
Abstract
Despite the global trend in urbanization, little is known about patterns of biodiversity or provisioning of ecosystem services in urban areas. Bee communities and the pollination services they provide are important in cities, both for small-scale urban agriculture and native gardens. To better understand this important ecological issue, we examined bee communities, their response to novel floral resources, and their potential to provide pollination services in 25 neighborhoods across Chicago, IL (USA). In these neighborhoods, we evaluated how local floral resources, socioeconomic factors, and surrounding land cover affected abundance, richness, and community composition of bees active in summer. We also quantified species-specific body pollen loads and visitation frequencies to potted flowering purple coneflower plants (Echinacea purpurea) to estimate potential pollination services in each neighborhood. We documented 37 bee species and 79 flowering plant genera across all neighborhoods, with 8 bee species and 14 flowering plant genera observed on average along each neighborhood block. We found that both bee abundance and richness increased in neighborhoods with higher human population density, as did visitation to purple coneflower flower heads. In more densely populated neighborhoods, bee communities shifted to a suite of species that carry more pollen and are more active pollinators in this system, including the European honey bee (Apis mellifera) and native species such as Agapostemon virescens. More densely populated neighborhoods also had a greater diversity of flowering plants, suggesting that the positive relationship between people and bees was mediated by the effect of people on floral resources. Other environmental variables that were important for bee communities included the amount of grass/herbaceous cover and solar radiation in the surrounding area. Our results indicate that bee communities and pollination services can be maintained in dense urban neighborhoods with single-family and multi-family homes, as long as those neighborhoods contain diverse and abundant floral resources.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Currently, over 50 % of the world’s population lives in cities (United Nations Population Division 2006). The global trend toward urbanization threatens biodiversity and provisioning of ecosystem services (McKinney 2002; McDonald et al. 2013). Ecosystem services and biodiversity are often linked, with positive effects of biodiversity found for some regulating and supporting services (Balvanera et al. 2006). Biodiversity has been linked to pollination (Klein et al. 2003; Hoehn et al. 2008) but the relationship is not always straightforward (e.g., Albrecht et al. 2012). Pollination is an ecosystem service that is increasingly important in cities because urban farming efforts are increasing worldwide (McClintock 2010) and many crops benefit from insect pollination (Klein et al. 2007). Despite the potential for ecosystem services to benefit urban populations, it remains unclear to what degree biodiversity and pollination are altered or maintained across urbanized landscapes.
Pollination services are often linked to attributes of the bee community such as abundance, species richness, and composition (Kremen et al. 2002). Several studies have documented a diversity of bees in urban landscapes (McIntyre and Hostetler 2001; Matteson and Langellotto 2010), although composition may shift relative to nearby semi-natural landscapes. For example, abundance of soil-nesting bees may be reduced due to heavy management and soil compaction in urban landscapes (Cane 2005; but see Banaszak-Cibicka and Zmihorski 2012). Studies of urban bees have also indicated that diversity may be maintained, to some degree, by ornamental flowers in gardens and other managed habitats (Frankie et al. 2005; Matteson and Langellotto 2010). Residential gardens may be particularly important for urban biodiversity; in some cities, domestic gardens cover up to 25 % of the land and comprise up to 46 % of the total green space (Loram et al. 2007). A recent study in Chicago, IL identified the importance of green space, excluding turfgrass, for enhancing bee richness and abundance (Tonietto et al. 2011). However, the structure, composition, and cover of urban gardens and green spaces varies greatly across a city with factors such as human population density and income (Cadenasso et al. 2007), which can have important effects on bee communities as well as other organisms (e.g., birds and plants, Kinzig et al. 2005). Therefore, socioeconomic factors can provide additional insight into urban ecological patterns and dynamics (Grove and Burch 1997).
Many studies have found that floral visitation and pollination services vary with landscape context (Swift et al. 2004; Lonsdorf et al. 2009), often increasing with proximity to natural landscapes (Ricketts et al. 2008; Winfree et al. 2008). Proximity of domestic gardens enhanced bee abundance, richness, and seed set of focal plants in agricultural landscapes (Samnegard et al. 2011). These relationships have been studied extensively in agricultural areas but much less in urban areas. Cities are similar to agricultural systems in having high levels of external disturbance and a complex set of factors that influence bees. However, landscape heterogeneity occurs at a much finer scale in urban landscapes due to the mosaic of buildings, streets, and green spaces (Grimm et al. 2008). The few studies that evaluated the effects of urbanization on pollination services have found contrasting results. Pauw (2007) and Cheptou and Avendaño (2006) found pollination or seed set to be greatly reduced in urbanized landscapes compared with rural populations. On the other hand, Ksiazek et al. (2012) found no evidence of pollen limitation in nine native plants. These differences may be related to the amount of other floral resources in the surrounding landscape. In many cases, plant diversity and abundance has been shown to enhance pollinator diversity and facilitate visitation to flowering plants (Moeller 2004; Ghazoul 2006). However, in other cases, heterospecific plants compete for pollinators, resulting in decreased visits or pollination of plants as heterospecific diversity or abundance increases (Hennig and Ghazoul 2011). The occurrence of facilitation or competition between heterospecific plants may be related to the presence of exotic species (Morales and Traveset 2009) and may be species- (Hegland et al. 2009) or landscape-specific (Lázaro and Totland 2010). This is still an active area of research and is particularly understudied in urban settings, where exotic ornamental plants are abundant (McKinney 2004) and pollinators may be especially limited.
In this study, we examined pollinator communities and visits to summer blooming plants across Chicago, IL (USA). We examined the importance of local floral resources, socioeconomic factors, and surrounding land cover on abundance, richness, and composition of bee communities. We used Purple Coneflower (Echinacea purpurea) as a phytometer (plants used to estimate pollination; Samnegard et al. 2011; Albrecht et al. 2007) to examine how floral visitation, an estimate of pollination services (Vazquez et al. 2005), was impacted by nearby floral resources. Finally, we estimated pollinator performance for bee species in our system and used this value to understand how shifts in bee community composition might influence provision of pollination services. Our results suggest intriguing and encouraging trends for the maintenance of urban biodiversity and agriculture.
Methods
Study location and sample design
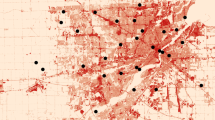
This study took place in Chicago, Illinois, the third largest city in the United States by population. We sampled 25 locations across the city, stratified by human population density (Fig. 1). Study locations included residential, industrial, and commercial areas. To maintain spatial independence, the minimum distance between nearest sites was 1.2 km (mean = 2.6 km), which is beyond the foraging range for most bees (Greenleaf et al. 2007) and a standard minimum distance in pollinator studies (e.g., Winfree et al. 2008). At each site, we evaluated the bee and floral community along a 150 × 6 m transect that followed a sidewalk. The sidewalk was surrounded on both sides by soil, grass, or other vegetation; a strip of public land (the “parkway”) separated the sidewalk from the road and the other side of the sidewalk was privately-owned land.

Map of study area with sample locations shown as black circles. Size of symbol varies according to the total neighborhood bee abundance. Small polygons are block groups as assigned by the U.S. Census Bureau; lighter shades indicate lower human population density
Our sampling was designed to observe pollinators that visit plants blooming during the summer months. We visited each study site three times between 7 July and 22 August, 2011, with ~2 weeks between each visit. Sampling was conducted only on calm, warm sunny days (21–27 °C). The order in which sites were visited was varied so that each site received roughly equivalent morning, noon, and afternoon sampling. At each location, we observed bee visits to Echinacea plants and extensively sampled bee communities and neighborhood floral resources.
Sampling neighborhood bee communities
We used a modified “Pollard walk” (Pollard 1977) to document the bee community at each site, employing hand nets to capture insects within 3 m of the transect during a 10-min sampling period. To avoid biasing other parts of our study, the Pollard walk took place after the floral visitation observations described below. We placed all bees netted on a single plant species in glass vials recently charged with ethyl acetate. Bees were identified to species by J.S. Ascher and were subsequently classified as native or non-native according to (Cane 2003).
For all bee species for which we collected more than two samples, we estimated the number of pollen grains on each bee’s body by viewing them under a 10 × power microscope. Pollen grains were not differentiated by plant species. The number of pollen grains was estimated separately for different body parts (head, thorax, abdomen, legs, and wings) and then summed. We scored the total number of pollen grains from 1 to 6, with an increase of 1 point for every 100 grains of pollen on the bee. We then calculated the average number of pollen grains found on each species, which we refer to as the species’ “pollen score”, and which served as a rough measure of pollination potential. The likelihood of a bee transferring pollen to another plant depends on the preparation and location of the pollen (e.g., whether or not it is moistened and packed onto corbicula, or added to other structures that might inhibit transfer), the size and behavior of the bee, and the reproductive biology of the receptive flower. For the purposes of this study, we generally assumed that bees that carry more total pollen would have a greater probability of pollen transfer. Upon capture, bees were quickly incapacitated but it is possible that some pollen was transferred among individuals at this stage.
Measuring environmental predictor variables
On each transect visit, we recorded all flowering plants and counted all associated inflorescences. We identified most plants in the field and took digital images of the remaining plants so that we could identify them later with a field guide; plant identifications were standardized at the genus level for analyses. We also used a number of spatial datasets to measure six land cover and socioeconomic variables around each sample location (Table 1). Data for human population density and median household income were gathered from the U.S. Census Bureau American Community Census (2006–2010) at the spatial resolution of census block groups. The percentage of grass/herbaceous cover, canopy cover, and impervious surface around each site were calculated using a classified high resolution (sub-meter) QuickBird satellite image and the Tabulate Area tool in ArcGIS 9.3. We calculated solar radiation around each site with the “Area Solar Radiation” tool in ArcGIS 10, adding a spatial data layer of buildings (and their heights) to ground elevation to use as the elevation surface. We initially measured each land cover and socioeconomic variable in 100 m buffer increments around each site from 100 to 1,500 m. We then analyzed Spearman’s rho correlations between each response variable and the environmental predictor variables at each scale. These analyses indicated a high degree of similarity in rho values among spatial scales, with most predictor variables showing a slight decrease in explanatory power with increasing spatial scale. Therefore, we limited analyses of our predictor variables to the 100 m buffer.
Assessing bee visitation to potted Echinacea plants
As a standardized estimate of pollination services, we placed and monitored a small group of potted flowering plants on the parkway at the center of each transect. While floral visitation is not a perfect measure of pollination, it has been linked to pollination in a number of studies (reviewed in Vazquez et al. 2005) and is frequently used as a surrogate for pollination (e.g., Ricketts et al. 2008; Winfree et al. 2008). We used Echinacea purpurea ‘Magnus’ (Echinacea hereafter), because the genus Echinacea is highly attractive to many bees (Wagenius and Lyon 2010). Mature Echinacea plants were purchased from a nursery and transplanted to two gallon pots in the greenhouse. The floral display was standardized so there were 20–30 flower heads at each location, with at least ten having pollen on the floral disc. We monitored insect visitors to the flowers for 60 min during each visit, capturing any insects that landed on the Echinacea flower heads during that time for later identification. The 60-min observation period provided a snapshot of pollinator activity and enabled us to visit multiple locations in a single day and minimize variation due to weather.
Statistical analyses
We had several goals in this study. First, we wanted to understand the factors that influence bee abundance, species richness, and community composition across the urban landscape. Second, by examining floral visitation rates to Echinacea plants, and by estimating pollen load and visitation frequency of each bee species, we aimed to gain insight into the factors that influence pollination services in the city. Because data from the three sampling dates were highly correlated, we pooled the data to determine abundance (summed over all visits) and species richness and composition (calculated by including any species that was present during any visit) at each site. For our analyses, we used several different subsets of data to describe the bee community and potential pollination services at each sample location: (1) all bees captured at a sample location, including bees visiting the Echinacea and bees observed on the transect, called “neighborhood bee abundance,” “neighborhood bee richness,” or “neighborhood community composition” hereafter, (2) bees that visited the potted Echinacea plants, called “Echinacea visitors” or “Echinacea bee richness” hereafter, and (3) bees that were captured along the transect—not visiting the Echinacea plants—after the 60-min Echinacea observation period (“transect bee abundance” or “transect bee richness”).
We first used scatter plots and Spearman’s correlations to examine the relationships between neighborhood bee abundance and richness and each environmental variable. We then used distance-based linear modeling (DistLM) to identify the most parsimonious models for neighborhood bee abundance, neighborhood bee richness, and neighborhood community composition. DistLM is similar to multiple linear regression but uses permutation to avoid the assumption of normality and can test the significance of explanatory variables for univariate or multivariate response variables (Anderson et al. 2008). We used a model selection approach to identify the best model among all possible subsets of predictor variables, with AICc as a selection criterion. We examined all top models with ΔAIC < 2. We calculated Akaike weights for each model and used model-averaging to determine the importance of each predictor variable (Burnham and Anderson 1998). Before DistLM analysis, response variables were square-root transformed. We used PERMANOVA + PRIMER v 6.1.13 (Anderson et al. 2008) to create the distance-based linear models.
To further explore the patterns in neighborhood bee community composition across our study area, we used non-metric multidimensional scaling (NMS) to ordinate bee species based on their patterns of association. Because NMS is an indirect ordination technique, environmental variables are not directly used in the ordination but can be overlaid on the plot to visually assess their relationship with each species. We removed all species present at only one location (as recommended by McCune and Grace 2002) and used Sorensen’s (Bray–Curtis) similarity index to calculate species dissimilarity between all pairs of samples. We performed a step-down procedure to determine the best number of axes (i.e., dimensionality) for the dataset, which are then used to create the final ordination from a random starting configuration with 500 iterations. NMS analyses were conducted with PC-ORD 6.0 (McCune and Mefford 2011).
To evaluate how neighborhood floral resources and bee communities affected potential pollination services, we used Spearman’s correlations to assess the influence of transect bee abundance, transect bee richness, total floral inflorescences, and floral genera richness on Echinacea bee visitation and richness. As a post hoc analysis, after we identified a strong positive relationship between human population density and floral resources, we also examined the relationship between human population density and total bee visits to Echinacea flower heads.
Finally, we created an index of pollinator performance so we could examine how shifts in community composition might affect pollination services. This index included both pollinator quantity and quality—two aspects of pollination performance that are important but independent of each other (Zych et al. 2013). The pollen score provided an indirect measure of pollinator quality for each species. As all bees were visiting flowers when they were observed, the abundance of each species provided a measure of pollinator visitation or quantity. To calculate pollinator performance for species s, we multiplied that species’ pollen score, the scalar metric for number of pollen grains on the body, by its total abundance, as shown below.
Results
Neighborhood bee communities
A total of 515 bees and 37 species (See Table 3 in Appendix) were observed at the 25 sample locations across Chicago. Nine species were non-native. On average, we observed 20.6 bees and 8.4 species at each sample location. Over all transects, bee species richness was highly correlated with mean bee abundance (rho = 0.88, p < 0.001). The most commonly encountered bees were Agapostemon virescens (15 sites, 11 % of all bees), Apis mellifera (14 sites, 13 % of all bees), Hylaeus leptocephalus (12 sites, 6 % of all bees), and Bombus impatiens (11 sites, 4 % of all bees). Five bee species were only present at a single site. The pollen score for each species ranged from 1.0 to 6.0 (See Table 3 in Appendix). The highest pollen scores were seen in the Apidae family (especially bumble bees and carpenter bees), while the lowest scores were seen in Hylaeus species.
Environmental predictor variables
There were 79 flowering plant genera observed over all transects. Flowers were present at all sites including areas with >85 % impervious surface. On average, each transect had 2539 flower inflorescences and 14 genera of flowering plants. The most common flowering plants in terms of number of transects were White Clover (Trifolium repens, present at 21 sites), dandelion agg. (Taraxacum spp., present at 20 sites), Common and Narrowleaf Plantain (Plantago spp., present at 18 sites), and Poor-man’s Pepper (Lepidium virginicum, present at 17 sites).
Impervious surface was the dominant land cover around sample sites, averaging 64 % of total land cover within 100 m of each transect. Because amount of impervious surface was not independent of other cover types, it was not used in any of our models. Grass/herbaceous cover and tree canopy comprised most of the remaining area, covering an average of 18 and 17 % of total land around each transect, respectively (Table 1). The following pairs of environmental variables had significant Spearman correlation coefficients: human population density and solar radiation (rho = −0.56, p = 0.004), human population density and floral richness (rho = 0.59, p = 0.002), and human population density and tree canopy cover (rho = 0.44, p = 0.03). The variance inflation factor for all variables was <2.3, indicating that multicollinearity was not a concern for our models.
Relationships between environmental variables and neighborhood bee communities
A number of environmental variables were significantly correlated with neighborhood bee abundance, including abundance of floral inflorescences on each transect (rho = 0.41, p = 0.04), solar radiation (rho = −0.39, p = 0.05), and human population density (rho = 0.46, p = 0.02) (Fig. 2). Neighborhood bee species richness was positively correlated with human population density and floral richness (rho = 0.41, p = 0.04; rho = 0.47, p = 0.02, respectively) but negatively correlated with solar radiation (rho = −0.53, p = 0.006). When examined separately, native bee species richness increased moderately with human population density (rho = 0.39, p = 0.06) but exotic bee species richness was unrelated to human population density (rho = 0.25, p = 0.23).

Relationship between human population density, floral resources, and the bee community in urban neighborhoods
The best DistLM model for neighborhood bee abundance explained 40 % of the variability, and included positive effects of human population density and floral abundance (Table 2). Human population density was the most important predictor variable (Fig. 3). The best model for neighborhood bee species richness explained 37 % of the variability and included a negative effect of solar radiation and a positive effect of floral richness. Solar radiation was the most important predictor variable (Fig. 3). The top model for neighborhood bee community composition explained 22 % of the variability and included grass/herbaceous cover and solar radiation. Grass/herbaceous cover was the most important predictor variable for community composition (Fig. 3).

Variable importance as determined by model averaging. For each of the top models (ΔAICc < 2) in which a variable appeared, the model Akaike weight was multiplied by the proportion of variation explained by that variable in a marginal test. This product was summed over all models to calculate variable importance
For the NMS ordination, the step-down procedure suggested a 3-dimensional solution with a stress of 14.4. The three axes combined explained 76 % of the variation in the species composition, with axes 1 and 2 explaining the most (32 and 31 %, respectively). Environmental variables were overlaid on the ordination plot to assess their correlation with each axis and variables with r > 0.3 are shown (Fig. 4). Axis 1 was positively correlated with grass/herbaceous cover (r = 0.43) and solar radiation (r = 0.33) while axis 2 was negatively correlated with human population density (r = −0.50) and positively correlated with solar radiation (r = 0.33). The NMS results correspond roughly to the results of the distance-based linear model, which showed that solar radiation and grass/herbaceous cover were important predictors of community composition. Hylaeus and Megachile species tended to be strongly and negatively associated with axis 1 (and thus negatively associated with grass/herbaceous cover), while most Lasioglossum species were positively associated with axis 1. Few species were positively associated with axis 2 (except Megachile concinna and Bombus impatiens) but most Hylaeus species and Lasioglossum species were strongly and negatively associated with axis 2 (and thus positively associated with human population density), along with Apis mellifera, Agapostemon virescens, Anthidium oblongatum, Bombus griseocollis, and Melissodes subillata.

NMS ordination of the bee community, with related predictor variables (r > 0.3) overlaid. Axis 1 is positively correlated with grass/herbaceous cover (r = 0.43) and solar radiation (r = 0.33) while axis 2 is negatively correlated with human population density (r = −0.50) and positively correlated with solar radiation (r = 0.33)
Bee visitation to potted Echinacea plants
On average, 7.8 bees (range 0–31) and 4 species (range = 0–11) visited the Echinacea plants at each site. After our 60-min Echinacea observations, we recorded an average of 12.8 bees (range = 1–34) and 6 species (range = 1 = 16) along each transect. Number of Echinacea visitors was significantly correlated with transect bee abundance (rho = 0.54, p = 0.005) and Echinacea bee richness was significantly correlated with transect bee richness (rho = 0.58, p = 0.002). Number of Echinacea visitors was not significantly related to abundance of floral inflorescences (rho = 0.24, p = 0.24) or plant genera richness (rho = 0.31, p = 0.13) on a transect, nor was Echinacea bee richness significantly related to abundance of floral inflorescences (rho = 0.10, p = 0.62) or plant genera richness (rho = 0.33, p = 0.11) (Fig. 2). Human population density was positively correlated with number of Echinacea visitors (rho = 0.54, p = 0.005) and Echinacea bee richness (rho = 0.50, p = 0.01).
Estimating pollinator performance for bee species
The pollinator performance index for different bee species ranged from 3 to 272 (See Table 3 in Appendix). There was a significant, positive relationship between pollen score and our measure of pollinator performance (rho = 0.52, p = 0.006), indicating that species that carry more pollen also tended to be ranked higher for pollinator performance. However, this relationship was not consistent, as some species with high pollen scores were uncommon flower visitors at our sample locations. The five mostly highly-ranked species for pollinator performance, and the only species with a measure of pollinator performance greater than 100, were (in order from highest to lowest) Apis mellifera (pollinator performance = 272), Agapostemon virescens (128), Bombus impatiens (127), Halictus ligatus (104), and Melissodes bimaculata (101).
Discussion
We found that floral diversity, bee abundance and richness, and bee visitation to potted Echinacea plants all increased significantly with human population density. Our results contrast somewhat with a similar study in New York City (NYC), in which pollinator richness decreased with increasing human population density (Matteson et al. 2013). In NYC, this relationship was mediated by a direct, negative relationship between people and floral resources that was most apparent at very high population densities characterized by high rise apartment and office buildings. In Chicago, we did not sample the most densely-populated areas (i.e., city center), as we could not set our potted plants down in those areas. Therefore, the maximum human population density in this study was much less than the NYC study (147 and 870 people/ha respectively).
Similar to the NYC study (Matteson et al. 2013), the link between humans and bees in Chicago is likely indirect and mediated partly by flowering plants, which are often planted and managed by people. In our study, neighborhoods with higher human population density did not have higher abundance of flowers but they did have a higher richness of flowering plant genera. Furthermore, densely-populated neighborhoods in our study area had more tree canopy and no increase in impervious surface (results not shown). When examined together, these results suggest that some densely populated urban neighborhoods are relatively good places to be an urban bee, as they can have a greater variety of floral resources, potentially more cavity-nesting sites, and no more impervious surface than less-densely populated neighborhoods.
We explain this trend with several observations. Among our study sites, population density increased across the following land uses: commercial/industrial, single-family homes, multi-family homes (e.g., townhouses or 3- or 4-flats). Commercial and industrial neighborhoods often had a lot of impervious surface and few gardens while neighborhoods with single-family homes tended to have relatively large lawns and only small flower beds (if any) near the house. In contrast, while neighborhoods with multi-family homes typically had small yards, available lawn area was often replaced by flower beds or pot gardens. We speculate that the increase in flowering plant genera with moderately high human population density may arise from people’s diverse preferences for garden plants (Kendal et al. 2012) and a desire to maximize floral plantings in available space. This pattern adds more diverse resources beyond common but less attractive flowers such as dandelions. Previous studies have linked richness of urban bee communities to ornamental flowers in private yards (Frankie et al. 2005) and suggested that increased availability of species-rich flower patches could improve the spatial extent of pollination services in urban areas (Jha and Kremen 2013). The relationship between people and bees may be maximized in densely populated urban neighborhoods with diverse, concentrated floral plantings.
Bee visitation to the potted Echinacea flower heads also increased with human population density. However, bee visitation does not always correspond to pollination services or plant fitness (Leong et al. 2014). On one hand, previous work has shown that plant-pollinator interactions in urban areas tend to be dominated by generalists (Jędrzejewska-Szmek and Zych 2013), and that is also true for bees in our study area (Tonietto et al. 2011). Generalist bees may be inferior pollinators in some systems (Larson 2005). On the other hand, a meta-analysis of the effect of animal mutualists on plants found that frequent animal mutualists tend to contribute the most to pollination or seed dispersal, even if they are not very effective at the per-interaction level (Vazquez et al. 2005).
Bee community composition also shifted across our study sites, which could influence pollination services if species varied in their performance as pollinators (Winfree and Kremen 2009). Our measure of pollinator performance, based on pollen load and visitation frequency, identified several species considered to be effective pollinators in other systems (e.g. Apis mellifera, Bombus spp. Megachile spp. Agapostemon virescens; Artz and Nault 2011; Wagenius and Lyon 2010). Our measure was indirect and not intended to be specific to any particular plant but identified species that carry a lot of pollen and are abundant in this setting. Several of the bees we identified as potentially effective pollinators were positively associated with Axis 2 of the ordination and increased in abundance with human population density. Therefore, we have two lines of indirect evidence suggesting that pollination may increase in densely-populated neighborhoods: increased visitation to potted plants, and an increase in several bee species that are likely effective pollinators. Our future work will examine this relationship more directly through seed and fruit set of potted plants.
One potential explanation for the observed shift in bee community composition with increasing population density involves the availability of nesting sites. For example, Hylaeus and Megachile, which are predominantly cavity-nesters, tended to be positively associated with human population density, while Lasioglossum, which are soil-nesters, tended to be positively associated with grass/herbaceous cover. However, there were exceptions to this trend and the degree to which bee nesting sites are limited in urban landscapes remains unclear. Some bee species have apparently adapted to urban conditions, nesting in non-traditional settings like artificial structures (Owen 1991) and flowerpots (Matteson et al. 2008). Other life history traits may also play a role in the shifting community composition. Traits such as smaller body size and later emergence date may increase a bee’s success at colonizing the urban core (Banaszak-Cibicka and Zmihorski 2012). Finally, bees with wider foraging ranges and more complex social structures, such as Bombus and Apis, may be more adept at locating and exploiting floral resources in densely populated neighborhoods.
In addition to human population density and floral resources, other important explanatory variables in our models include grass/herbaceous cover and solar radiation. Grass/herbaceous cover provide nesting (Michener 2006) and floral resources for bees. Solar radiation has been shown to have a positive influence on bees (Vicens and Bosch 2000; Matteson and Langellotto 2010) but, in our study, was negatively correlated with species richness, perhaps due to its negative correlation with human population density. It may be that any negative influence of shading on bees was not enough to overwhelm the positive effect of increased floral resources in densely populated neighborhoods. Alternatively, some level of shading may reduce overheating of insects and plants and prolong the period of nectar availability, thus altering bee foraging patterns (Willmer 1983).
Many studies conducted along rural–urban gradients show that diversity of pollinating insects decreases as urbanization or human disturbance increases (Ahrné et al. 2009; Bates et al. 2011). Our study does not contradict these studies but instead offers a nuanced perspective on the effects of urbanization on bee communities. Inside city limits, human population density does not necessarily equate to reduced resources for bees. All of our sample locations—whether low or high in human population density—occupy what is usually considered the “urban core” of rural–urban gradients, defined by McKinney (2008) as “hardscapes” with more than 50 % impervious surface. Instead of encompassing a gradient, our sample locations reveal differences across urban land use types. As McKinney (2008) states, “the complex nature of urban land use can have a complicated influence on local biodiversity.” Our data support this statement and suggest that broad generalizations about urban land should be avoided. However, our data do suggest a synergistic relationship between humans, bees, and pollination services in moderate to high (but not extreme) population densities. Although a few studies suggest that pollinator species richness may benefit from moderate levels of development (Carper et al. 2014; Winfree et al. 2007), this is the first study to propose that human population density within the urban core acts in a positive fashion for pollination services.
It is important to point out that our results are based on data collected during the summer months, and conclusions may differ if spring-flying bees had been included. However, despite this limitation, our study provides an optimistic perspective on the sustainability of urban biodiversity and agriculture. Similar to Wojcik and McBride (2012), our results show that the surrounding landscape influences community composition but local floral resources are also an important driver of urban pollinator assemblages. Although not all ornamental flowers provide nectar or pollen, diverse floral plantings in urban neighborhoods should help mitigate the otherwise negative impacts of development.
References
Ahrné K, Bengtsson J, Elmqvist T (2009) Bumble bees (Bombus spp.) along a gradient of increasing urbanization. PLoS ONE 4:e5574
Albrecht M, Duelli P, Muller C, Kleijn D, Schmid B (2007) The Swiss agri-environment scheme enhances pollinator diversity and plant reproductive success in nearby intensively managed farmland. J Appl Ecol 44:813–822
Albrecht M, Schmid B, Hautier Y, Muller CB (2012) Diverse pollinator communities enhance plant reproductive success. Proc R Soc B 279(4845):4852
Anderson MJ, Gorley RN, Clarke KR (2008) PERMANOVA + for PRIMER: guide to software and statistical methods. Primer E, Plymouth
Artz DR, Nault BA (2011) Performance of Apis mellifera, Bombus impatiens, and Peponapis pruinosa (Hymenoptera:Apidae) as pollinators of pumpkin. J Econ Entomol 104:1153–1161
Balvanera P, Pfisterer AB, Buchmann N, He J-S, Nakashizuka T, Raffaelli D, Schmid B (2006) Quantifying the evidence for biodiversity effects on ecosystem functioning and services. Ecol Lett 9:1146–1156
Banaszak-Cibicka W, Zmihorski M (2012) Wild bees along an urban gradient: winners and losers. J Insect Conserv 16:331–343
Bates AJ, Sadler JP, Fairbrass AJ, Falk SJ, Hale JD, Matthews TJ (2011) Changing bee and hoverfly pollinator assemblages along an urban-rural gradient. PLoS ONE 8:e23459
Burnham KP, Anderson DR (1998) Model selection and inference: a practical information-theoretic approach. Springer, New York
Cadenasso ML, Pickett STA, Schwarz K (2007) Spatial heterogeneity in urban ecosystems: reconceptualizing land cover and a framework for classification. Front Ecol Environ 5:80–88
Cane JH (2003) Exotic nonsocial bees in North American: ecological implications. In: Strickler K, Cane JH (eds) For nonnative crops, whence pollination of the future. Thomas Say Foundation. Entomological Society of America, Lanham, pp 113–126
Cane JH (2005) Bees, pollination, and the challenges of sprawl. In: Johnson EA, Klemens MW (eds) Nature in fragments: the legacy of sprawl. Columbia University Press, New York, pp 109–124
Carper AL, Adler LS, Warren PS, Irwin RE (2014) Effects of suburbanization on forest bee communities. Environ Entomol 43:253–262
Cheptou PO, Avendaño VLG (2006) Pollination processes and the Allee effect in highly fragmented populations: consequences for the mating system in urban environments. New Phytol 172:774–783
Frankie GW, Thorp RW, Schindler M, Hernandez J, Ertter B, Rizzardi M (2005) Ecological patterns of bees and their host ornamental flowers in two northern California cities. J Kansas Entomol Soc 78:227–246
Ghazoul J (2006) Floral diversity and the facilitation of pollination. J Ecol 94:295–304
Greenleaf SS, Williams NM, Winfree R, Kremen C (2007) Bee foraging ranges and their relationship to body size. Oecologia 153:589–596
Grimm NB, Faeth SH, Golubiewski NE, Redman CL, Wu J, Bai X, Briggs JM (2008) Global change and the ecology of cities. Science 319:756–760
Grove JM, Burch WR Jr (1997) A social ecology approach and applications of urban ecosystem and landscape analyses: a case study of Baltimore Maryland. Urban Ecosyst 1:259–275
Hegland SJ, Grytnes J-A, Totland Ø (2009) The relative importance of positive and negative interactions for pollinator attraction in a plant community. Ecol Res 24:929–936
Hennig EI, Ghazoul J (2011) Plant pollinator interactions within the urban environment. Perspect Plant Eco Evol Syst 13:137–150
Hoehn P, Tscharntke T, Tylianakis JM, Steffan-Dewenter I (2008) Functional group diversity of bee pollinators increases crop yield. Proc R Soc B 275:2283–2291
Jędrzejewska-Szmek K, Zych M (2013) Flower-visitor and pollen transport networks in a large city: structure and properties. Arthropod-Plant Interact 7:503–516
Jha S, Kremen C (2013) Resource diversity and landscape-level homogeneity drive native bee foraging. Proc Natl Acad Sci 110:555–558
Kendal D, Williams KJH, Williams NSG (2012) Plant traits link people’s plant preferences to the composition of their gardens. Landsc Urban Plan 105:34–42
Kinzig AP, Warren P, Martin C, Hope D, Katti M (2005) The effects of human socioeconomic status and cultural characteristics on urban patterns of biodiversity. Ecol Soc 10:23
Klein AM, Steffan-Dewenter I, Tscharntke T (2003) Fruit set of highland coffee increases with the diversity of pollinating bees. Proc R Soc B 270:955–961
Klein AM, Vaissiere BE, Cane JH, Steffan-Dewenter I, Cunningham SA, Kremen C, Tscharntke T (2007) Importance of pollinators in changing landscapes for world crops. Proc R Soc B 274:303–313
Kremen C, Williams NM, Thorp RW (2002) Crop pollination from native bees at risk from agricultural intensification. Proc Natl Acad Sci 99:16812–16816
Ksiazek K, Fant J, Skogen K (2012) An assessment of pollen limitation on Chicago green roofs. Landsc and Urban Plan 107:401–408
Larson M (2005) Higher pollinator effectiveness by specialist than generalist flower-visitors of unspecialized Knautia arvensis (Dipsacaceae). Oecologia 146:394–403
Lázaro A, Totland Ø (2010) Population dependence in the interactions with neighbors for pollination: a field experiment with Taraxacum officinale. Am J Bot 97:760–769
Leong M, Kremen C, Roderick GK (2014) Pollinator interactions with yellow starthistle (Centaurea solstitialis) across urban, agricultural, and natural landscapes. PLoS ONE 9:e86357
Lonsdorf E, Kremen C, Ricketts T, Winfree R, Williams N, Greenleaf S (2009) Modelling pollination services across agricultural landscapes. Ann Bot 103:1589–1600
Loram A, Tratalos J, Warren PH, Gaston KJ (2007) Urban domestic gardens (X): the extent & structure of the resource in five major cities. Landsc Ecol 22:601–615
Matteson KC, Langellotto GA (2010) Determinates of inner city butterfly and bee species richness. Urban Ecosyst 13:333–347
Matteson KC, Ascher JS, Langellotto GA (2008) Bee richness and abundance in New York City gardens. Ann Entomol Soc Am 101:140–150
Matteson KC, Grace JB, Minor ES (2013) Direct and indirect effects of land use on floral resources and flower-visiting insects across an urban landscape. Oikos 122:682–694
McClintock N (2010) Why farm the city? Theorizing urban agriculture through a lens of metabolic rift. Camb J Reg Econ Soc 3:191–207
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software Design, Gleneden Beach
McCune B, Mefford MJ (2011) PC-ORD. Multivariate analysis of ecological data. Version 6. MjM Software, Gleneden Beach
McDonald RI, Marcotullio PJ, Güneralp B (2013) Urbanization and global trends in biodiversity and ecosystem services. In: Elmqvist T, Fragkias M, Goodness J, Güneralp B, Marcotullio PJ, McDonald RI, Parnell S, Schewenius M, Sendstad M, Seto KC, Wilkinson C (eds) Urbanization, biodiversity and ecosystem services: challenges and opportunities. Springer, New York
McIntyre NE, Hostetler ME (2001) Effects of urban land use on pollinator communities in a desert metropolis. Basic Appl Ecol 2:209–218
McKinney ML (2002) Urbanization, biodiversity, and conservation. Bioscience 52:883–890
McKinney ML (2004) Measuring floristic homogenization by non-native plants in north America. Glob Ecol Biogeogr 13:47–53
McKinney ML (2008) Effects of urbanization on species richness: a review of plants and animals. Urban Ecosyst 11:161–176
Michener CD (2006) The bees of the World, 2nd edn. Johns Hopkins University Press, Baltimore
Moeller DA (2004) Facilitative interactions among plants via shared pollinators. Ecology 84:3289–3301
Morales CL, Traveset A (2009) A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecol Lett 12:716–728
Owen J (1991) The ecology of a garden, the first fifteen years. Cambridge University Press, Cambridge, p 225
Pauw A (2007) Collapse of a pollination web in small conservation areas. Ecology 88:1759–1769
Pollard E (1977) A method for assessing changes in the abundance of butterflies. Biol Conserv 12:115–134
Ricketts TH, Regetz J, Steffan-Dewenter I, Cunningham SA, Kremen C, Bogdanski A, Gemmill-Herren B, Greenleaf SS, Klein AM, Mayfield MM, Morandin LA, Ochieng A, Viana BF (2008) Landscape effects on crop pollination services: are there general patterns? Ecol Lett 11:499–515
Samnegard U, Persson AS, Smith HG (2011) Gardens benefit bees and enhance pollination in intensively managed farmland. Biol Conserv 144:2602–2606
Swift MJ, Izac AMN, van Noordwijk M (2004) Biodiversity and ecosystem services in agricultural landscapes–are we asking the right questions? Agric Ecosyst Environ 104:113–134
Tonietto R, Fant J, Ascher J, Ellis K, Larkin D (2011) A comparison of bee communities of Chicago green roofs, parks, and prairies. Landsc Urban Plan 103:102–108
United Nations Population Division (2006) World Urbanization Prospects: The 2005 Revision. United Nations, Department of Economic and Social Affairs, New York, New York, USA
Vazquez DP, Morris WF, Jordano P (2005) Interaction frequency as a surrogate for the total effect of animal mutualists on plants. Ecol Lett 8:1088–1094
Vicens N, Bosch J (2000) Weather-dependent pollinator activity in an apple orchard, with special reference to Osmia cornuta and Apis mellifera (Hymenoptera: Megachilidae and Apidae). Environ Entomol 29:413–420
Wagenius W, Lyon SP (2010) Reproduction of Echinacea angustifolia in fragmented prairie is pollen-limited but not pollinator-limited. Ecology 91:733–742
Willmer PG (1983) Thermal constraints on activity patterns in nectar-feeding insects. Ecol Entomol 8:455–469
Winfree R, Kremen C (2009) Are ecosystem services stabilized by differences among species? A test using crop pollination. Proc R Soc B 276:229–237
Winfree R, Griswold T, Kremen C (2007) Effects of human disturbance on bee communities in a forested ecosystem. Conserv Biol 21:213–223
Winfree R, Williams NM, Gaines H, Ascher JS, Kremen C (2008) Wild bee pollinators provide the majority of crop visitation across land-use gradients in New Jersey and Pennsylvania, USA. J Appl Ecol 45:793–802
Wojcik VA, McBride JR (2012) Common factors influence bee foraging in urban and wildland landscapes. Urban Ecosyst 15:581–598
Zych M, Goldstein J, Roguz K, Stpiczynska M (2013) The most effective pollinator revisited: pollen dynamics in a spring-flowering herb. Arthropod-Plant Interact 7:315–322
Acknowledgments
The authors thank the staff at UIC’s plant research laboratory for assisting with care for Echinacea and John Ascher for bee identification. Thanks to Amélie Davis for thoughtful comments on an earlier version of this manuscript and Eric Lonsdorf for help with the solar radiation calculations. We also thank two anonymous reviewers, whose suggestions improved the manuscript. This work was funded by NSF Proposal Number: DEB-1120376.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Jens Wolfgang Dauber.
Appendix
Appendix
See Table 3.
Rights and permissions
About this article

Cite this article
Lowenstein, D.M., Matteson, K.C., Xiao, I. et al. Humans, bees, and pollination services in the city: the case of Chicago, IL (USA). Biodivers Conserv 23, 2857–2874 (2014). https://doi.org/10.1007/s10531-014-0752-0
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-014-0752-0