
Abstract
China is facing an unprecedented set of challenges in balancing the effects of economic development and global climate change with environmental protection and maintaining biodiversity. Although positive steps have been undertaken to remedy this situation, currently 80% of China’s 25 extant primate species are threatened, 15–18 species have population sizes of less than 3000 individuals, and two species of gibbons and one species of langur have been extirpated over the past few decades. Today, virtually all species of primates in China inhabit fragmented landscapes and are distributed in small isolated subpopulations with limited opportunities to exchange individuals or genetic information. Here we present a historical framework examining how human-induced environmental changes, particularly since the second half of the 20th century, accelerated primate population decline in China. In addition, we modeled the expected spatial conflict between agricultural expansion and primate distributions over the next 25–75 years and assessed the current overlap between protected areas and primate distributions. Depending on the assumptions of the spatial conflict model, primate distributions are expected to decline by an additional 51–87% by the year 2100. Thus, unless large-scale conservation policies are implemented immediately the current trend of primate population decline, local extirpation, and species extinctions will accelerate. To mitigate against such extinction scenarios, we advocate the creation of a Chinese national agency and repository of environmental information focused on public awareness and education, the implementation of targeted programs of habitat restoration designed to return impacted forests to a more natural state especially within and at the boundaries of nature reserves, the establishment of additional protect areas, and the construction of a latticework of corridors connecting isolated primate subpopulations. This comprehensive approach offers the most effective way to protect China’s animal and plant biodiversity, including its endangered primate populations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The world’s nonhuman primate populations (hereafter referred to as primates) are facing an impending extinction crisis, with almost 60% of all species listed as Vulnerable, Endangered, or Critically Endangered (Estrada et al. 2017, 2018). This includes several taxa with fewer than 1000 individuals remaining in the wild. The two world regions at most immediate risk are Madagascar, with 97% of 100 primate species threatened and 97% of populations declining and Asia, with 73% of 119 primate species threatened and 95% of populations either declining, or without sufficient information to make an assessment (Estrada et al. 2017, 2018). These two regions include 15 of the 25 most endangered primate species in the world (Schwitzer et al. 2015).
Here we focus on the impending extinction crisis of primates in China (including Taiwan), a country with high animal and plant biodiversity (Jiang et al. 2015). China is the second-most primate diverse country in Asia and is home to four subfamilies, seven genera, and 25 species of primates including lorises, macaques, langurs, snub-nosed monkeys, and gibbons (Table 1). Thirty six percent of these species are considered endemic and 6 species are listed as critically endangered (Fig. 1, http://www.iucnredlist.org/). Distinct from many other primate habitat countries, China is characterized by a large and growing economy (2nd largest economy—IMF’s World Economic Outlook; www.worldbank.org), an expanding middle class, a large cadre of outstanding scientists, and ranks 12th among 90 primate habitat countries worldwide in its Human Development Index (HDI or a measure life expectancy, education, and income as calculated by the United Nations Development Program, http://hdr.undp.org/en/composite/HDI). Thus, China has both the human capital and economic resources to promote effective policies of conservation and play a leading role in sustainably managing wild animal populations (Wei et al. 2015). However, in moving from a largely agrarian economy to a highly industrial economy, China has dramatically transformed its natural environment in ways that have negatively impacted biodiversity and species survivorship (Laurance 2017; Liu 2010; Ren et al. 2015). Although human-induced environmental change has occurred over a period of several 1000 years, over the past few decades China has constructed over 4.5 million kilometers of roads, approximately 23,500 km of rail for high speed trains, 50% of the worlds large dams and from 2003 to 2014 built 70 million new apartments (Fang et al. 2018; New and Xie 2008; Wen 2016; www.economist.com/blogs/graphicdetail/2017/09/daily-chart; www.statista.com/statistics/242963/number-of-newly-built-apartments-in-china/).

Images of the six critically endangered Chinese primates
Today, China faces significant environmental challenges in attempting to balance development and conservation in response to its large human population (approaching 1.4 billion, with an average density of 147 people/km2; www.worldbank.org), long history of agricultural production and deforestation, recent expansion of urban centers (from 13% of the population in 1952 to 51.2% in 2011; Liu 2013) (Fig. 2), extensive water, soil, and air pollution, massive energy needs, and the fact that almost one-third of its land area is susceptible to desertification (Ahrends et al. 2017; Fang et al. 2018; Liu 2013; Qiu 2010; Tang et al. 2015; Xu et al. 2017). This has resulted in 80% of China’s 25 extant primate species listed as threatened, with 92% of populations declining or status unknown (IUCN Red List 2017, http://www.iucnredlist.org/) (Table 1). Unless immediate action is taken, many, if not most, of China’s primate species will become extinct or effectively extinct by the end of this century (Fan et al. 2013; Ren et al. 2015).

Changing trends from rural to urban residence in China
Based on an extensive review of the literature, we examined the social, demographic, economic, and ecological factors that have contributed to the decline of natural forests and primates in China. We also integrated databases on primate conservation from the International Union for the Conservation of Nature (IUCN), with information on agricultural production and expansion from the Food and Agricultural Organization of the United Nations (FAO) and the World Bank, information on the location of protected reserves and national parks from the Protected Planet (https://www.protectedplanet.net/country/CHN), and information from the Integrated Model to Assess the Global Environment (IMAGE, http://themasites.pbl.nl/models/image/index.php/Agricultural_economy) to (1) describe the historical and current set of anthropogenic drivers of primate population decline; (2) present an up-to-date evaluation of the conservation status and risk of extinction of China’s primates; and (3) propose a framework and a series of initial steps that can be taken to increase the long-term survivability of primates in China.
Drivers of primate population decline
Historical context
A series of recent articles (Li et al. in preparation; Liu 2013; Pan et al. 2016; Zhao et al. 2018) provide a historical summary of the set of human activities (political, social, cultural, and economic) and environmental changes that have significantly impacted biodiversity in China over the past several 1000 years. Based on archaeological evidence, 7000–9000 years ago, China experienced significant changes in patterns of land use and food production associated with animal and plant domestication, sedentism, and a major expansion of its human population. Thus, by the start of the Han Dynasty, ~ 2200 years ago, there was an estimated 60 million people in China (Li et al. 2009; Liu and Chen 2012; Liu et al. 2013). Beginning with the Ming Dynasty (1368 to 1644 ad), followed by the Qing Dynasty (1644 to 1912 ad), China’s modern identity as a unified multiethnic state had emerged (Ho 1967; Lee and Wang 1999; Li et al. 2002; Zhao 2006). During these periods northern, central and southern China became the most densely populated and technologically advanced regions, while the Qinghai-Tibet Plateau and northwestern China remained the least populated and least technologically advanced (Lin et al. 2010; Waley-Cohen 2004; Wang et al. 2014).
Although quantitative information on China’s primate biodiversity and distribution prior to the 1970’s is extremely limited, there exists some 2000 years of written records from local gazetteers, dynastic histories and inventories, poems, paintings and other historical sources that provide insights into species presence and distributions (Fan 2017; Zhao et al. 2018). Clearly these records must be viewed with caution as local names for individual species or taxa may vary or change over time. That said, information on some taxa such as the golden snub-nosed monkeys (which have a distinctively blue colored face and long golden pelage) and gibbons (which are anatomically and behaviorally distinct from monkeys) indicates that especially over the past 400–500 years, wildlife hunting and human transformation of forested landscapes for agricultural development and extraction of forest products have led to loss of habitat, elevational shifts in species’ distributions, and primate population decline and extirpation (Fan et al. 2017a, b; Li et al. 2002; Zhao et al. 2018; Zhou and Zhang 2013). These data also indicate the loss of primate populations in the northern and eastern parts of China, with the southwestern and northwestern provinces continuing to maintain the highest primate diversity (Fig. 3).

Number of primate species present in individual provinces in China. The most specious provinces are Yunnan with 15 primate species, Tibet with 8 primate species, Guangxi with 8 primate species, and Quizhou with 6 primate species
Here, we focus on environmental transformations that have occurred in China since the end of World War II. These are rooted in a series of social, political and economical decisions under the system of Monopolized Lands, Monopolized Industry, and Monopolized Commerce that began in the early 1950s (Meng 2015). In 1951 and 1952 China initiated a campaign of Land Reform in which agricultural lands were nationalized and redistributed to some 300,000,000 peasant farmers who were given the right to plant and alter the land, but were not granted ownership. These transactions were enacted by the Village Peasant’s Association and by 1956, approximately 43% of the nation’s cultivated lands had changed from private to government ownership (Bramall 2004; Huang 1995). A second nationalized land ownership program was completed in 1958 with the establishment of more than 50,000 People’s Communes. Organizationally, these communes were considered as basic units of government and their number increased to 75,000 by 1963 (Ahn 1975; Crook 1978). This policy returned lands that had been allocated to peasants during the period of Land Reform back into government control, and initiated a series of People’s Movement/Campaigns designed to more effectively direct social and environmental policy and economic reform.
In 1953, China also embarked upon the first of several Five-Year Plans (1953–1957) targeted at a “socialist transformation” in which the government controlled industry and commerce through the “joint state-private” enterprise program. This led to the nationalization of major economic sectors and the diminishing role of private ownership until 1979, when new economic reforms were enacted (Meng 2015; Zhu 2015). The Great Leap Forward (1958–1959), and “Steel First” (1958) campaigns resulted in nationwide deforestation to fuel inefficient backyard steel producing furnaces; the Learn from Dazhai agricultural program (1963–1976) along with the policy of “Grain First” (1960–1976) were designed to greatly expand the availability of arable lands; and the “Economic Reform”, “Economy First”, and the “Made in China” policies (from the 1980s to the present) promoted the conversion of large tracts of forest to farmland or to provide space for industry, infrastructure, and urbanization (Ho 2003; Pan et al. 2016).
Coincident with the nationalization of land, industry, and commerce during the period from the 1950s to 1980s, over 30 government forestry bureaus were established for the purposes of logging and the expansion of the timber industry (Ministry of Forestry 1987; Sun et al. 2005). This led to the loss of 1.5 million ha of forest, in addition to losses incurred by the activities of private companies and illegal loggers (Zhang 2000). In many cases, illegal logging was targeted to areas of primary forest (Economy 2007). Today, China plays an expanded role in the timber industry worldwide, both as a producer and a consumer of high-quality value-added wood products. According to Sun et al. (2005), China’s timber resources are distributed principally in three major regions: northeast, southwest, and south China, including the provinces of Sichuan, Yunnan, Guangdong, Guangxi, Hunan, Hubei, Fujian, Jiangxi, Guizhou, Zhejiang, Anhui, and Hainan. Many of these areas coincide with zones of high primate diversity (Liu et al. 2013).
In the 1950’s China embarked on an aggressive campaign to develop a rubber industry in its tropical and subtropical regions, principally the provinces of Hainan, Fujian, Guangdong, Guangxi, and Yunnan. In 2001, China emerged as the world’s largest consumer and exporter of rubber, with 4450 km2 of land (8.6% of the world’s total) converted from forest to rubber plantations (Kou et al. 2015; Viswanathan 2007; Ziegler et al. 2009). In 2004, Yunnan Province accounted for 41% of China’s rubber production and Hainan accounted for 55% (Viswanathan 2007). More recently, Yunnan, China’s most biodiverse province and home to the greatest number of primate species (Fig. 3), accounted for 47.9% of the total rubber output in China (Qiu 2009). Rubber plantations now cover 20% of the Xishuangbanna prefecture in southern Yunnan (Qiu 2009), significantly altering ecosystem functions and reducing biodiversity (Hu et al. 2008; Pan 1986). Since the year 2000, the area of land devoted to rubber production in China has increased by almost 60% and industrial round wood extraction from planted forests has tripled (http://www.fao.org/faostat/en/#data). In contrast, land devoted to food crops and habitat conversion for grazing domesticated animals (pasture) has stabilized over the past several years (http://www.fao.org/faostat/en/#data), as China has increased imports of meat and agricultural products (Gale et al. 2015).
China’s primate populations
China is characterized by a taxonomically diverse radiation of 25 primate species including lorises, macaques, langurs, snub-nosed monkeys, and gibbons (Table 1). Nine species are considered endemic. One of these, Macaca leucogenys, is located in China as well as in a south-eastern region of Tibet (Fan et al. 2017b; Li et al. 2015a, b). Virtually all of China’s primate species are included on the Red List of China’s Vertebrates (RLCV) Designated for Legal Protection and 80% of China’s primates are listed as threatened (Vulnerable, Endangered or Critically Endangered) on the IUCN Red List (Table 1). According to the IUCN data base, 92% of Chinese primate species are either declining (76%) or their status is unknown (16%) (Table 1). The populations of two species, the Taiwanese macaque (Macaca cyclopis) and the Hainan gibbon (Nomascus hainanus), are considered stable. In the case of the Hainan gibbon, the current population size is only 26 individuals (Fan 2017; Turvey et al. 2017). In the 1960’s, the population size was estimated at 2000 individuals (Liu et al. 1984). By the 1980’s there were only ten Hainan gibbons remaining in the wild (Bryant et al. 2016). The population increased to approximately 20 individuals by 2003 (Fellowes et al. 2008) and this positive trend has continued. The entire Hainan gibbon population presently resides in an area of some 15 km2 within the 300 km2 Bawangli National Nature Reserve (Turvey et al. 2017). However, Fellowes et al. (2008) report that “the current area of continuous primary forest available to the gibbons is just 1600 ha” (16 km2), and Zhang et al. (2010) indicates that between 1991 and 2008, 7% of the remaining forest suitable for gibbons within the protected reserve was lost. And, although there exists some 80 km2 of nearby suitable habitat this species could expand into, these forests are not currently part of a nature reserve and remain unprotected (Fan and Bartlett 2017). The Critically Endangered Cao-vit gibbon (Nomascus nasutus) also has a current population size of < 30 individuals in China (Turvey et al. 2015; Zhou et al. 2005; Table 1), whereas the endemic and Critically Endangered Skywalker gibbon (Hoolock tianxing) numbers less than 200 individuals (Table 1). And, although, the Skywalker gibbon potentially has access to a forested area of some 108,000 km2, only 3% of that area is protected (Fan and Bartlett 2017). Given the small size, isolated and disjunct distribution of the few remaining gibbon populations in China, without immediate intervention, extinction is imminent (Fan et al. 2017a, b; Turvey et al. 2017). We note that during the past 20 years, two gibbon species, the northern white-cheeked gibbon (Nomascus leucogenys) and the white-handed gibbon (Hylobates lar yunnanensis) have been extirpated from China. These species are present in other parts of Asia, however their conservation status is listed by the IUCN as Critically Endangered or Endangered (Grueter et al. 2009; Fan et al. 2014a; Table 1).
We were able to document one primate species in China whose population appears to have increased relative to IUCN estimates. A recent study by Zhao et al. (2018) indicates that compared with population estimates of 2000 black-and-white snub-nosed monkeys (Rhinopithecus bieti) based on surveys conducted between 1987 and 1992 (Long et al. 1994), the current population numbers are close to 3000 individuals. Although there is evidence of increases and decreases in herd size across its range, the primary factor resulting in an overall increase in black-and-white snub-nosed monkey population size is a decrease in anthropogenic disturbance including hunting and deforestation (Zhao et al. 2018).
At present primates occur in 21 of China’s 34 provincial-level administrative units, with four provinces in west and southwest China (Yunnan = 15 species, Guangxi = 8 species, Tibet = 8 species, Guizhou = 6 species), containing the greatest biodiversity (Fig. 3). Yunnan province, which has an area of 394,000 km2 (4.1% of the total land surface of China), includes 60% (15/25) of China’s primate species and therefore is an area of extreme conservation priority. Unfortunately, human-induced changes to the landscape, exacerbated by the destruction of forest cover and global climate change in Yunnan and its neighboring provinces of Guizhou, Guangxi and Sichuan, have resulted in the most severe droughts recorded in recent decades, negatively impacting fauna and flora, as well as the livelihoods of local residents (Qiu 2010). Moreover, most primate species in China have a highly circumscribed distribution. This is evidenced by the fact that 76% of species are restricted either to a single province (N = 17) or to two provinces (N = 2), and only five species (four species of macaques and one species of snub-nosed monkey, Rhinopithecus roxellana) range into as many as four or more provinces (Table 1). We acknowledge that in many instances only a limited part of a species distribution may extend into a given province or a species entire range may be located in only a small area within a province. For example, François’ langurs (Trachypithecus francoisi) are currently found in only 10 counties across Guangxi Province covering a disjunct and discontinuous area descreased 516 km2 (Wu et al. 1987; Ye 1993). The population of this species has decrease in size by 90% since the 1980s (Li et al. 2007). In Chongqing municipality there remain only 200 François’ langurs, and they are limited to three isolated patches encompassing an area of 57 km2 (Han et al., 2013). Similarly, although the distribution of the black-and -white snub-nosed monkey (Rhinopithecus bieti) includes the provinces of Yunnan and Tibet, the three Tibetan subpopulations are estimated to contain a total of 300–400 individuals and range over an area of 971 km2, which is less than 1% of Tibet province (Xiang et al. 2007). In the case of the Cao-Vit gibbon (Nomascus nasutus), their present day distribution in China is restricted entirely to a single country (Jingxi) within Guangxi Province on the border with Vietnam (Chan et al. 2008). However, focusing on primate distributions and conservation priorities at the provincial level is important because provincial reserves account for 38% of China’s protected areas (Ren et al. 2015), and the actions of provincial leaders can result in the loss or expansion of local animal and plant communities.
Recent population estimates are available for 23 of China’s 25 primate species (no estimates are available for Macaca leucogenys and Trachypithecus pileatus, but their populations are assumed to be extremely small, Table 1). Of these 23 species, 13 (56.5%) contain fewer than 1000 individuals remaining in the wild, and for three additional species population estimates range from 1200 to < 3000 individuals (Table 1). Primates with less than 1000 individuals in China include two species of crested gibbons (Nomascus nasutus and N. hainanus, each with < 30 individuals), the recently named Skywalker Gibbon (Hoolock tianxing, < 200 individuals; Fan et al. 2017a), the pygmy loris (Nycticebus pygmaeus, < 100 individuals), the Arunachal macaque (Macaca munzala, < 500 individuals, with no population residing in a protected reserve), the northern pig-tailed macaque (M. leonina, < 800 individuals), two species of snub-nosed monkeys (Rhinopithecus strykeri with approximately 200 individuals and R. brelichi, each with < 700 individuals), and three species of langurs (Trachypithecus leucocephalus and Semnopithecus schistaceus, each with approximately 1000 individuals, and Trachypithecus shortridgei with < 400 individuals) (Table 1). Although some of these species also are present outside of China (e.g., Nycticebus pygmaeus, Nomascus nasutus, R. strykeri, and M. munzala), most are Endangered or Critically Endangered, and except for the pygmy loris, which is listed by the IUCN as Vulnerable (IUCN 2017), the non-Chinese populations of these species are estimated to be < 500 individuals (Table 1).
Seven primate species in China have population sizes greater than 3500 individuals (Table 1). Five of these are species of macaques. The two largest primate populations remaining in China are Macaca cyclopis, which is present only on the island of Taiwan and has an estimated population size of ~ 300,000 and Macaca mulatta with an estimated population size of 96,000 (Lu et al. 2018; Zhang et al. 2002). The relatively large number of Taiwanese macaques is likely due to a series of strict wildlife conservations laws effectively imposed in Taiwan in 1989 (Agoramoorthy and Hsu 1999) as well as the fact that the species is present in several nature parks and ecotourist sites (Hsu et al. 2009). In the case of Chinese rhesus macaques (M. mulatta), they exhibit a fragmented distribution across some 20 provinces that includes a range of habitats and nature reserves (Lu et al. 2007). However, whereas some regions contain large populations other areas may contain only 2000 individuals (e.g., Taihangshan Mountains National Nature Reserve; Lu et al. 2007). All other species of China’s primates are distributed in small, widely scattered and isolated subpopulations (Li et al. 2015a, b; Lu et al. in press). For example, black-and-white snub-nosed monkeys are currently distributed across 5 isolated subpopulations, four of which contain fewer than 450 individuals (Li et al. 2015a, b; Zhao et al. 2018). Similarly, the remaining population of approximately 1000 white-headed langurs is distributed across as many as 15 isolated subpopulations inhabiting limestone habitats in Guangxi Province (Huang et al. 2002; Wang and Yao 2017). Some 550 individuals reside in Fusui country and an additional 300 individuals reside in Chongzu county (Wang and Yao 2017). These populations are isolated from one another by 40–50 km of roads, villages, and farmland. Similarly, the clearing of forests for agriculture, forest fragmentation, and hunting, especially over the past several 100 years have resulted in the extirpation of gibbons from eastern and northern China (Fan 2017). Today, extremely small and remnant gibbon populations remain only in Hainan, Yunnan, and Guangxi provinces (Fan 2017). The total combined population of the four remaining gibbon species in China is less than 1500 individuals.
In the case of the endemic Tibetan macaque (Macaca thibetana), recent census data indicate that habitat fragmentation has resulted in significant population declines, local extirpation in several areas, and an increase in the number of isolated subpopulations across the species distribution (Sun et al. 2010; Wang et al. 2016a, b). These subpopulations range in size from 35 to > 600 individuals (Wang et al. 2015, 2016a, b; Zhang et al. 2005, 2008a, b; Li et al. 2015a, b; Zhao et al. 2016). Sun et al. (2010) report that given mtDNA haplotype differences across the major Tibetan macaque subpopulations, each is best considered a distinct conservation unit that will require individualized management plans to ensure survivorship. Moreover, rates of genetic heterozygosity in Tibetan macaques are approximately three times lower than that reported in Chinese rhesus macaques (Fan et al. 2014b).
Similarly, Ni and colleagues (unpublished data) found that much of the remaining population of 800–1200 Chinese Bengal slow lorises (Nycticebus bengalensis) is distributed in 29 isolated forest fragments spread across 27 counties and eight prefectures in Yunnan Province. Sixty-five per cent of these forests patches contained less than 30 individuals, and none contained more than 100 individuals. Although there are no published data on fragment size, individual forested patches occupied by these lorises are small (e.g., generally less than 1 km2; Ni, unpublished data) and embedded in a mosaic of disturbed habitat types. An analogous situation exists for François’ langur. Prior to 1980, the population of Trachypithecus francoisi in China was estimated at 8000–10,000 individuals. By 2007, however, the population was reduced to approximately 1000 individuals distributed across three geographically isolated subpopulations (Li et al. 2007). The subpopulation in Guangxi Province contains some 300 individuals and the subpopulation in Guizhou Province contains < 700 individuals (Li et al. 2007).
Three of four species of snub-nosed monkeys (Rhinopithecus roxellana, R. bieti, and R. brelichi) present in China are endemic (R. strykeri also is present in Myanmar and is listed as Critically Endangered) and each is listed by the IUCN as Endangered. Ancestral snub-nosed monkeys appear to have radiated some 2 million years ago in association with the uplifting of the Qinghai-Tibetan Plateau and glacial advances and retreats associated with changes in paleoclimate (Liedigk et al. 2012, Zhou et al. 2014). Today, all species of Chinese snub-nosed monkeys inhabit high altitude deciduous broad-leaf and conifer evergreen forests between 1000 and 4500 m (Grueter 2013; Li et al. 2002; Zhou et al. 2014). Beginning in 2007, a research team led by the Institute of Zoology, Chinese Academy of Sciences in Beijing launched a conservation genetics project focused on understanding the demographic population history of R. roxellana, R. bieti, and R. brelichi. With the discovery in 2010 of a new Rhinopithecus species, R. strykeri (Myanmar or black snub-nosed monkey, with a total population conservatively estimated at approximately 200 individuals in China and 200 in Myanmar; Geissmann et al. 2011; Long et al. 2012; Meyer et al. 2017), whole genome sequence analyses of all four species indicate that population heterozygosity ranges from 0.015 to 0.068%, which is lower than the values reported for Endangered great apes (0.065–0.178%), and similar to that reported for Endangered carnivores such as the Amur tiger (formerly Siberian tiger, Panthera tigris), African lion (Panthera leo), and the snow leopard (Panthera uncia) (0.0073–0.048%) (Zhou et al. 2014, 2016). The diversity of the mitochondrial genome in snub-nosed monkeys also is extremely low, implying a strong decrease in effective female population size (Hong et al. 2017; Kolleck et al. 2013) leading to an increased risk of extinction. A study by Li et al. (2002) documented that over the past 400 years, all populations of snub-nosed monkeys inhabiting forests below an elevation of 800 m across 11 Chinese provinces have gone extinct. Coupled with estimates that more than 80% of the present distribution of snub-nosed monkeys includes areas of unsuitable habitat (Li et al. 2002; Zhao et al. 2018), places this iconic primate radiation at significant risk.
Hunting and illegal and legal primate trade
Given these primate population declines, illegal trading, hunting and the legal capture of primates for biomedical research have only exacerbated conservation challenges. There continues to exist a large network of wildlife smugglers in southwestern China, principally concentrated in Yunnan, Qinghai, and Guangxi provinces, a region of high primate diversity (Zhang et al. 2008a, b). According to Li and Wang (1999), primate species that are represented in this illegal pet trade include the Bengal slow loris, the pygmy loris, several species of macaques (Macaca assamensis, M. mulatta, M. leonina, M. arctoides), as well as gibbons (Nomascus leucogenys/N. gabriellae and Hoolock spp). Lorises in China are illegally harvested for traditional medicine and the pet trade. More than 300 lorises, mostly wild-caught or cross-border imported, have been rescued or confiscated in China in the last ten years (CITES Trade Data Base). There also are an estimated 200 captive lorises in China, the majority of which are housed as pets for the purpose of breeding and illegal trade (Ni Qingyong unpublished data; Nijman et al. 2014). In addition, the bones and other body parts of virtually all primate species are traded as traditional medicine at local markets (Li and Wang 1999; R. Pan pers. obser.) and, the hunting of langurs for use in traditional medicine in China remains a significant threat to their survival (Li et al. 2007; Nadler 2016a, b). In some cases, direct hunting is less of a problem than indirect hunting (e.g., snares aimed at capturing musk deer also catch primates, Gray et al. 2018; Grueter pers. observ.). A recent report by Meyer et al. (2017) indicates that over the past 20–30 years, approximately 80 Critically Endangered black snub-nosed monkeys were hunted in Myanmar and an additional 19 were killed in China. However, both countries have implemented measures to reduce or eliminate hunting in protected reserves (Ren et al. 2015). Finally, a review of 12 years of seizure records (1996–2008) compiled by TRAFFIC (including almost 200,000 live animals) indicated that over 4000 live primates were confiscated along with 13 seizures that contained primate body parts and products (Rosen and Smith 2010). Overall, 11% of the confiscated specimens were from China.
Since the 1980’s several species of macaques, Macaca mulatta, M. arctoides, M. fascicularis, M. assamensis, and M. leonina, have been legally imported into or exported from China for biomedical research (Fan and Song 2003). Monkeys captured from the wild are used either to establish new breeding centers, or for export (Fan and Song 2003; Hao 2007; Jiang et al. 2008). Hao (2007) indicates that during the early part of the 21st century, China was exporting approximately 20,000 primates per year, and by 2008 there were 40 primate-breeding centers in the country containing some 40,000 individuals, mostly rhesus macaques (Cyranoski 2016; Jiang et al. 2008). Since India banned primate exports in 2013 (Behal 2014), China has steadily increased the number of captive-bred and exported primates. In 2014, the total number of crab-eating macaques (M. fascicularis) and rhesus macaques (M. mulatta) in captivity in China reached ~ 250,000 and ~ 42,000, individuals respectively, with M. fascicularis accounting for more than 90% of primates exported (CITES Trade Data Base). Between 2010 and 2016 alone, China legally exported more than 350,000 M. fascicularis and 35,000 M. mulatta, principally for biomedical research (CITES Trade Database). Unfortunately, coinciding with legal trade, there has been an increase in the illegal primate trade across China’s borders. For example in 2006, 197 crab-eating macaques were confiscated at the Vietnam-China border (Hao 2007). Finally, Nijman (2010) reports that based on the WCMC-CITES database, 77% of trade in wild-caught mammals from Southeast Asia during the period of 1998–2007 came from China, with Japan and the European Union the most frequent importers. Reducing wildlife trade and the legal and illegal networks that profit from this trade will play an important role in protecting threatened primate populations (Estrada et al. 2017, 2018).
Deforestation and forestation
Over the past 50 years, China has experienced greater environmental change and rates of deforestation than most other major countries, such that by 2005 it ranked 133rd among the 146 countries whose environmental sustainability was evaluated (Liu 2010). Today, China is one of the world’s most forest deficient countries, with only 0.2 ha of forest cover/person, compared with a world average of 0.6 ha, and forests cover approximately 20% of the country’s land area compared with 31% worldwide (Dai et al. 2013; Liu et al. 2013). We caution that data on deforestation, reforestation, and land use patterns are subject to error, as these are difficult to accurately measure, are reported by governmental agencies which may have a biased perspective, and evaluation based on of satellite imagery can misclassify areas that are fields and agricultural plantations as forests (Wang et al. 2017). A study by Wang et al. (2017) using high resolution images of 1798 random forest points to verify reported habitat assessments across several regions of China found that interpretation accuracy ranged from 63 to 95% (average of 86%).
Notwithstanding some degree of reporting error, a recent evaluation by Ahrends et al. (2017) indicates that China has lost 59–67% of its original forest (1.9–2.7 million km2). Moreover, between 1970 and 1996 China experienced a 2.6 to 3.3% annual deforestation rate (Ren et al. 2015). In an effort to reverse this trend and promote sustainable development, beginning in the late 1990’s China implemented a large reforestation initiative called The Natural Forest Protection Program (NFPP). This effort was intended to conserve and expand forest cover by planting tens of billions of trees and thereby limit environmental damage, promote the sustainable use of forest products, decrease soil erosion on steep slopes, convert land previously used for agriculture into forest (Grain for Green Program), outlaw logging of primary forests, expand protected areas to increase and conserve biodiversity, and increase carbon sequestration and forest cover across the country (Ahrends et al. 2017; Liu 2010, 2013; Ren et al. 2015).
China’s reforesting efforts appear to have met with some measure of success; for example between 2000 and 2014, the land area devoted to food agriculture in China has remained relatively constant, and the area of primary forest appears to have stabilized or slightly increased (http://www.fao.org/countryprofiles/index/en/?iso3=CHN). This resulted in the annual rate of deforestation to drop to 1.1% during the 10 year period from 2000 to 2010 (provinces that participated in the NFPP program had an annual deforestation rate 0.62% whereas provinces that did not participate experienced a deforestation rate of 2.26%; Ren et al. 2015), and the area of tree cover in China to increase by 30,000 km2 per year (Keenan et al. 2015), with a commitment to increase forest cover to 40,000 km2 per year by 2020 (Vina et al. 2016). This is clearly a positive step in land management.
The definition of forest used by China is 20% canopy cover with no specified tree height and no requirement for taxonomic diversity (Ahrends et al. 2017). In this regard, the increase in forest cover over the past decade is best described either as afforestation (planting trees in areas that previously contained no tree cover) or reforestation (planting trees in areas that originally contained forest cover). In both cases, however, generally this has not been part of a focused attempt to regenerate natural ecosystems. Much of this effort has resulted in the replanting of relatively small areas with fast growing monocultures of non-native species for commercial use (e.g., eucalyptus, rubber, pine, and fruit trees; Bremer and Farley 2010; Cao 2008; Xu 2011), the planting of shrubs, and the planting of a relatively limited number of continuous tracts (> 5 ha) of closed canopy forest (> 5 m in height and > 50 canopy cover) (Ahrends et al. 2017). This, coupled with a recent evaluation of reforestation effectiveness indicates that between 2000 and 2010 only “1.6% of China’s territory displayed a significant gain in percent tree cover” (Vina et al. 2016). Moreover, gains in forest cover have tended to occur in areas that originally contained low tree cover, losses in forest cover have tended to occur in regions that initially contained high tree cover (Vina et al. 2016), and in most instances reforested areas are dominated by planted forests rather than native forests (Wang et al. 2017).
The Food and Agriculture Organization of the United Nations (FAO) defines forest as vegetation of at least 0.5 ha in area, trees greater than 5 m in height, and a tree canopy cover of at least 10% (http://www.fao.org/docrep/005/Y4171E/Y4171E10.htm). Based on those criteria, during the 10-year period between 2000 and 2010, China added 434.000 km2 of new forest cover (Ahrends et al. 2017). These forests, however, are unlikely to be suitable for most arboreal primate species, that require large areas of continuous multilevel forest canopies and a taxonomically diverse plant based diet. Using the same criteria but requiring at least 50% of canopy cover, reduces reforestation to an area of some 33,000 km2 (Ahrends et al. 2017). Moreover as suggested by Fan and Bartlett (2017), habitats suitable for gibbons require > 60% forest cover. Thus, despite the large amount of human and financial capital invested by China in increasing forest cover, unless policies are specifically designed to expand the natural ecosystems required by China’s primates and other wildlife populations, the effectiveness of these policies for conservation is limited.
Effects of agricultural expansion on primate survival
In Fig. 4, we examine the predicted effects of future agricultural expansion on primate survivorship and distributions in China. Data for expected agricultural expansion are from the Global Agro-Ecological Zones, a database compiled by the United Nations Food and Agriculture Organization and the International Institute for Applied Systems Analysis (http://www.fao.org/nr/gaez/en/), and species geographical distributions are from IUCN databases (http://www.iucnredlist.org/technical-documents/spatial-data). The IMAGE or the Integrated Model to Assess the Global Environment (http://themasites.pbl.nl/models/image/index.php/Agricultural_economy) was used to identify predicted overlap in agricultural production and primate distributions in each grid cell (0.5° of spatial resolution; Dobrovolski et al. 2013). To model the future distribution of agricultural land, IMAGE considers predictors like previous land cover, potential vegetation cover, crop productivity, management factors, human population density, and demand for food, biofuel and timber products (IMAGE Team 2001). The scenarios used in this IMAGE model represent different pathways of future social and economic human development, which differ in terms of the degree of global change in the demand for goods and services, and anticipated changes in the value of resources, products, and human development (IMAGE Team 2001). Zones of conflict are defined as areas in which primate distributions are predicted to overlap with areas of expanding agriculture. Given the fragmented nature of primate distributions across China, these zones of overlap could involve relatively small-scale agriculture including family farms and village plots on the boundaries or inside protected areas as well as in unprotected areas. Our models predict that even under the most ‘optimistic’ scenario (which presumes a change in society towards cleaner and more resource-efficient technologies that can be shared in a globally integrated world), there is likely to be a 51% reduction in the geographical range of China’s primates by the year 2100. This will have the greatest effect on China’s remaining gibbon and loris populations. Under the business-as-usual model (which assumes continuation of national policies that promote environmental protection), gibbons, lorises, and most langur species will be lost. Under the assumptions of the ‘pessimistic model (which assumes national policies that permit agricultural expansion into currently protected areas), primate distributions across China will shrink by 87%, with only a few taxa, including golden snub-nosed monkeys, black-and-white snub-nosed monkeys and some macaque species surviving in small isolated populations. Given these future scenarios, China needs to redirect and prioritize its environmental policies to protect its extremely vulnerable primate fauna and their habitats.

Forecasted spatial conflict between primate distributions and agricultural expansion on mainland China to the end of the 21st century. Three scenarios are presented for 2050 and for 2100: optimistic, business as usual and pessimistic. Primate species richness is based on data provided by the IUCN (consulted May, 2017) on the geographic ranges of 19 of China’s 25 primate species. Agricultural expansion describes the expansion of crop production predicted by the Integrated Model to Assess the Global Environment (IMAGE) database (IMAGE Team 2001)
A way forward: conservation priorities for China
Our review of the historical, cultural, social, and economic factors that have contributed to deforestation and major environmental transformations across China highlights the unprecedented challenges China faces in providing for its citizens while protecting and conserving its biodiversity and primate populations (Estrada et al. 2017; Fan 2017; Grueter et al. 2009; Li et al. 2002; Pan et al. 2016; Turvey et al. 2015; Zhang et al. 2005). We have reached a moment in which immediate and joint action led by the Chinese government, Chinese scientists, national and international conservation organizations, and the Chinese public is required or, over the next 25–75 years, the overwhelming majority of China’s primate populations will face extinction. A collective effort of all stakeholders is the only effective way to protect and save our closet living relatives and their invaluable contributions to Chinese culture, art, history, ideology religion, and ecosystems health (Estrada et al. 2017; Fan 2017; Pan et al. 2016). If left to continue, the current pattern of human-induced habitat degradation and climate change will only accelerate processes of desertification, loss of plant and animal biodiversity, air and water pollution, and the coastal flooding of large urban centers, each of which are significant threats to the well-being of China’s human and nonhuman primate populations (Ahrends et al. 2017; Piao et al. 2010; Tang et al. 2015).
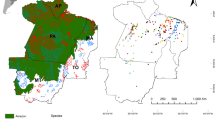
To balance the social benefits of economic growth and the negative costs of the human impact on ecosystem health and biodiversity, the government of China has expanded the number of protected areas. Based on data from the United Nations Environment World Conservation Monitoring Centre (UNEP-WCMC 2017—https://www.unep-wcmc.org), 17.1% of China’s land area (some 1.6 million km2) has been set aside either as National Nature Reserves (N = 428, covering 9.9% of China’s land area), provincial reserves (N = 854 reserves) or local reserves (N = 1307 reserves) to protect habitats for the conservation and maintenance of biodiversity (Xu et al. 2017). The largest of these reserves are located in the provinces of Qinghai and Tibet in the southwest of mainland China (Fig. 5). This is a mountainous region that accounts for 75.3% of the total National Nature Reserve system, and includes 33.6% of China’s mammalian habitats (Xu et al. 2017). Few primate populations are present in these largest reserves (Fig. 5). Primates are present in 418 reserves (185 national-level and 233 provincial-level reserves; Xu et al. 2017). These reserves represent an area of 352,089.4 km2 or approximately 22% of China’s protected areas (https://www.protectedplanet.net/country/CN). And, although China has expanded its system of nature reserves (which has helped reduce deforestation and hunting within reserve boundaries but not in areas adjacent to reserves, Liu 2010; Ren et al. 2015), and has allocated billions of dollars to reforestation, most of these programs are not designed to regenerate native habitats, which are crucial for primate survival (Vina et al. 2016; Ahrends et al. 2017).

Current distribution of primate species richness, protected areas, and forest cover in China
Given the rapid urbanization of China’s human population, we propose that the Natural Forest Protection Program and the National Nature Reserve Program be expanded to benefit primate population recovery and animal and plant biodiversity by focusing on the restoration of natural habitats, especially in rural areas with reduced human populations. In the case of the Critically Endangered Hainan gibbon, a program of reforestation in the Bawangling National Nature Reserve was initiated in 2004 with the goal of converting pine plantations into a matrix of native and nonnative forest (Fellowes et al. 2008). Experience gained from this initiative can serve as a model for reforestation programs in other parts of China, including those targeted at habitat restoration, within and at the boundaries of nature reserves. These projects must prioritize connecting isolated primate subpopulations using a lattice-work of protected corridors that enhance forest connectivity at both altitudinal and transverse levels (Newmark et al. 2017; Townsend and Masters 2015). Expanding suitable habitat is critical. Given their slow life history; a declining primate population can persist in suboptimal forest fragments for decades before eventually succumbing to local extinction. Two recent examples of successfully targeted forest regeneration and restoration programs, one in Brazil and the other in Tanzania (Newmark et al. 2017) provide an instructive model. Newmark et al. (2017) report that in the Eastern Arc Mountains of Tanzania, the planting of 8134 ha of forested corridors intersecting 42 isolated forest fragments (fragment size ranged from 92 to 8000 ha) would create a single continuous forest of 316,663 ha. Similarly, in an area of Atlantic Forest of eastern Brazil, the restoration of 6489 ha of targeted forest corridors designed to connect 29 forest fragments, would result in a continuous forest of 251,952 ha (Newmark et al. 2017). It is argued that the construction of forest corridors “between the largest and the closest forest fragments could provide one of the highest returns on investment worldwide for biodiversity” (Newmark et al. 2017). In addition, the presence of multiple linkage points between forest fragments enable species currently isolated in one or more of these fragments to serve as a source population for migration and gene flow into other parts of the newly created continuous forest (Newmark et al. 2017). Support for the effectiveness of this approach in China comes from a recent attempt to connect the Fanjingshan National Reserve with the Yinjiang Yangxi Provincial Nature Reserve in northeastern Guizhou province, combining the area harboring the entire population of the Guizhou or gray snub-nosed monkey (Rhinopithecus brelichi, Grueter pers. comm.).
Primate species that are high priority for habitat linking (corridors) include the smallest of the remaining golden-snub-nosed monkey subpopulations or herds, which are located in Shennongjia National Reserve, Hubei Province; the five southern subpopulations of R. bieti in Lanping and Yunlong counties which contain 30, 30, 220, 160 and 130 individuals; the subpopulation of Trachypithecus francoisi in central Guangxi Province (population size ~ 300 animals); all three T. leucocephalus subpopulations in central Guangxi Province; and subpopulations of R. strykeri located on the eastern and western slopes of the Gaoligong Mountains in the border area with Myanmar. In some areas or in the short-term, these corridors could include human-made bridges or other constructed zones of safe passage and comprise non-native fast growing tree species. In the long-term, however, corridors must be comprised of native forest communities with the goal of expanding suitable habitat and increasing the size and genetic variability of the primate populations. The restoration of native forest communities also benefit the livelihoods of people in the local human communities, who collect and use a variety of forest products (Dosen et al. 2017; Newmark et al. 2017; Struebig et al. 2015).
A second priority is the creation of a national network to monitor and genotype captive primate populations living in China’s zoos and sanctuaries with the goal of using these individuals as a source population to repopulate and increase genetic variation in the declining wild populations. Urgency must be given to Endangered and endemic taxa, but no wild individuals should be harvested for the purpose of restocking existing captive populations. For a small number of China’s primate species, there exist breeding or ‘rescue’ centers that can play a role in reintroduction (e.g., Rhinopithecus roxellana). For other species, such as R. bieti and R. brelichi, the number of individuals in these facilities is quite small and they are not specifically designed as effective captive breeding programs (Grueter pers. comm.). For three Critically Endangered gibbon species in China (Nomascus nasutus, N. hainanus, and N. concolor) there are no known individuals in captivity and therefore expanding suitable habitat and strict enforcement of existing conservation laws remains an imperative, if these primates are to be saved from extinction (Deng and Zhou 2017; Fan and Bartlett 2017). This also is true for the Critically Endangered black snub-nosed monkey (R. strykeri). There are only two individuals in captivity, both are female, and habitat loss in response to illegal logging and hunting remain the greatest threats to this species existence (Yang et al. submitted).
Finally, to accomplish the goals of conservation and protect China’s Endangered primate populations, we recommend that the Chinese Government, conservation agencies, and leading members of the recently established China Primatological Society (CPS) develop an international conservation centre. This centre would create a highly effective and responsive governance structure that functions as a national archive and shared data repository for information, data management, data analysis, and the implementation of effective conservation policies that promote biodiversity, habitat restoration, and public education based on the most recent advances in climate science, sustainable land use patterns, animal and plant species requirements, and the needs of the local human communities that overlap with primate distributions (Pan et al. 2016). The databases of the centre would be linked with those of large international organizations, such as The World Bank, World Health Organization, IUCN and CITES (the Convention on International Trade in Endangered Species of Wild Fauna and Flora), and shared publically. This would facilitate direct comparison and scientific collaboration with other global regions. This centre would spearhead a national-scale systematic conservation plan to determine the optimized spatial priorities for biodiversity conservation in China (Wu et al. 2014) and collaborate with primate scientists worldwide to develop new technologies and analytical tools to monitor, evaluate, and promote effective conservation programs such as the use of wireless systems to integrate GPS data obtained from drones, satellite imagery, and camera traps to more accurately monitor and model species distributions, group size, and patterns of range use with changes in forest cover, habitat disturbance, the location of agricultural fields, and proximity to human settlements (Estrada et al. 2017, 2018).
In addition the centre would generate a nationwide educational and public information campaign, similar to that used to conserve and protect the giant panda (Wei et al. 2015). The goals of this campaign are to highlight the close evolutionary relationship between human and nonhuman primates, promote the fact that China’s primates are a fundamental part of the country’s cultural and natural heritage, and that almost 40% of China’s primate species are endemic. To this end, several of China’s primate species should be designated as nation-wide and/or province-wide flagship species and be part of a national priority to promote primate biodiversity and conservation. Such programs have been successfully conducted at the local level. For example, the Zoological Society of Shanghai, the NGO Seacology, and the Hainan Environmental Ecological Education Centre initiated a program to educate and increase awareness of the conservation status of the Hainan gibbon (Fellowes et al. 2008). Educational programs targeted to school children in rural communities have proven extremely effective in promoting awareness of primate conservation, especially when these programs have focused on ecosystems health, are relevant to the concerns of the local community, and encourage the direct participation of community leaders in conservation action (Wallis and Lonsdorf 2010). In addition, we are pleased to note, that the Chinese government has recently identified ten animals species as a focus of their national conservation efforts (Zhang and Wang 2018). One of these is the endangered golden snub-nosed monkey (Rhinopithecus roxellana). However, none of the 15–18 primate species with population sizes of less than 3000 are on this list.
In conclusion, 60–72% of China’s 25 primate species have population sizes of less than 3000 individuals, 80% are threatened, only 4% are increasing in numbers, and virtually all species are distributed in small and isolated subpopulations that can no longer exchange individuals or genetic information. Several species live in habitats that have limited suitability for their long-term survivorship (Fan 2017). Across many primate habitat countries, effective governance is a key to promoting laws and policies for effective primate conservation (Estrada et al. 2018). In this regard, China is unique in that it has the economic and intellectual capital and governmental structures required to promote sustainable land use practices. However, unless large-scale and targeted fact-based conservation policies are implemented immediately, the current trend of population decline, local extirpation, and species extinction will be further accelerated. China is at a historic moment and poised to take advantage of its human capital of outstanding scientists and economic prosperity to become a world leader in environmental sustainability, conservation biology, and scientific research, designed to protect its declining primate populations.
References
Agoramoorthy G, Hsu M (1999) CITES implementation through the wildlife conservation law of Taiwan. J Int Wildl Law Policy 2:64–68
Ahn BJ (1975) The political economy of the People’s commune in China: changes and continuities. J Asian Stud 34:631–658. https://doi.org/10.2307/2052547
Ahrends A, Hollingsworth PM, Beckshafer P, Chen H, Zomer RJ, Zhang L et al (2017) China’s fight to halt tree cover loss. Proc R Soc B 284:20162559. https://doi.org/10.1098/rspb.2016.2559
Behal S (2014) Primate export: Ending the monkey business. India today. https://www.indiatoday.in/magazine/indiascope/story/19780228-government-bans-export-of-monkeys-us-forced-to-curtail-experiments-822869-2014-04-29. Accessed 29 April 2014
Bramall C (2004) Chinese land reform in long-run perspective and in the wider East Asian context. J Agrar Change 4:107–141. https://doi.org/10.1111/j.1471-0366.2004.00074.x
Bremer LL, Farley KA (2010) Does plantation forestry restore biodiversity or create green deserts? A synthesis of the effects of land-use transitions on plant species richness. Biodivers Conserv 19:3893–3915. https://doi.org/10.1007/s10531-010-9936-4
Bryant JV, Gottelli D, Zeng X, Hong X, Chan BP, Fellowes JR et al (2016) Assessing current genetic status of the Hainan gibbon using historical and demographic baselines: implications for conservation management of species of extreme rarity. Mol Ecol 25:3540–3556. https://doi.org/10.1111/mec.13716
Cao SX (2008) Why large-scale afforestation efforts in China have failed to solve the desertification problems. Environ Sci Technol 42:1826–1831. https://doi.org/10.1021/es0870597
Chan BPL, Tan XF, Tan WJ (2008) Rediscovery of the critically endangered eastern black crested gibbon Nomascus nasutus (Hylobatidae) in China, with preliminary notes on population size, ecology, and conservation status. Asian Primates J 1:17–25
Crook FW (1978) The commune system in the People’s Republic of China, 1963–1974. Joint Economic Committee, Congress of the United States. Policy Perform 1:366–410
Cui LW, Li YC, Ma C, Scott MB, Li JF et al (2016) Distribution and conservation status of Shortridge’s capped langurs Trachypithecus shortridgei in China. Oryx 50:732–741. https://doi.org/10.1017/S0030605315000319
Cyranoski D (2016) Monkey Kingdom: China is positioning itself as a world leader in primate research. Nature 532:300–302. https://doi.org/10.1038/532300a
Dai LM, Zhao W, Shao GF, Lewis BJ, Yu DP, Zhou L et al (2013) The progress and challenges in sustainable forestry development in China. Int J Sust Dev World 20:394–403. https://doi.org/10.1080/13504509.2013.775193
Deng HQ, Zhou J (2017) Diet composition of Hainan gibbons (Nomascus hainanus) based on 13 years of observation. North-West J Zool e171703
Deng HQ, Zhou J, Yang YW (2014) Sound Spectrum Characteristics of Songs of Hainan Gibbon (Nomascus hainanus). Int J Primatol 35:547–556. https://doi.org/10.1007/s10764-014-9767-3
Dobrovolski R, Loyola RD, Guihaumon F, Gouveia SF, Diniz-Filho JAF (2013) Global agricultural expansion and carnivore conservation biogeography. Biol Conserv 165:162–170. https://doi.org/10.1016/j.biocon.2013.06.004
Dosen J, Raboy BE, Fortib MJ (2017) Restoration strategies to improve connectivity for golden-headed lion tamarins (Leontopithecus chrysomelas) in the Bahian Atlantic Forest, Brazil. Int J Primatol 38:962–983. https://doi.org/10.1007/s10764-017-9991-8
Economy EC (2007) The great leap backward? The cost of China’s environment. Foreign Aff 86:38–59. https://doi.org/10.2307/20032433
Estrada A, Garber PA, Rylands AB, Roos C, Fernandez-Duque E, Di Fiore A et al (2017) Impending extinction crisis of the world’s primates: why primates matter. Sci Adv 3:e1600946. https://doi.org/10.1126/sciadv.1600946
Estrada A, Garber PA, Mittermeier RA, Wich W, Goveia S et al (2018) Primates in peril: the significance of Brazil, Madagascar, Indonesia and the Democratic Republic of the Congo for global primate conservation. Peer J 6:e4869. https://doi.org/10.7717/peerj.4869
Fan PF (2017) The past, present, and future of gibbons in China. Biol Conserv 210:29–39. https://doi.org/10.1016/j.biocon.2016.02.024
Fan PF, Bartlett TQ (2017) Overlooked small apes need more attention! Am J Primatol. https://doi.org/10.1002/ajp.22658
Fan Z, Song YL (2003) Chinese primate status and primate captive breeding for biomedical research in China. In: Institute for Laboratory Animal Research (U.S.). International Perspectives: The Future of Nonhuman Primate Resources: Proceedings of the Workshop Held April 17–19, 2002. The National Academies Press, Washington DC, pp 36–45
Fan PF, Xiao W, Huo S, Ai HS, Wang TC, Lin RT (2011) Distribution and conservation status of Hoolock leuconedys in China. Oryx 45:129–134. https://doi.org/10.1017/s003060531000089x
Fan PF, Fei HL, Luo AD (2013) Ecological extinction of critically endangered northern white-cheeked gibbon (Nomascus leucogenys) in China. Oryx 48:52–55. https://doi.org/10.1017/s0030605312001305
Fan PF, Fei HL, Luo AD, Luo AD (2014a) Ecological extinction of the critically endangered northern white-cheeked gibbon Nomascus leucogenys in China. Oryx 48:52–55. https://doi.org/10.1017/s0030605312001305
Fan ZX, Zhao G, Li P, Osada N, Xing JC, Yi Y et al (2014b) Whole-genome sequencing of Tibetan macaque (Macaca thibetana) provides new insight into the macaque evolutionary history. Mol Biol Evol 31:1475–1489. https://doi.org/10.1093/molbev/msu104
Fan PF, He K, Chen X, Ortiz A, Zhang B, Zhao C et al (2017a) Description of a new species of Hoolock gibbon (Primates: Hylobatidae) based on integrative taxonomy. Am J Primatol. https://doi.org/10.1002/ajp.22631
Fan PF, Liu Y, Zhang ZC, Zhao C, Li C, Liu WL et al (2017b) Phylogenetic position of the white-cheeked macaque (Macaca leucogenys), a newly described primate from southeastern Tibet. Mol Phylogenet Evol 107:80–89. https://doi.org/10.1016/j.ympev.2016.10.012
Fang J, Yu G, Liu L, Shuijin H, Chapin FS III (2018) Climate change human impacts, and carbon sequestration in China. Proc Natl Acad Sci USA 115:4015–4020
Fellowes JR, Chan BPL, Zhou J, Chen SH, Yang SB, Chit NS (2008) Current status of the Hainan gibbon (Nomascus hainanus): progress of population monitoring and other priority actions. Asian Primates J 1:2–8
Gale F, Hansen J, Jewison M (2015) China’s growing demand for agricultural imports. United States Department of Agriculture, EIB-138 Economic Research Service, February 2015
Geissmann T, Lwin N, Aung SS, Aung TN, Aung ZM, Hla TH et al (2011) A new species of snub-nosed monkey, genus Rhinopithecus Milne-Edwards, 1872 (Primates, Colobinae), from northern Kachin state, northeastern Myanmar. Am J Primatol 73:96–107. https://doi.org/10.1002/ajp.20894
Gray TNE, Hughes AC, Laurance WF, Long B, Lynam AJ, O’Kelly H, Ripple WJ, Seng T, Scotson L, Wilkinson NM (2018) The wildlife snaring cirisis” an insidious and pervasice threat to biodiversity in Southeast Asia. Biodivers Conserv 27:1031–1037
Grueter CC (2013) The biology of the snub-nosed monkeys, Douc langurs, proboscis monkeys and simakobu. Hauppauge, Nova Biomedical, New York
Grueter CC, Jiang XL, Konrad R, Fan PF, Guan ZH, Geissmann T (2009) Are Hylobates lar extirpated from China? Int J Primatol 30:553–567. https://doi.org/10.1007/s10764-009-9360-3
Guo YQ, Zhou J, Song XH, Den GHQ, Qiu Y, Shi L et al (2017) The population of Rhinopithecus brelichi in Fanjingshan National Nature Reserve, Guizhou, China. Acta Theriol Sin 37:104–108. https://doi.org/10.16829/j.slxb.201701009
Han ZX, Hu G, Wu SB, Cao CL, Dong X (2013) A census and status review of the Endangered François’ langur Trachypithecus francoisi in Chongqing, China. Oryx 47(1):128–133. https://doi.org/10.1017/S0030605311001396
Hao X (2007) Monkey research in China: developing a natural resource. Cell 129:1033–1036. https://doi.org/10.1016/j.cell.2007.05.051
Ho PT (1967) The significance of the Ch’ing period in Chinese history. J Asian Stud 26:189–195. https://doi.org/10.2307/2051924
Ho P (2003) Mao’s war against nature? The environmental impact of the Grain-First campaign in China. China J 50:37–59. https://doi.org/10.2307/3182245
Hong YY, Duo HR, Hong JY, Yang JY, Liu SM, Yu LH et al (2017) Resequencing and comparison of whole mitochondrial genome to gain insight into the evolutionary status of the Shennongjia golden snub-nosed monkey (SNJ R. roxellana). Ecol Evol 7:4456–4464. https://doi.org/10.1002/ece3.3011
Hsu MJ, Kao CC, Agoramoorthy G (2009) Interactions between visitors and Formosan macaques (Macaca cyclopis) at Shou-Shan Nature Park, Taiwan. Am J Primatol 71:214–222. https://doi.org/10.1002/ajp.20638
Hu HB, Liu WJ, Cao M (2008) Impact of land use and land cover changes on ecosystem services in Menglun, Xishuangbanna, Southwest China. Environ Monit Assess 146:147–156. https://doi.org/10.1007/s10661-007-0067-7
Huang PCC (1995) Rural class struggle in the Chinese revolution representational and objective realities from the land reform to the cultural revolution. Mod China 21:105–143. https://doi.org/10.1177/009770049502100105
Huang CM, Wei FW, Li M, Quan GQ, Li HH (2002) Current status and conservation of white-headed langur (Trachypithecus leucophalus) in China. Biol Conserv 104:221–225. https://doi.org/10.1016/s0006-3207(01)00168-9
IMAGE Team (2001) The IMAGE 2.2 implementation of the SRES Scenarios. A comprehensive analysis of emissions, climate change and impacts in the 21st century. Netherlands Environmental Assessment Agency (MNP) CD-ROM Publication 500110001, Bilthoven
IUCN (2017) http://www.iucnredlist.org/
Jiang ZG, Meng ZB, Zeng Y, Wu ZG, Zhou ZH (2008) CITES no-detrimental finding for exporting rhesus monkeys (Macaca mulatta) from China. NDF Workshop case study. WG5-Mammals 6:1–15
Jiang ZG, Ma Y, Wu Y, Wang YX, Zhou KY, Liu SY et al (2015) China’s mammal diversity and geographic distribution. Science Press, Beijing
Keenan RJ, Reams GA, Achard F, de Freitas JV, Grainger A, Lindquist E (2015) Dynamics of global forest area: results from the FAO global forest resources assessment. For Ecol Manag 352:9–20. https://doi.org/10.1016/j.foreco.2015.06.014
Kolleck J, Yang MY, Zinner D, Roos C (2013) Genetic diversity in endangered Guizhou snub-nosed monkeys (Rhinopithecus brelichi): contrasting results from microsatellite and mitochondrial DNA data. PLoS ONE 8:e73647. https://doi.org/10.1371/journal.pone.0073647
Kou WL, Xiao XG, Dong JW, Gan S, Zhai DL, Zhang GL et al (2015) Mapping deciduous rubber plantation areas and stand ages with PALSAR and landsat images. Remote Sens 7:1048–1073. https://doi.org/10.3390/rs70101048
Laurance W (2017) The dark legacy of China’s drive for global resources. Yale Environment 360. March 28 (http://e360.yale.edu/features/the-dark-legacy-of-china’s-drive-for-globalresources?from=timeline&isappinstalled=0. Accessed 28 March 2017
Lee J, Wang F (1999) Malthusian models and Chinese realities: the Chinese demographic system 1700–2000. Popul Dev Rev 25:33–65. https://doi.org/10.1111/j.1728-4457.1999.00033.x
Li W, Wang H (1999) Wildlife in Yunnan Province, China, at the border with Vietnam. Traffic B 18:21–30
Li BG, Pan RL, Oxnard CE (2002) Extinction of snub-nosed monkeys in China during the past 400 years. Int J Primatol 23:1227–1244. https://doi.org/10.1023/A:1021122819845
Li YB, Huang CM, Ding P, Tang Z, Wood C (2007) Dramatic decline of François’ langur Trachypithecus francoisi in Guangxi province, China. Oryx 41:38–43. https://doi.org/10.1017/s0030605307001500
Li XQ, Dodson J, Zhou J, Zhou XY (2009) Increases of population and expansion of rice agriculture in Asia, and anthropogenic methane emissions since 5000BP. Quat Int 202:41–50. https://doi.org/10.1016/j.quaint.2008.02.009
Li YC, Liu F, He XY, Ma C, Sun J, Li DH, Xiao W, Cui LW (2015a) Social organization of Shortridge’s capped langur (Trachypithecus shortridgei) at the Dulongjiang Valley in Yunnan, China. Zool Res 36:152–160
Li C, Zhao C, Fan PF (2015b) White-cheeked macaque (Macaca leucogenys): a new macaque species from Medog, southeastern Tibet. Am J Primatol 77:753–766. https://doi.org/10.1002/ajp.22394
Liedigk R, Yang M, Jablonski NG, Momberg F, Geissmann T, Lwin N et al (2012) Evolutionary history of the odd-nosed monkeys and the phylogenetic position of the newly described Myanmar snub-nosed monkey Rhinopithecus strykeri. PLoS ONE 7:e37418. https://doi.org/10.1371/journal.pone.0037418
Lin HB, Fan H, Zhang F, Huang XQ, Lin KQ, Shi L, Hu SN et al (2010) Genetic relationships of ethnic minorities in southwest China revealed by microsatellite markers. PLoS ONE 5:e9895. https://doi.org/10.1371/journal.pone.0009895
Liu J (2010) China’s road to sustainability. Science. https://doi.org/10.1126/science.1186234
Liu J (2013) Forest sustainability in China and implications for a telecoupled world. Asian Pac Policy Stud 1:230–250. https://doi.org/10.1002/app5.17
Liu L, Chen XC (2012) The archaeology of China: from the late paleolithic to the early bronze age. Cambridge University Press, Cambridge
Liu ZH, Yu SM, Yuan XC (1984) Resources of the Hainan black gibbons and its present situation. Chin Wildl 6:1–4
Liu XN, Feng ZM, Jiang LU, Li P, Liao CH, Yang YZ et al (2013) Rubber plantation and its relationship with topographical factors in the border region of China, Laos and Myanmar. J Geogr Sci 23:1019–1040. https://doi.org/10.1007/s11442-013-1060-4
Long YC, and Richardson M (2008) Rhinopithecus roxellana. The IUCN Red List of Threatened Species 2008: e.T19596A8985735
Long YC, Kirkpatrick CR, Zhong T, Xiao L (1994) Report on distribution, population and ecology of the Yunnan snub-nosed monkey (Rhinopithecus bieti). Primates 35:241–250. https://doi.org/10.1007/bf02382060
Long YC, Momberg F, Ma J, Wang Y, Luo YM, Li HS et al (2012) Rhinopithecus strykeri found in China! Am J Primatol 74:871–873. https://doi.org/10.1002/ajp.22041
Lu JQ, Hou JH, Wang HF, Qu WY (2007) Current status of Macaca mulatta in Taihangshan Mountains Area, Jiyuan, Henan, China. Int J Primatol 28:1085–1091. https://doi.org/10.1007/s10764-007-9197-6
Lu JQ, Tian JD, Zhang P (2018) Advances in ecological research regarding rhesus macaques (Macaca mulatta) in China. Acta Theriol Sin 37:74–84
Ma C, Huang ZP, Zhao XF, Zhang LX, Sun WM, Scott MB et al (2014) Distribution and conservation status of Rhinopithecus strykeri in China. Primates 55:1–6. https://doi.org/10.1007/s10329-014-0425-3
Ma C, Luo ZH, Liu CM, Orkin JD, Xiao W, Fan PF (2015) Population and Conservation Status of Indochinese Gray Langurs (Trachypithecus crepusculus) in the Wuliang Mountains, Jingdong, Yunnan, China. Int J Primatol 36:749–763. https://doi.org/10.1007/s10764-015-9852-2
Meng QY (2015) The land revolution in China from an academic history perspective: changes of topics and paradigms. Chin J Sociol 1:419–446. https://doi.org/10.1177/2057150x15593713
Meyer D, Momberg F, Matauschek C, Oswald P, Lwin N, Aung SS et al (2017) Conservation status of the Myanmar or black snub-nosed monkey Rhinopithecus strykeri. Fauna & Flora International, Yangon, Myanmar and German Primate Center, Göttingen
Ministry of Forestry (1987) China’s forest zoning. China’s Forestry Publishing House, Beijing
Nadler T (2016a) Indochinese Gray Langur—Trachypithecus crepusculus. In: Rowe N, Myers M (eds) All the World’s Primates. Pogonias Press, Charlestown, p 592
Nadler T (2016b) White-headed langur—Trachypithecus leucocephalus. In: Rowe N, Myers M (eds) All the world’s primates. Pogonias Press, Charlestown, pp 604–605
New T, Xie ZQ (2008) Impacts of large dams on riparian vegetation: applying global experience to the case of China’s Three Gorges Dam. Biodivers Conserv 17:3149–3163. https://doi.org/10.1007/s10531-008-9416-2
Newmark WD, Jenkins CN, Pimm SL, McNeally PB, Halley JM (2017) Targeted habitat restoration can reduce extinction rates in fragmented forests. Proc Natl Acad Sci USA 114:9635–9640. https://doi.org/10.1073/pnas.1705834114
Nijman V (2010) An overview of international wildlife trade from southeast Asia. Biodivers Conserv 19:1101–1114. https://doi.org/10.1007/s10531-009-9758-4
Nijman V, Shepherd CR, Nekaris KAI (2014) Trade in Bengal slow lorises in Mong La, Myanmar, on the China border. Primate Conserv 28:139–142. https://doi.org/10.1896/052.028.0112
Pan RL (1986) Environmental changes at Menghai and other cultivated areas in Xishangbanna, Yunnan Province. Zool Res 7:190–202 (in Chinese)
Pan RL, Oxnard C, Grueter CC, Li BG, Qi XG, He G et al (2016) A new conservation strategy for China: a model starting with primates. Am J Primatol 78:1137–1148. https://doi.org/10.1002/ajp.22577
Piao SL, Ciais P, Huang Y, Shen ZH, Peng SS, Li JS et al (2010) The impacts of climate change on water resources and agriculture in China. Nature 467:43–51. https://doi.org/10.1038/nature09364
Qiu J (2009) Where the rubber meets the garden. Nature 457:246–247. https://doi.org/10.1038/457246a
Qiu J (2010) China drought highlights future climate threats. Nature 465:142–143. https://doi.org/10.1038/465142a
Ren GP, Young SS, Wang L, Wang W, Long YC, Wu RD et al (2015) Effectiveness of China’s national forest protection program and nature reserves. Conserv Biol 29:1368–1377. https://doi.org/10.1111/cobi.12561
Rosen GE, Smith KF (2010) Summarizing the evidence on the international trade in illegal wildlife. EcoHealth 7:24–32. https://doi.org/10.1007/s10393-010-0317-y
Ruan GW, Huang ZY (2014) Preliminary study on population ecology of Macaca arctoides in Tiantang Mountain Nature Reserve, Yulin, Guangxi. J Yulin Norm Univ (Nat Sci) 35:64–67. https://doi.org/10.3969/j.issn.1004-4671.2014.02.016
Schwitzer C, Mittermeier RA, Rylands AB, Chiozza F, Williamson EA, Wallis J, Cotton A (2015) Primates in Peril: the world’s most endangered primates 2014–2016. Conservation International, Arlington
Struebig MJ, Fischer M, Gaveau DLA, Meijaard E, Wich SA, Gonner C et al (2015) Anticipated climate and land-cover changes reveal refuge areas for Borneo’s orang-utans. Glob Chang Biol 21:2891–2904. https://doi.org/10.1111/gcb.12814
Sun XF, Wang LQ, Gu ZB (2005) A brief overview of China’s timber market system. Int For Rev 6:221–226. https://doi.org/10.1505/ifor.6.3.221.59973
Sun BH, Li JH, Yong ZH, Xia DP (2010) Mitochondrial DNA variation in Tibetan macaques (Macaca thibetana). Folia Zool 59:301–307. https://doi.org/10.25225/fozo.v59.i4.a5.2010
Tang X, McLellan BC, Snowden S, Zhang BS, Höök M (2015) Dilemmas for China: energy, economy and environment. Sustainability 7:5508–5520. https://doi.org/10.3390/su7055508
Thang VN (2016) Eastern Black crested gibbon: Nomascus nasutus. In: Rowe N, Myers M (eds) All the World’s primates. Pogonias Press, Charlestown, pp 657–658
Townsend PA, Masters KL (2015) Lattice-work corridors for climate change: a conceptual framework for biodiversity conservation and social-ecological resilience in a tropical elevational gradient. Ecol Soc 20:1. https://doi.org/10.5751/es-07324-200201
Turvey ST, Grees JJ, Di Fonzo MMI (2015) Historical data as a baseline for conservation: reconstructing long-term faunal extinction dynamics in Late Imperial—modern China. Proc R Soc B. https://doi.org/10.1098/rspb.2015.1299
Turvey ST, Bryant JV, Duncan C, Wong MHG, Fei H, Ma C, Hong X, Nash HC, Chan BPL, Xu Y, Fan PF (2017) How many remnant gibbon populations are left on Hainan? Testing the use of local ecological knowledge to detect cryptic threatened primates. Am J Primatol 79:e22593. https://doi.org/10.1002/ajp.22593
UNEP-WCMC (2017) World database of protected areas, May 2017. www.protectedplanet.net
Vina A, McConell WJ, Yang HB, Xu ZC, Liu JG (2016) Effects of conservation policy on China’s forest recovery. Sci Adv 2:e1500965. https://doi.org/10.1126/sciadv.1500965
Viswanathan PK (2007) Critical issues facing China’s rubber industry in the era of market integration: an analysis in retrospect and prospect. Working Paper No. 177. Gujarat Institute of Development Research, Gota, India (ISBN 81-89023-35-7)
Waley-Cohen J (2004) Expansion and colonization in early modern Chinese history. Hist Compass 2:1–6. https://doi.org/10.1111/j.1478-0542.2004.00076.x
Wallis J, Lonsdorf EV (2010) Summary of recommendations for primate conservation education programs. Am J Primatol 72:441–444. https://doi.org/10.1002/ajp.20764
Wang WR, Yao M (2017) Fine-scale genetic structure analyses reveal dispersal patterns in a critically endangered primate, Trachypithecus leucocephalus. Am J Primatol 79:e22635. https://doi.org/10.1002/ajp.22635
Wang C, Lu HY, Zhang JP, Gu ZY, He KY (2014) Prehistoric demographic fluctuations in China inferred from radiocarbon data and their linkage with climate change over the past 50,000 years. Quat Sci Rev 98:45–59. https://doi.org/10.1016/j.quascirev.2014.05.015
Wang HH, Liu ZH, Xu ZM, Ding W, Xiao W (2015) Primate species diversity and disturbed way in Lanping. J Fujian For Sci 42:154–160
Wang J, Xin L, Tan M, Wang Y (2016a) Spatial heterogeneity in Chinese forest area change in the early 21st century. Forests 7:232. https://doi.org/10.3390/f7100232
Wang HH, Zhang SX, Xu ZM, Xiao W, Ding W, Xu LY et al (2016b) The temporal and spatial dynamics change of primate population in Yunlong County. Sichuan J Zool 35:801–809
Wang YY, Wu GL, Deng L, Tang ZS, Wang KB, Sun WY, Shangguan ZP (2017) Prediction of aboveground grassland biomass on the Loess Plateau, China, using a random forest algorithm. Sci Rep 7:6940. https://doi.org/10.1038/s41598-017-07197-6
Wei FW, Swaisgood R, Hu YB, Nie YG, Yan L, Zhang ZJ et al (2015) Progress in ecology and conservation of giant pandas. Conserv Biol 29:1497–1507. https://doi.org/10.1111/cobi.12582
Wen Y (2016) China’s rapid rise from backward agrarian society to industrial powerhouse in just 35 years. Reg Econ 24:8–14
Wu MC, Wei ZY, He NL (1987) Distribution and ecology of black langur (Presbytis francoisi Pousargues) in Guangxi Province. Chin Wildl 4:12–13 (in Chinese)
Wu RD, Long YC, Malanson GP, Garber PA, Zhang S, Li DQ et al (2014) Optimized spatial priorities for biodiversity conservation in China: a systematic conservation planning perspective. PLoS ONE 9:e103783. https://doi.org/10.1371/journal.pone.0103783
Xiang ZF, Huo S, Wang L, Cui LW, Xiao W, Quan RC, Tai Z (2007) Distribution, status and conservation of the black-and-white snub-nosed monkey Rhinopithecus bieti in Tibet. Oryx 41:525–531. https://doi.org/10.1017/s0030605307012124
Xu JC (2011) China’s new forests aren’t as green as they seem. Nature 477:371. https://doi.org/10.1038/477371a
Xu WH, Xiao Y, Zhang JG, Yang W, Zhang L, Hull V et al (2017) Strengthening protected areas for biodiversity and ecosystem services in China. Proc Natl Acad Sci USA 114:1601–1606. https://doi.org/10.1073/pnas.1620503114
Yang Y, Groves C, Garber PA, Wang X, Li H, Long Y, Li G, Tian Y, Dong S, Yang S, Behie A, Xiao W (2018) First insights into the feeding habits of the Critically Endangered Rhinopithecus strykeri: dietary characteristics and food resource distribution patterns. Am J Primatol
Ye ZZ (1993) Biology of Langurs (Presbytis). Yunnan Science and Technology Press, Kunming
Zhang YQ (2000) Deforestation and forest transition: theory and evidence in China. In: Palo M, Vanhanen H (eds) World forests from deforestation to transition?. Kluwer Academic Publishers, Dordrecht
Zhang YR, Wang SN (2018) Top ten conservation projects for Chinese wild animals—maintaining the beauty of the world. China Green Time (in Chinese). April 3
Zhang RZ, Chen LW, Qu WY, Chris C (2002) The primates of China: biogeography and conservation status. China Forestry Publishing House, Beijing
Zhang T, Teng JR, Wang HX (2005) Ecological habits and resource status of Tibetan macaques in Baishuijiang Nature Reserve of Gansu province. Sichuan J Zool 24:165–167
Zhang L, Hua N, Sun S (2008a) Wildlife trade, consumption and conservation awareness in southwest China. Biodivers Conserv 17:1493–1516. https://doi.org/10.1007/s10531-008-9358-8
Zhang Q, Lan SJ, Hu DM, Yu XX, Gu HJ, Chen X et al (2008b) Activities of Tibetan macaque in Baishuihe National Nature Reserve, Sichuan province. Sichuan J Zool 27:131–135
Zhang MX, Fellowes JR, Jiang XL, Wang W, Bosco PLC, Ren GP, Zhu JG (2010) Degradation of tropical forest in Hainan, China, 1991–2008: conservation implications for Hainan gibbon (Nomascus hainanus). Biol Conserv 143:1397–1404. https://doi.org/10.1016/j.biocon.2010.03.014
Zhao G (2006) Reinventing China imperial Qing ideology and the rise of modern Chinese National identity in the Early Twentieth Century. Mod China 32:3–30. https://doi.org/10.1177/0097700405282349
Zhao HT, Wang CL, Wang XW, Wang KF, Zhang BY, Li JF et al (2016) Discovery of Tibetan macaques (Macaca thibetana) in Shaanxi Micangshan National Nature Reserve. Chin J Zool 49:793–797. https://doi.org/10.13859/j.cjz.201406001
Zhao XM, Ren BP, Garber PA, Li XH, Li M (2018) Impacts of human activities and climate change on the distribution of snub-nosed monkeys in China during the past 2000 Years. Divers Distrib 24:92–102. https://doi.org/10.1111/ddi.12657
Zhou Y, Zhang P (2013) Distribution and vicissitude of gibbons (Hylobatidae) in China during the last 500 years. Acta Theriol Sin 33:258–266
Zhou J, Wei FW, Li M, Zhang JF, Wang DL, Pan RL (2005) Hainan Black-crested gibbon is headed for extinction. Int J Primatol 26:453–465. https://doi.org/10.1007/s10764-005-2933-x
Zhou XM, Wang BS, Pan Q, Zhang JB, Kumar S, Sun XQ et al (2014) Whole-genome sequencing of the snub-nosed monkey provides insights into folivory and evolutionary history. Nat Genet 46:1303–1310. https://doi.org/10.1038/ng.3137
Zhou XM, Meng XH, Liu ZZ, Chang J, Wang BS, Li MZ et al (2016) Population genomics reveals low genetic diversity and adaptation to hypoxia in snub-nosed monkeys. Mol Biol Evol 33:2670–2681. https://doi.org/10.1093/molbev/msw150
Zhu Q (2015) From the first five-year plan to the cultural revolution: the pre-reform urban transformation of Hangzhou. Plan Perspect 30:571–595. https://doi.org/10.1080/02665433.2014.995694
Ziegler AD, Fox JM, Xu JC (2009) The rubber juggernaut. Science 324:1024–1025. https://doi.org/10.1126/science.1173833
Acknowledgements
This project was sponsored by the following Grants: Baoguo Li, Key Project of Natural Science Foundation of China (31730104) and National Key Program of Research and Development, Ministry of Science and Technology (2016YFC0503200), Ming Li (National Key R&D Program of China, 2016YFC0503200 and Strategic Priority Research Program of the Chinese Academy of Sciences, XDPB0202), Pengfei Fan (National Program for Support of Top-notch Young Professionals in China, National Natural Science Foundation of China—NNSFC, 30900169; 31160424; 31372216). Jiqi Lu (NNSFC, 31472018 and 31672302) and Qingyong Ni (NNSFC, 31501873 and Kadoorie Conservation China, Kadoorie Farm & Botanic Garden Corporation). Jinhua Li (NSFC, No. 31672307 and 31372215). Sidney Gouveia (Brazilian CNPq Grants, Procs. #303180/2016-1 and 402469/2016-0); Zhipang Huang (Collaborative Innovation Center for Biodiversity and Conservation in the Three Parallel Rivers Region of China). Colin Groves unfortunately passed away during the final stage of this project, and we wish to highlight his contributions to this manuscript. We also wish to take this opportunity to celebrate his significant achievements in primatology, particularly those related to China through his cooperation with a great number of Chinese scholars. Ruliang Pan wishes to thank his wife, Aiqun Zhang, for her enthusiastic support, so that he was able to use spare time on the project. Paul A. Garber also wishes to thank Chrissie, Sara, and Jenni for their love, guidance, and support, which have inspired his work on primate conservation. Our final thanks is given to Rong Hou who assisted in manuscript preparation.
Author information
Authors and Affiliations
Corresponding authors
Additional information
Communicated by Xiaoli Shen.
Rights and permissions
About this article

Cite this article
Li, B., Li, M., Li, J. et al. The primate extinction crisis in China: immediate challenges and a way forward. Biodivers Conserv 27, 3301–3327 (2018). https://doi.org/10.1007/s10531-018-1614-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-018-1614-y