheric inputs. Dry season changes in lake water chemistry were triggered principally by atmospheric inputs indicating the influence of changing atmosphere–water coupling.
DOC enrichment
Our data indicated a rising trend in DOC concentration over time from 2003 to 2009. The increases in DOC concentrations in lakes of Europe and North America have been generally linked to enhanced pelagic production in response to nutrient enrichment and/or enhanced terrestrial flushing driven by climate and hydrological changes (Evans et al. 2006; Porcal et al. 2009). Among the climatic variables, temperature is considered as the principal driver of DOC concentration in boreal and subarctic lakes (Weyhenmeyer and Karlsson 2009). However, temperature did not appear a major driver of increasing DOC concentration in the study lake. A close correspondence between stream, runoff and lake DOC indicates that hydrological flushing is an important driver. On the other hand, phytoplankton pulse and associated shift in DOC, especially during dry seasons, is important evidence that pelagic production is driving changes in lake DOC. It seemed that overall trend in study lake DOC was the result of tightly coupled effect of hydrologic flushing and pelagic production.
Spatial variation in runoff DOC appeared to be due probably to differences in land use, as the values were lowest for PT devoid of vegetation and highest for AL witnessing agricultural activity. Although we have shown remarkably strong hydrologic forcing of rainy season DOC, year-wise trend could be linked to atmospheric nutrients in a major way. DOC of terrestrial origin is sensitive to climate and watershed properties such as litter quality and microbial activity in soil, streams and land–water interface (Tranvik and Jansson 2002; Porcal et al. 2009). In the present study, it seemed that atmospheric nutrient input did enhance the phytoplankton production and runoff DOC and, by implication, DOC in lake water. Studies have indicated that atmospheric deposition of nutrients enhances phytoplankton development (Bergstrom et al. 2008) and lake DOC (Pandey 2011). Significant correlation between runoff DOC and atmospheric-N input suggests that air–watershed interaction could influence DOC loading to Jaisamand. Changes in soil organic matter (SOM) solubility and the rate of microbial production or consumption of DOC from SOM in uppermost carbon rich soil horizon, where the majority of DOC production occurs (Kalbitz et al. 2000), are important drivers of DOC in receiving waters. In a recent study, a substantial increase in microbial activity at land–water interface of a woodland catchment and consequent increase in DOC release in response to nutrient deposition was observed (Pandey 2011). Organic matter rich in tree foliage (as in case of woodland catchment) with high concentration of phenolics and high C:N ratio (Elser et al. 2000) is less prone to microbial attack. Significant temporal coherence between microbial biomass (Cmic), atmospheric nutrient input and runoff DOC indicate that air-driven nutrients could induce microbial colonization by improving substrate quality (Pandey and Pandey 2009). Nitrogen deposition has been shown to decrease C:N ratio, increase water soluble organic-C (Wedin and Tilman 1996) and microbial activity (Pandey and Pandey 2009). Similarly, long-term phosphorus fertilization enhances soluble P and alters C:N ratio in soil (Schipper et al. 2011). Atmospheric nutrient-linked increase in Cmic merits attention in context of terrestrial carbon cycling and associated shift in DOC export to surface waters.
Nutrient limitation
To assess the possible effects of changing nutrient balance on nutrient limitation of phytoplankton, we examined N:P stoichiometry of atmospheric deposition and lake water and the relationships between nutrients and phytoplankton chlorophyll a in the study lake over time. Unlike the lakes of temperate and polar regions, light and temperature are not among the major limiting factors for phytoplankton in tropical freshwaters (Yang et al. 2008). Further, since we collected samples at 6–10 inch depth, we consider that phytoplankton development was not limited by light availability.
In the present study, the N:P stoichiometry of atmospheric deposition (15:1) did not change but the levels of these nutrients enhanced causing higher phytoplankton biomass in the lake. Despite low DIN:DRP ratios in the runoff from 2006 onward, DIN:DRP ratios in lake water increased during subsequent years suggesting that N-limitation of initial years shifted to P-limitation in accordance of rising input of air-driven N. Bergstrom et al. (2008) showed that the regional differences in nutrient limitation of phytoplankton in unproductive Swedish lakes were related to atmospheric-N input and lake DIN concentrations. Earlier studies have indicated that unproductive lakes in low N deposition areas are naturally N-limited but that a transition to P-limitation could result from alterations in the balance of N and P through increased anthropogenic N inputs (Bergstrom and Jansson 2006; Elser et al. 2009). Further, as also reported by Bergstrom et al. (2005), high DIN:DRP ratios as well as Chl a:DRP ratios in winter indicated that the lake was under P-limitation during winter but shifted to N-limitation during summer. Inter-annual variations in DIN:DRP as well as in Chl a:DRP ratios indicated that although N-limitation was more pronounced than P-limitation, phytoplankton in study lake were mainly co-limited by N and P. The idea of co-limitation is further supported by the positive correlations observed between atmospheric deposition and lake chemistry and phytoplankton productivity. P-limitation might be expected in lakes, and we might expect N deposition to perpetuate this limitation. However, co-limitation of phytoplankton observed in the study lake could be due probably to the synergistic effects of N + P enrichment in the pelagic zone of the lake (Elser et al. 2009). A seasonal switchover to N- or P-limitation could be due to altered nutrient balance under the influence of hydrological flushing and atmospheric deposition.
Light limitation
Catchment export of terrestrial organic matter induces pronounced effect on lake ecosystems by changing the light climate and productivity (Karlsson et al. 2009). Here, we hypothesized that atmospheric deposition of nutrients that increase phytoplankton production and catchment flushing of organic matter (Bergstrom et al. 2005; Pandey and Pandey 2009), will have pronounced effect on benthic algal production through increased light attenuation in a large freshwater lake of India. Our results show that the atmospheric deposition of nutrients modifies the interactions of habitat boundaries (decreases light availability to benthic primary producers through increased phytoplankton-shading effect) and cross-ecosystem boundaries (increases terrestrial DOC flushing to reduce benthic production by attenuating light penetration) which may, in turn, modify the lake ecosystem functioning in long-run.
Periphyton biomass appeared to be controlled by something other than nutrients. The decrease in periphyton biomass with increasing nutrient input could be related to rising DOC and phytoplankton which constrains light penetration (Carey et al. 2007). Correlative evidence suggests that both, phytoplankton and DOC could reduce benthic algal production through light attenuation. Vadeboncoeur et al. (2003) observed that light attenuation associated with increased phytoplankton production along a gradient of P input in Greenland and Danish lakes caused significant loss of benthic periphyton. Our results indicate, although the effect of DOC and phytoplankton shading did not rule out one from the other, the former could be more crucial than the latter in attenuating light penetration. Both, DOC and phytoplankton biomass increased over time from 2003 to 2009 while light penetration and periphyton biomass showed an opposite trend. Benthic periphyton showed a sharp dip in July and had stronger negative correlation with DOC than with Chl a suggesting that terrestrially derived DOC being very optic dense could reduce light penetration more effectively than Chl a. High terrestrial DOC flushing during rainy season seemed to reduce light penetration and consequently periphyton despite low growth of phytoplankton. Additionally, a rapid turnover of DOC at the time of phytoplankton fall could enhance its overall concentration in lake water (Kirchman et al. 1991). A decrease in phytoplankton production during rainy season could be linked to continue terrestrial flushing and increased turbulence (O’ Reilly 2006). Reduced precipitation, as happens during drought, may reduce allochthonous DOC loading to streams and lakes (Eimers et al. 2008; Sowerby et al. 2010). Although additional variables need to be considered (Worrall et al. 2006), this effect of drought on lake DOC, could partly be offset by enhanced pelagic production (and consequently autochthonous DOC pool) caused by atmospheric nutrient input. As evidenced through seasonal pattern, phytoplankton standing crop, which may primarily be nutrient limited in tropical waters (Yang et al. 2008), showed pulsed growth during the period of high atmospheric deposition.
In the present study, a 7 mg l−1 increase in lake DOC caused light penetration to decline by 34 %. Further increase in DOC showed a consistently increasing light attenuation. These observations are in accordance with those recorded by Bergstrom et al. (2001), where a 7 mg l−1 increase in lake DOC had been shown to correspond to an approximately 37 % decrease in light climate. McEachern et al. (2000) observed that a 9 % rise in DOC led Secchi disk transparency to decline by 46 %. Luijn et al. (1995) have reported that even 15–20 % attenuation in Secchi depth can significantly alter the growth of benthic diatoms. Light limitation of periphyton was indicated also by a benthic red alga B. moniliformae wherein the phycoerythrin pigment reduced by over 75 % during highest concentration of lake DOC. Kaczmarezyk and Sheath (1991) have reported marked synchrony between irradiance and phycoerythrin pigment in Batrachospermum boryanum. It is obvious that increased catchment DOC loading and phytoplankton production in response to atmospheric deposition effectively decreased the benthic primary production by attenuating light penetration in the lake. It is possible that, for the whole lake productivity, the net reduction in benthic production may partly be offset by an enhanced pelagic production caused by higher nutrient levels in the water mass. However, since a substantial part of the resources supporting higher trophic levels are generated in the benthic habitats (Karlsson et al. 2009), a shift in benthic primary production may lead to long-term changes in the lake ecosystem functioning.
Conclusions
Correlative evidence in this 7 year study suggests that rising atmospheric deposition of nutrients is increasing lake nutrient concentrations and phytoplankton productivity. Although there was a seasonal switchover to N or P-limitation, phytoplankton in Study Lake were mainly co-limited by N and P and air-driven inputs appeared to have compensated nutrient constraints to phytoplankton during drought.
We conclude that rising atmospheric deposition of nutrients will likely re-structure the patterns of lake productivity by eliminating periphyton community of freshwater tropical lakes. Since benthic algae help maintaining dissolved oxygen, reducing nutrient release from bottom sediments and supporting higher trophic levels, a shift in benthic primary production may lead to long-term changes in the lake ecosystem functioning. We further conclude that atmospheric deposition of nutrients could be an important driver to elevate DOC concentration in freshwater tropical lakes. Since DOC is an important component of aquatic ecosystems and global carbon cycle, our observations on air-driven nutrients-linked rise in DOC highlight the need for large-scale inter-regional time series data on changes in land–water interactions driven by atmospheric deposition and climate. Data on these issues may help establishing crucial links between air- and watershed properties and long-term lake ecosystem functioning for predicting climate change drivers and designing integrated lake basin management strategies.
References
American Public Health Association (1998) Standard methods for the examination of water and wastewater. APHA, Washington, DC
Anderson JPE, Domsch KH (1978) A physiologically active method for the quantitative measurement of microbial biomass in soils. Soil Biol Biochem 10:215–221
Beer S, Eshel A (1985) Determining phycoerythrin and phycocyanin concentrations in aqueous crude extracts of red algae. Aust J Mar Freshwat Res 36:785–792
Bergstrom AK, Jansson M (2006) Atmospheric nitrogen deposition has caused nitrogen enrichment and eutrophication of lakes in the northern hemisphere. Global Change Biol 12:635–643
Bergstrom AK, Jansson M, Blomqvist P, Drakare S (2001) The influence of water colour and effective light climate on mixotrophic phytoflagellates in three small Swedish dystrophic lakes. Int Assoc Theor Appl Limnol 27:1861–1865
Bergstrom AK, Blomqvist P, Jansson M (2005) Effects of atmospheric nitrogen deposition on nutrient limitation and phytoplankton biomass in unproductive Swedish lakes. Limnol Oceanogr 50:987–993
Bergstrom AK, Jonsson A, Jansson M (2008) Phytoplankton responses to nitrogen and phosphorus enrichment in unproductive Swedish lakes along a gradient of atmospheric nitrogen deposition. Aquat Biol 4:55–64
Bobbink KR, Lamers LPM (2002) Effects of increased nitrogen deposition. In: Bell JNB, Treshow M (eds) Air pollution and plant life. Wiley, New York, pp 202–235
Carey RO, Vellidis G, Lowrance R, Pringle CM (2007) Do nutrient limit algal periphyton in small black-water coastal plain streams. J Am Water Resour Assoc 5:1183–1193
Chen N, Hong H (2011) Nitrogen export by surface runoff from a small agricultural watershed in southeast China: seasonal patterns and primary mechanisms. Biogeochemistry 106:311–321
Christensen KK, Andersen FO, Jensen HS (1997) Comparison of iron, manganese and phosphorus retention in freshwater littoral sediment with growth of Littorella uniflora and benthic microalgae. Biogeochemistry 38:149–171
Eimers MC, Watmough SA, Buttle JM (2008) Long term trends in dissolved organic carbon concentration: a cautionary note. Biogeochemistry 87:71–81
Elser JJ, Fafan WF, Denno RF, Dobberfuhl DR, Folorin A, Huberty A, Interlandi S, Kilham SS, Mecauley E, Schulz KL, Siemann EH, Sterner RW (2000) Nutritional constraints in terrestrial and freshwater foodwebs. Nature 408:578–580
Elser JJ, Andersen T, Baron JS, Bergstrom AK, Jansson M, Kyle M, Nydick KR, Steger L, Hessen DO (2009) Shifts in lake stoichiometry and nutrient limitation driven by atmospheric nitrogen deposition. Science 326:835–837
Evans CD, Chapman PJ, Clark JM, Monteith DT, Cresser MS (2006) Alternative explanations for rising dissolved organic carbon export from organic soils. Global Change Biol 12:2044–2053
Jansson M, Bergstrom AK, Blomqvist P, Drakare S (2000) Organic carbon and phytoplankton/bacterioplankton production relationships in lakes. Ecology 82:3250–3255
Kaczmarezyk D, Sheath RG (1991) The effect of light regime on the photosynthetic apparatus of the freshwater red alga Batrachospermum boryanum. Crypto Algol 12:249–263
Kalbitz K, Solinger S, Park J-H, Michalzik B, Matzner E (2000) Control on the dynamics of dissolved organic matter in soils: a review. Soil Sci 165:277–304
Karlsson J, Bystrom P, Ask J, Ask P, Persson L, Jansson M (2009) Light limitation of nutrient-poor lake ecosystems. Nature 460:506–509
Kirchman DL, Suzuki Y, Garside C, Ducklow HW (1991) High turnover rates of dissolved organic carbon during a spring phytoplankton bloom. Nature 352:612–614
Krivtsov V, Siegee D, Bellinger E (2001) A one year study of the Rostherne Mere ecosystem: seasonal dynamics of water chemistry, plankton, internal nutrient release and implications for long term trophic status and overall functioning of the lake. Hydrol Process 15:10–17
Lobo V (2008) Characterization of Jaisamand Lake and its implication for management. In: Sengupta M, Dalwani R (eds) Proceedings Taal 2007: the 12th world lake conference, Jaipur, India, pp 1542–1556
Lohse KA, Hope D, Sponseller R, Allen JO, Grimm NB (2008) Atmospheric deposition of carbon and nutrients across an arid metropolitan area. Sci Tot Environ 402:95–105
Luijn FV, van der Molen DT, Luttmer WJ, Boers PCM (1995) Influence of benthic diatoms on the nutrient release from sediments of shallow lakes recovering from eutrophication. Water Sci Technol 32:89–97
Mackereth FJH (1963) Some methods of water analysis for limnologists. Scientific publication no 21. Freshwater Biological Association, Ambleside
Maiti SK (2001) Handbook of methods in environmental studies, vol 1. Water and wastewater. ABD Publisher, Jaipur, India
McEachern P, Prepas EE, Gibson JJ, Dinsmore WP (2000) Forest fire induced impacts on phosphorus, nitrogen and chlorophyll a concentrations in boreal subarctic lakes of northern Alberta. Can J Fish Aquat Sci 57:73–81
Michel P (1984) Ecological Methods for field and laboratory investigation. Tata McGraw-Hill Publ Comp, New Delhi, India
Monteith DT, Stoddard JL, Evans CD, de Wit HA, Forsius M, Hogasen T, Wilander A, Skjelkvale BL, Jeffries DS, Vuorenmma J, Keller B, Kopacek J, Vesely J (2007) Dissolved organic carbon trends resulting from changes in atmospheric deposition chemistry. Nature 450:537–541
O’ Reilly CM (2006) Seasonal dynamics of periphyton in a large tropical lake. Hydrobiologia 553:293–301
Pandey J (2011) The influence of atmospheric deposition of pollutant elements on cross-domain causal relationships at three tropical freshwater lakes of India. Lakes Reserv Res Manag 16:111–119
Pandey J, Pandey U (2001) The influence of catchment on ecosystem properties of a tropical freshwater lake. Biotronics 30:85–92
Pandey J, Pandey U (2009) Microbial processes at land water interface and cross-domain causal relationships as influenced by atmospheric deposition of pollutants in three freshwater lakes in India. Lakes Reserv Res Manag 14:71–84
Perakis SS, Hedin LO (2002) Nitrogen loss from unpolluted South American forests mainly via dissolved organic compound. Nature 145:416–419
Porcal P, Koprivnjak JF, Molot LA, Dillon PJ (2009) Humic substances-part 7: the biogeochemistry of dissolved organic carbon and its interactions with climate change. Environ Sci Pollut Res Int 16:714–726
Schipper LA, Dodd MB, Fisk LM, Power IL, Parenzee J, Arnold G (2011) Trends in soil carbon and nutrients of hill-country pastures receiving different phosphorus fertilizer loadings for 20 years. Biogeochemistry 104:35–48
Singh KP, Tripathi SK (2000) Impact of environmental nutrient loading on the structure and functioning of terrestrial ecosystems. Curr Sci 79:316–323
Sowerby A, Emmett BA, Williams D, Beier C, Evans CD (2010) The response of dissolved organic carbon (DOC) and the ecosystem carbon balance to experimental drought in a temperate shrub land. Euro J Soil Sci 16:697–709
Tate KR, Ross DJ, Filtham CW (1998) A direct method to estimate soil microbial biomass C: effect of experimental variables on some different calibration procedure. Soil Biol Biochem 20:329–335
Tilman D, Fargione J, Wolff B, Antonia CD, Dobson A, Howarth R, Schindler D (2001) Forecasting agriculturally driven global environmental change. Science 292:281–284
Tranvik LJ, Jansson M (2002) Climate change: terrestrial export of organic carbon. Nature 415:861–862
Vadeboncoeur Y, Jeppesen E, Zanden MJV, Schierup H–H, Christoffersen K, Lodge DM (2003) From Greenland to green lakes: cultural eutrophication and the loss of benthic pathways in lakes. Limnol Oceanogr 48:1408–1418
Voghe AI (1971) A Textbook of quantitative inorganic analysis, 4th edn. The English Language Book Society/Longman, Essex
Wedin DA, Tilman D (1996) Influence of nitrogen loading and species composition on the carbon balance of grasslands. Science 274:1720–1723
Wetzel RG (2001) Limnology: lakes and river ecosystems. Academic Press, San Diego
Weyhenmeyer GA, Karlsson J (2009) Nonlinear response of dissolved organic carbon concentrations in boreal lakes to increasing temperature. Limnol Oceanogr 54:2513–2519
Worrall F, Burt TP, Adamson JK (2006) Trends in drought frequency: the fate of DOC export from British peatlands. Clim Chang 76:339–359
Yang Y, He Z, Lin Y, Phlips EJ, Yang J, Chem G, Stoffella PJ, Powell CA (2008) Temporal and spatial variations of nutrients in the Ten Mile Creek of South Florida, USA and effects on phytoplankton. J Environ Monit 4:508–516
Zade M, Ray SS, Dutta S, Panigrahy S (2005) Analysis of run-off pattern for all major basins of India derived using remote sensing data. Curr Sci 88:1301–1305
Acknowledgments
We sincerely acknowledge Prof. H. R. Tyagi, formerly Convener, Department of Environmental Science, M. L. S. University for supports identifying sites and for facilities at the first stage of the study; Prof. R. C. Tewari (Maharana Pratap University of Agriculture and Technology), for help in catchment soil processing and Dr. B. Pandey (Banaras Hindu University) for geological classification and suggestions.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Pandey, U., Pandey, J. Impact of DOC trends resulting from changing climatic extremes and atmospheric deposition chemistry on periphyton community of a freshwater tropical lake of India. Biogeochemistry 112, 537–553 (2013). https://doi.org/10.1007/s10533-012-9747-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-012-9747-7