
Abstract
Tropical botanical gardens have played an important role in the distribution, naturalisation and spread of non-native plants worldwide. Appropriate guidance relating to risk assessments of established botanical garden collections is often scarce. This paper uses the Amani Botanical Garden (ABG), Tanzania, as a case study to highlight appropriate methods to assess the risks posed by existing and future collections in tropical botanical gardens. Key considerations included field assessments of species status using accepted definitions of naturalisation, spread and invasion, distinguishing between intentionally and unintentionally introduced species, identifying taxonomic patterns in invasion status, assessing patterns in habitats colonised, and determining how knowledge of invasion elsewhere might be useful in forecasting risk. Out of the 214 alien plant species surviving from the original plantings in the early 20th century, 35 had only regenerated, 38 had locally naturalised while 16 had spread widely in the botanical garden. A further 16 species with unclear introduction records in the garden were also found to be naturalised. A greater proportion of introduced species were potentially invasive than might be expected from previous analyses of global floras. Overall, just over half of all naturalised and spreading species were also observed in forest fragments and edges. The proportion of species that had been recorded elsewhere as naturalised/invasive was significantly related to their status in ABG, with 94% of spreading species and 79% of naturalising species being recorded as naturalised or invasive elsewhere, compared to 57% of species that were only regenerating and 49% of species only surviving. Recommendations for further risk assessments of botanical garden collections are discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
A substantial body of evidence highlights the important role horticulture has played in the dissemination, naturalisation and invasion of alien plants worldwide (Reichard and White 2001; Weber 2003; Hulme 2005; Mack 2005; Dehnen-Schmutz et al. 2007). The increasing awareness among industry representatives to tackle this issue (Peters et al. 2006) has resulted in attempts to establish new codes of conduct within the horticultural trade (Baskin 2002; Reichard 2004). Botanical gardens play a unique role within the horticultural sector by contributing to the selection and propagation of new plant varieties (Dosman and Del Tredici 2003; Anderson et al. 2006), the conservation of endangered or economically valuable species, public education and recreation (Maunder 1994; Muller 1994; Maunder et al. 2001; Waylen 2006) and in many cases they are of historical as well as cultural significance. However, in contrast to the current philosophy of botanical gardens, it was the importance of trade and commerce as well as competition between colonial powers in the 18 and 19th centuries that drove the establishment of botanical gardens in the tropics (Heywood 1987). As a result, large numbers of tropical and subtropical plant species, including fruits, spices, legumes, as well as hardwoods and softwoods were transported across the tropical regions of the globe and ultimately, some became invasive in the subsequent decades and centuries (Cronk and Fuller 1995).
Although direct attribution is difficult, there are several cases where tropical botanical gardens have allegedly played a key role in the establishment of invasive plant species (Table 1). This risk was highlighted at the 2nd World Botanical Gardens Congress (Leadlay 2004) and resulted in a recommendation that all botanical gardens carry out invasive species risk assessments of their collections and management practices. Yet, appropriate guidance relating to risk assessments of established collections is scarce (Jefferson et al. 2004). In this paper, we use the Amani Botanical Garden (ABG) in the East Usambara Mountains, Tanzania, as a case study to highlight key considerations and appropriate methods to assess the risks posed by existing and future collections in tropical botanical gardens. Previous surveys of the ABG (Sheil 1994), and surrounding forests in the Usambara Mountains (Iversen 1991), have highlighted the relatively high number of naturalised alien plant species (48 and 239 respectively). Many of the species listed in both inventories were intentionally introduced via the botanical garden (Greenway 1934), but survey methods to construct these lists did not focus specifically on invasion risk from the ABG collections. The quantitative assessment of the risks posed by ABG presented here include:
-
(1)
Clearly differentiating among regenerating, naturalised and spreading species (sensu Richardson et al. 2000a; Pyšek et al. 2004);
-
(2)
Distinguishing between opportunistic species unintentionally introduced into the botanical garden from species establishing from the collections (Hulme et al. in press);
-
(3)
Assessing whether colonisation occurs exclusively in disturbed areas or also in neighbouring semi-natural ecosystems (Hulme 2003).
Consistent data collation procedures should assist in identifying whether the risks of invasion from botanical gardens are as low as the 0.1% found in other floristic inventories (Williamson 1996), help recognise taxonomic patterns in invasion risk (Daehler 1998; Pyšek 1998) and assess whether knowledge of invasion from other tropical ecosystems is a useful indicator of potential risk (Daehler et al. 2004).
Methods
Study site
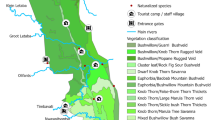
The ABG (5°05′30″ S, 38°38′10″ E) is situated in the lowland and submontane rainforests of the East Usambara Mountains in northeast Tanzania. The ABG was formally established under the German administration in 1902, although a small medicinal garden was present in the late nineteenth century (Greenway 1934; Iversen 1991). After the First World War, the British managed the gardens for agricultural research until the early 1950s, when the research station closed and the herbarium was moved to Nairobi (Iversen 1991). Over 500 species (mostly woody) were planted at ABG over a 30-year period from 1902 to 1930 in a series of trial plantations (Greenway 1934). The majority of species were introduced for potential commercial gain, with economic development of the area being the central goal (Iversen 1991; Cronk and Fuller 1995). The botanical gardens are spread over some 300 ha, and originally consisted of 20 plantation blocks, divided into 141 compartments; these compartments vary in shape and size, from 0.1 to 7 ha, and originally contained almost 2,000 species plots of varying size (Greenway 1934; Sandy et al. 1997). Greenway (1934) and Honess (1963) conducted detailed surveys of the plantations, and recorded plot location and species survival, and in many cases the planting date as well as the numbers of individuals planted. Today, approximately one third of species and plots remain.
Survey protocol
In the 1960s a total of 1,005 plots (amounting to 1,032 species plots, as a number of plots contained more than one species) containing the original species recorded by Greenway (1934) were still in existence (Honess 1963). These surviving plots were resurveyed during 2005. There were 12 compartments (containing a total of 41 plots in 1934) that were omitted from the survey that had either been destroyed, were inaccessible, or originally comprised unplanted areas containing remnant native forest trees. The 2005 resurvey accounted for just over half of those plots originally surveyed by Greenway (1934). Extant plots were found throughout the ABG including both lowland (below 700 m altitude) and submontane forest areas (above 700 m) and while 45% of the original plots from 1934 were no longer present, there was no systematic bias in the loss of plots either in relation to elevation, species or plot size (Table 2). Efforts were taken to make survey methods as systematic as possible, and plots were examined either in their entirety for smaller areas, or by walking a diagonal transect across larger plots. The survival of originally planted individuals, evidence of regeneration and the presence of other unplanted alien species were recorded. Observations of alien plant presence were made in the majority of forest areas adjoining the botanical garden, especially forest edges. In addition, where access to other forest areas was possible, further records were made. For taxa with no clear record of planting effort or location, presence or absence was also recorded in each plot. Alien species status was classified following ecologically sound definitions of naturalisation and invasion (Richardson et al. 2000b; Pyšek et al. 2004) where to be naturalised, a species must have a self-perpetuating population that is persistent independently of human intervention and further introductions, whilst evidence of significant spatial spread is indicative of invasion. Three invasion status categories were identified:
-
(1)
Regenerating: presence of seedlings, saplings, or vegetative propagation, but not necessarily naturalized;
-
(2)
Naturalised: regenerating species that have recruited new adults, but have spread to less than 10 compartments where previously unplanted. For specific life-forms this was defined as:
-
Palmoid species—presence of new adults, stemmed individuals;
-
Bamboos—presence of at least second generation individuals (for monocarpic species);
-
Lianas—new stems >5 cm diameter at breast height (dbh), or on trees other than originals;
-
Shrubs/small trees—new stems > 5 cm dbh;
-
Trees—new stems >10 cm dbh;
-
-
(3)
Spreading: a naturalised species that has spread to 10 or more new compartments. The threshold demarcating spreading species reflects a clear separation from the bulk of regenerating and naturalising species (Fig. 1).

Frequency distribution of alien plant species found in additional compartments other than those originally planted. Species found in zero new compartments were regenerating or naturalising in original compartments only. The spreading species found in five compartments was Chimonobambusa quadrangularis (spreading vegetatively)
The exceptions to this classification were species spreading vegetatively, for example the bamboos Phyllostachys spp. and Chimonobambusa quadrangularis. These species were either considered to be regenerating (1), naturalised (2), or spreading (3), with spread defined as escaping to more than one new compartment. In all cases, if adults were found outside the originally planted plots, the species was automatically considered naturalised. Existing herbaceous perennials were automatically considered naturalised, as original plants of herbaceous species were assumed to have perished after at least 75 years. All species were checked for records of naturalisation and invasion elsewhere in the tropics, using a range of sources: Cronk and Fuller (1995), Binggeli (1998), Haysom and Murphy (2003), CABI Forestry Compendium (CAB International 2005); the Global Compendium of Weeds online version (Randall 2007) and the Pacific Islands Ecosystems at Risk (PIER) project species list version 5.1 (PIER 2006). Species nomenclature follows the International Plant Names Index, accessed via the Electronic Plant Information Centre (Royal Botanic Gardens, Kew 2002), and the Germplasm Resources Information Network (GRIN) taxonomy for plants (USDA, ARS, National Genetic Resources Program).
Results
Regenerating, naturalising and spreading species
Of the 554 named alien plant species that were originally planted in the early 20th century, 214 species were still extant in 2005. Examination of the data in Honess (1963) revealed that by 1963, the families Asparagaceae, Dipterocarpaceae, Malvaceae, Fabaceae (subfamily Mimosoidae), Poaceae, Rosaceae, Rutaceae, Sapindaceae and Solanaceae were under-represented among the extant taxa, whereas the Araceae, Araucariaceae, Arecaceae, Cupressaceae, Clusiaceae and Myrtaceae were over-represented (χ2 = 74.41, df = 31, P < 0.001, across 32 families). By 2005, the Araucariaceae, Cupressaceae, Clusiaceae, Lauraceae, and Myrtaceae were still over-represented among surviving species (χ2 = 57.33, df = 31, P = 0.003, across 32 families). Survival of species between 1963 and 2005 did not vary significantly among the extant families (χ2 = 25.47, df = 19, P = 0.15, 20 families).
In 2005, out of the 214 surviving alien plant species with detailed records, 35 (16%) were regenerating but not naturalised, 38 (18%) had naturalised and a further 16 (7%) had spread to more than 10 compartments (Tables 3–5). Overall, 16% (89) of the original 554 species planted were at least regenerating, 10% (54) were at least naturalising and 3% (16) were spreading. A further 16 species with unclear introduction records were also observed as at least naturalised (Table 6). Only four species, Caesalpinia decapetala, Clidemia hirta, Olyra latifolia and Stachyphrynium repens (Table 6), were not mentioned in Greenway (1934) or Honess (1963). However, only Clidemia hirta was likely to be an unintentional introduction as the native distribution of Olyra latifolia is unclear (PIER 2006), and Caesalpinia decapetala and Stachyphrynium repens have probably escaped from nearby gardens (Table 6).
Families well represented by species that were able to regenerate included the Arecaceae, Clusiaceae, Myrtaceae and Fabaceae (subfamily Papilionoidae) (Table 3). Naturalised species failing to spread to 10 or more compartments were well represented within the Araceae and Euphorbiaceae (Table 4). Three species in the Myrtaceae and two species in each of the Arecaceae, Meliaceae and Rhamnaceae were found to have spread to 10 or more compartments (Table 5). The proportions of species that were at least naturalised varied significantly across 13 families with more than five surviving species in 2005 (χ2 = 22.36, df = 12, P = 0.03). Only in the Araceae and Euphorbiaceae were more than half of the species naturalised (4 out of 6 and 4 out of 7 species, respectively). The proportion of species that were naturalising also varied significantly but weakly across 33 families with more than five species originally introduced (χ2 = 47.65, df = 32, P = 0.05). Across all 99 families represented in ABG, there was a significant positive correlation at the family level between the number of species originally planted and the number subsequently naturalised (rho = 0.46, P < 0.001). There was also a significant positive correlation between number of surviving species in 2005 and the number of species at least naturalised (rho = 0.47, P < 0.001, for 61 families with at least one species surviving).
Colonisation patterns
Of the 56 species at least regenerating in compartments other than where originally planted, 72% were present in fewer than 10 new compartments (Fig. 1). The majority of regenerating species that were not known to have naturalised had propagated only in the compartment in which they had been originally planted (71%), with relatively few species found elsewhere and none found in more than five compartments (Fig. 1). Naturalising species were more widespread with 76% found in at least one other compartment, and a total of 16 species had spread to more than 10 new compartments (except Chimonobambusa quadrangularis, which was found in five new compartments but had spread vegetatively, thus it was considered ‘spreading’). There was also a significant correlation between compartment area and the number of alien species found that were not planted (rho = 0.46, P < 0.001, across 125 compartments with area data available), suggesting that survey efforts were systematic and in proportion with compartment size.
Only 32 species were found outside of the plantations in forest fragments and edges. The majority of regenerating species that were not known to have naturalised had propagated only in plantation/botanical garden areas (97%), with only a single sapling of Michelia champaca found in a forest area (Table 3). One third of species that had only naturalised were observed in a forest fragment or edge (Table 4). The more widely spread within the plantation area, the greater the likelihood the species would be found in neighbouring forest, with all but two of the spreading species occurring in forest (Table 5). Overall, around half of species that were at least naturalised were found in forest areas (Tables 4 and 5). Two species were only found naturalising outside of compartments—Mimosa pudica was a common weed on paths and roadsides (Table 4), and the vine Monstera deliciosa, was observed growing up trees in a forest fragment (Table 4). A third of species with unclear introduction records were observed both in plantation and forest areas (Table 6).
Invasion elsewhere
The proportion of species recorded as at least naturalised elsewhere was significantly related to invasion status in ABG (χ2 = 19.64, df = 3, P < 0.001). There was a clear association with species status in ABG and records of naturalisation and/or invasion elsewhere in the tropics. Thus, while 49% (61 out of 125) of species only surviving were recorded as naturalised or invasive elsewhere, 57% of regenerating species, 79% of naturalised species and 94% of spreading species were recorded as naturalised or invasive elsewhere (Tables 3–5; surviving only species not shown). Overall, 83% of species at least naturalising in ABG were recorded as naturalised elsewhere (Tables 4 and 5). Two-thirds of species with unclear introduction records that had at least naturalised in ABG were also recorded as naturalising elsewhere (Table 6). Only one species, the Congo Rubber Vine Landolphia owariensis, was found to be spreading in ABG but not listed as invasive elsewhere (Table 5).
Discussion
Out of the original 554 species that were introduced intentionally into ABG at the beginning of the 20th century, 39% have survived, 41% of survivors are capable of regeneration, 61% of those regenerating have naturalised, and 30% of naturalised species have spread considerably. These percentages of successful species at each stage of the invasion process clearly violate the ‘tens rule’, which states that only 10% of species would progress to the subsequent stage of the invasion pathway (Williamson and Fitter 1996). Even though only 16 species originally introduced have spread, the tens rule would predict just one species. The tens rule does not clearly hold for alien plant species introduced into ABG. However, Williamson and Fitter (1996) note a number of exceptions to the rule, and several reasons may explain why ABG and botanical gardens and arboreta in general pose a higher invasion risk than other sources of invasive plant species.
First, the deliberate cultivation of alien species in botanical gardens increases the likelihood of plant survival, by reducing competition from native species and managing the impacts of natural enemies. Such practices, often combined with extensive planting and high propagule pressure, increase the likelihood of escape and establishment in uncultivated areas (Mack 2005). Conversely, the absence of cultivation practice for accidentally introduced species would result in success of only opportunistic and ‘weedy’ species, which can tolerate a wide range of environmental conditions (Mack 2005). Historical records suggest that ABG was largely well managed until the 1960s (Honess 1963), ensuring survival of many woody species to maturity. The lack of management since the 1960s may have enabled seedlings and saplings to reach maturity, naturalise in other compartments and to spread into neighbouring forest. In contrast, perhaps only one accidentally introduced species has naturalised and spread.
Second, species and cultivars chosen for horticultural use are often selected according to traits that correlate with invasiveness (Anderson et al. 2006). Plant breeders, horticulturalists and foresters often select species on the basis of early age to maturity, fast growth rate, high seed output, and a tolerance of various environmental conditions, which would all lead to rapid domestication (Bellingham et al. 2004; Richardson and Rejmánek 2004; Anderson et al. 2006). These same traits however have been linked to invasion success, at least in disturbed ecosystems (Rejmánek and Richardson 1996; Rejmánek 2000). Given the economic thrust behind the establishment of colonial botanical gardens in the tropics, the same selection strategies may have applied in the past, therefore increasing the subsequent establishment rate of introduced taxa.
Whilst no strong taxonomic patterns were apparent among naturalised and spreading species, the Araceae and the Euphorbiaceae were well represented by naturalising species, and the Arecaceae and Myrtaceae by species in all three invasion categories (Tables 3 and 5). Although frequently naturalising, the Euphorbiaceae were not well represented in the spreading category. This result partly corroborates previous global studies on taxonomic patterns of invasive species that describe both the Euphorbiaceae and Myrtaceae as ‘invasive families’ (Daehler 1998; Pyšek 1998). However, taxonomic trends can be misleading if the introduction effort varies across families (Daehler 1998; Pyšek 1998). This study found that the number of species surviving and naturalising in a particular family in the 2005 resurvey was a function of the number of species initially planted in the early 20th century. This may account for the over-representation by the Myrtaceae and Arecaceae (the two most speciose families in ABG) and to a lesser extent the Euphorbiaceae and Araceae. Out of 99 families planted at ABG the Myrtaceae was the most numerous (45 species), followed by the Arecaceae (43 species); the Euphorbiaceae was the sixth most numerous family (23 species) whilst the Araceae family was 13th (13 species). However, only 9% (4 out of 45) of species that were originally introduced from the Myrtaceae had naturalised. In contrast, three families had disproportionately high numbers of species naturalising; these were the Bignoniaceae with 43% (3 out of 7 species), Lauraceae with 40% (2 out of 5 species), and Araceae with 30% (4 out of 13 species). Inferring real taxonomic trends in naturalisation rates is difficult, given the relatively low number of species originally introduced for these families. Thus, while some families have disproportionately high numbers of species naturalising, over-representation of some families in absolute terms will also be a consequence of greater family introduction effort.
What does ‘ invasive’ mean? The importance of definitions
For a species to be invasive, Richardson et al. (2000b) suggest somewhat arbitrarily a spread of 100 m distance in under 50 years between original plants and 2nd generation adults for (naturalised) seeded species, and more than 6 m spread over a 3-year-period for vegetative invaders. These measures of spread are probably on the low side for most invasive plants (Pyšek and Hulme 2005). Spread is used in this survey rather than invasion, as colonisation of overgrown plantations and abandoned areas by an introduced species may not constitute invasion (Richardson et al. 2000a). Furthermore, spread at ABG may only be local in compartments adjacent to those originally planted with the species. However, there does appear to be a valid distinction between the majority of species being found in fewer than 10 new compartments, and those that have spread to more than 10 compartments after 75–103 years (Fig. 1).
Many species that have spread in the ABG plantations were also observed at forest edges and in forest fragments (Table 5), suggesting that species spreading in ABG pose the greatest invasion risk to forest areas. Indeed, some may already be invasive, and the survey efforts presented here would not have detected these potential invasion events in more distant areas of forest. In total, 5% of the 554 species originally introduced to ABG have naturalised in forest areas. All but one species found in forest areas were at least naturalised in plantations, indicating that all species that are able to naturalise could be considered to have an elevated risk of invasion, especially those that have already spread considerably within the compartments. Fine (2002) reviewed instances of alien plant invasion into tropical forests, and concluded that the low number of invasion events observed globally may be attributed to low levels of human disturbance, and a lack of life history traits required for success in forests among introduced alien species, such as shade tolerance. In this study, it is evident that when alien plants are introduced to a site directly juxtaposed to existing forest, spread into forest areas can be achieved by a number of species, at least in more disturbed sites such as forest edges. The pioneer tree Maesopsis eminii is the most well known invasive plant in the East Usambaras, and has infiltrated natural forest disturbances such as treefalls, through dispersal by hornbills (Binggeli and Hamilton 1993; Cordeiro et al. 2004). A number of species spreading at ABG are known to exhibit shade tolerance, such as Syzygium jambos (CAB International 2005) and Psidium cattleianum (Cronk and Fuller 1995). It remains to be seen if such species are capable of invading or have invaded areas of undisturbed, interior forest in the East Usambaras.
Invasion elsewhere: the usual suspects?
A large proportion of naturalising and spreading species found in ABG had records of naturalisation or invasion elsewhere. Several studies have pointed out that “invasion elsewhere” is an important predictor of invasion success (Scott and Panetta 1993; Reichard and Hamilton 1997; Rejmánek 2000; Daehler et al. 2004). Interestingly, 12 of the 23 species listed in Table 1 as naturalised/invasive in other tropical botanical gardens were introduced to ABG (Greenway 1934), of which nine have at least naturalised. Only Clidemia hirta was introduced unintentionally. The same economically valuable species planted at Amani would have been introduced to many other new regions during and since the colonial era (Cronk and Fuller 1995), and if these species were planted in great numbers (i.e. they had a high introduction effort), then likelihood of naturalisation and spread would be increased (Lockwood et al. 2005). The colonial networks of botanical gardens would have ensured exchange of plant material and information on optimal planting sites (Heywood 1987; Cronk and Fuller 1995), resulting in early climate matching that would increase the likelihood of establishment. Such exchange of plant material and soil is likely to have contributed to the unintentional introduction of Clidemia hirta to areas throughout the tropics. Alternatively, species introduced throughout the tropics may also possess certain traits that predispose them to successful naturalisation and invasion in the introduced range (Rejmánek and Richardson 1996). For example, species planted for their edible fruits may attract new mutualist frugivorous birds and animals in the introduced range that will aid in dispersal and establishment of an escaped population (Richardson et al. 2000a; Hulme 2006). Further, a number of species mentioned here have long been used and cultivated by local people, including Cedrela odorata and Artocarpus heterophyllus (Jackfruit); their spread in ABG may be partially explained by human factors, with villages, paths and roads located throughout the botanical garden plantations and surrounding forest.
The result obtained for ABG reinforces the importance of using previous invasion history in botanical garden invasion risk assessments. However, there will be exceptions; one species was spreading at ABG that had not been recorded elsewhere. In contrast, nearly half of those species only surviving (not regenerating) were also recorded as naturalised/invasive elsewhere in the tropics. The use of the “invasion elsewhere” criterion is only valuable if the species has been introduced before, and preferably, widely. In circumstances where a species is introduced to a region for the first time or no records of past introductions are kept, then intuitively “invasion elsewhere” is not especially helpful. Further, the absence of a species from other lists may be due to a lack of adequate records where this species is actually a problem. Invasion success of any alien introduced species will be context dependent, to some extent, and relying too heavily on “invasion elsewhere” as a predictive tool could lead to large numbers of low risk species being misidentified as higher risk, whilst novel invasive species may go unnoticed, and become a problem.
Implications for other botanical gardens
There is a growing awareness among botanical gardens and horticultural bodies that the risk of alien plant escape and invasion from cultivation should be formally assessed (Baskin 2002; Mack 2005). Both the Missouri and Chicago Botanical Gardens have devised and implemented protocols to assess invasion risk from established collections (Baskin 2002; Jefferson et al. 2004) but this approach has yet to be applied in the tropics. The Chicago Botanical Garden model presents an integrated approach, incorporating the use of an adapted Weed Risk Assessment scheme, regional invasive plant lists, on-site plant evaluation of collections and information on commercial availability (Jefferson et al. 2004). The case study of ABG presented here highlights that botanical gardens should concentrate risk assessment efforts on plant species introduced intentionally, whilst also aiming to minimise the risk of unintentional introductions, by avoiding transport of plants in soil from elsewhere.
It is widely recognised that plant invasions are likely to occur but that alien plant introductions are on the whole socially, economically and scientifically beneficial (Dosman and Del Tredici 2003; Mack 2005; Anderson et al. 2006). However, it should be possible to meet human needs for introduction of new taxa, whilst simultaneously reducing the likelihood of invasion by the most pernicious alien plant species. Botanical gardens have a potentially unique role to play in preventing and mitigating plant invasions, and would benefit from adopting a methodologically rigorous protocol as presented here, to quantify the risk of invasion from their own collections into surrounding ecosystems that may be of significant conservation value. As such, assessing invasion risk should become an integral part of conservation goals for all botanical gardens, and not just those located in the tropics. The survey and methods used have highlighted some key issues to consider regarding patterns in species survival, naturalisation and spread, invasion terminology, and knowledge of invasion elsewhere.
Abbreviations
- ABG:
-
Amani Botanical Garden
References
Anderson NO, Galatowitsch SM, Gomez N (2006) Selection strategies to reduce invasive potential in introduced plants. Euphytica 148:203–216
Baskin Y (2002) The greening of horticulture: new codes of conduct aim to curb plant invasions. Bioscience 52:464–471
Bellingham PJ, Duncan RP et al (2004) Seedling growth rate and survival do not predict invasiveness in naturalized woody plants in New Zealand. Oikos 106:308–316
Bellingham PJ, Tanner EVJ, Healey JR (2005) Hurricane disturbance accelerates invasion by the alien tree Pittosporum undulatum in Jamaican montane rain forests. J Veg Sci 16:675–684
Binggeli P (1998) An overview of invasive woody plants in the tropics. School of Agricultural and Forest Sciences publication number 13, University of Wales, Bangor
Binggeli P, Hamilton AC (1993) Biological invasion by Maesopsis eminii in the East Usambara forests, Tanzania. Opera Bot 121:229–235
CAB International (2005) Forestry compendium (CD Rom). Wallingford, UK
Cordeiro NJ, Patrick DAG et al (2004) Role of dispersal in the invasion of an exotic tree in an East African submontane rainforest. J Trop Ecol 20:449–457
Cronk QCB, Fuller JL (1995) Plant invaders: the threat to natural ecosystems. Chapman & Hall, London
Daehler CC (1998) The taxonomic distribution of invasive angiosperm plants: ecological insights and comparison to agricultural weeds. Biol Conserv 84:167–180
Daehler CC, Denslow JS et al (2004) A risk-assessment system for screening out invasive pest plants from Hawaii and other Pacific Islands. Conserv Biol 18:360–368
Dehnen-Schmutz K, Touza J et al (2007) The horticultural trade and ornamental plant invasions in Britain. Conserv Biol 21:224–231
Dosman M, Del Tredici P (2003) Plant introduction, distribution and survival: a case study of the 1980 Sino-American botanical expedition. Bioscience 53:588–597
Fine PVA (2002) The invasibility of tropical forests by exotic plants. J Trop Ecol 18:687–705
Greenway PJ (1934) Report of a botanical survey of the indigenous and exotic plants in cultivation at the East African Agricultural Research Station, Amani, Tanganyika Territory
Hartemink AE (2001) Biomass and nutrient accumulation of Piper aduncum and Imperata cylindrica fallows in the humid lowlands of Papua New Guinea. For Ecol Manage 144:19–32
Haysom KA, Murphy ST (2003) The status of invasiveness of forest tree species outside their natural habitat: a global review and discussion paper. In: Forest ecology and management. Forest health and biosecurity working paper FBS/3E. Forestry Department, FAO, Rome
Heywood VH (1987) The changing role of the botanic garden. In: Bramwell D, Hamann O, Heywood V, Synge H (eds) Botanic gardens and the world conservation strategy. IUCN, Academic Press, London, pp 3–18
Honess BL (1963) The Amani plantations: a detailed report on a survey made in 1961–1962. Agricultural Officer, Muheza, Tanzania
Hulme PE (2003) Biological invasions: winning the science battles but losing the conservation war? Oryx 37:178–193
Hulme PE (2005) Nursery crimes: agriculture as victim and perpetrator in the spread of invasive species. In: British Crop Protection Council (eds) Crop sciences and technology: proceedings of an international congress held in Glasgow. 31st October–2nd November 2005, pp 744–740
Hulme PE (2006) Beyond control: wider implications for the management of biological invasions. J Appl Ecol 43:835–847
Hulme PE, Bacher S, Kenis M, Klotz S, Kühn I, Minchin D, Nentwig W, Olenin S, Panov V, Pergl J, Pyšek P, Roques A, Sol D, Solarz W, Vilà M Grasping at the routes of biological invasions: a framework to better integrate pathways into policy. J Appl Ecol (in press)
ISSG—Invasive Species Specialist Group (2007) Global invasive species database. http://www.issg.org/database/welcome/. Cited Feb 2007
Iversen ST (1991) The Usambara Mountains, NE Tanzania: phytogeography of the vascular plant flora. Dissertation, Acta Universitatis Upsaliensis XXIX: 3, Almqvist and Wiksell International, Stockholm
Jayeola AA (2006) Stemming the menace of alien plant invasion through botanic garden education. African Botanic Gardens Network Bulletin 11
Jefferson L, Havens K, Ault J (2004) Implementing invasive screening procedures: the Chicago Botanic Garden model. Weed Technol 18:1434–1440
Kembelo K (1996) The botanical gardens of Zaire and the present state of biodiversity in Zaire. Bot Gard Conserv News 2:7
Leadlay E (2004) Report of the 2nd world botanic gardens congress, Barcelona, Spain. Bot Gard J 1(1). http://www.bgci.org/worldwide/article/229/. Cited October, 2007
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228
Lorence DH, Sussman RW (1986) Exotic species invasion into Mauritius wet forest remnants. J Trop Ecol 2:147–162
Mack RN (2005) Predicting the identity of plant invaders: future contributions from horticulture. Hortscience 40:1168–1174
Marambe B (2001) Alien invasive plants in Sri Lanka: current concerns and future perspectives. In: Balakrishna P (ed) Report of the workshop on alien invasive species, global biodiversity forum, South and Southeast Asia session, October 1999, Colombo, Sri Lanka. IUCN, pp 61–66
Maunder M (1994) Botanical gardens—future challenges and responsibilities. Biodivers Conserv 3:97–103
Maunder M, Higgens S, Culham A (2001) The effectiveness of botanic garden collections in supporting plant conservation: a European case study. Biodivers Conserv 10:383–401
McKey D (1988) Cecropia peltata, an introduced pioneer tropical tree, is replacing Musanga cecropioides in Southwestern Cameroon. Biotropica 20:262–264
Meier W (2006) Contribution to the knowledge of Persicaria nepalensis (Meisn.) H.Gross and P.capitata (Buch.-Ham. ex D.Don) H.Gross, invasive species in theAvila National Park, Venezuela. Acta Bot Ven 29:1–16
Meyer JY (1998) Observations on the reproductive biology of Miconia calvescens DC (Melastomataceae), an alien invasive tree on the island of Tahiti (South Pacific Ocean). Biotropica 30:609–624
Miller IL, Lonsdale WM (1987) Early records of Mimosa pigra in the Northern Territory. Plant Prot Q 2:140–142
Muller T (1994) The role a botanical institute can play in the conservation of the terrestrial biodiversity in a developing country. Biodivers Conserv 3:116–125
Newmark WD (2002) Conserving biodiversity in Eastern African forests: a study of the Eastern Arc Mountains. Ecological studies, vol 155. Springer, Berlin
PIER—Pacific Island Ecosystems at Risk (2006) Plant threats to Pacific ecosystems: index of all PIER species listed by scientific name, version 5.1. Institute of Pacific Islands Forestry/US Forest Service and Department of Agriculture. http://www.hear.org/pier/scientificnames/index.html. Cited Jan 2007
Peters WL, Meyer MH, Anderson NO (2006) Minnesota horticultural industry survey on invasive plants. Euphytica 148:75–86
Pyšek P (1998) Is there a taxonomic pattern to plant invasions? Oikos 82:282–294
Pyšek P, Hulme PE (2005) Spatio-temporal dynamics of plant invasions: linking pattern to process. Ecoscience 12:302–315
Pyšek P, Richardson DM et al (2004) Alien plants in checklists and floras: towards better communication between taxonomists and ecologists. Taxon 53:131–143
Randall RP (2007) Global compendium of weeds (online verision). Dated 24th January 2007. http://www.hear.org/gcw/. Cited Nov 2007
Reichard SH (2004) Conflicting values and common goals: codes of conduct to reduce the threat invasive species. Weed Technol 18:1503–1507
Reichard SH, Hamilton CW (1997) Predicting invasions of woody plants introduced into North America. Conserv Biol 11:193–203
Reichard SH, White P (2001) Horticulture as a pathway of invasive plant introduction in the United States. Bioscience 51:103–113
Rejmánek M (2000) Invasive plants: approaches and predictions. Austral Ecol 25:497–506
Rejmánek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77:1655–1661
Richardson DM, Rejmánek M (2004) Conifers as invasive aliens: a global survey and predictive framework. Divers Distrib 10:321–331
Richardson DM, Pyšek P et al (2000a) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107
Richardson DM, Allsopp N et al (2000b) Plant invasions—the role of mutualisms. Biol Rev 75:65–93
Royal Botanic Gardens, Kew (2002) Electronic Plant Information Centre. http://www.kew.org/epic/. Cited Sept 2006
Sandy RF, Boniface G, Rajabu I (1997) A survey and inventory of the Amani Botanical Garden. East Usambara catchment forest project technical paper 38. Forest and Beekeeping Division/Finnish Forest and Park Service, Dar es Salaam/Vantaa
Scott JK, Panetta FD (1993) Predicting the Australian weed status of Southern African plants. J Biogeogr 20:87–93
Sheil D (1994) Naturalized and invasive plant species in the evergreen forests of the East Usambara Mountains, Tanzania. Afr J Ecol 32:66–71
USDA, ARS, National Genetic Resources Program. Germplasm Resources Information Network—(GRIN) [Online Database]. National Germplasm Resources Laboratory, Beltsville, Maryland. http://www.ars-grin.gov/cgi-bin/npgs/html/index.pl. Cited February 2007
Vaughan RE, Wiehe PO (1937) Studies on the vegetation of Mauritius: I. A preliminary survey of the plant communities. J Ecol 25:289–343
Waylen K (2006) Botanic gardens: more than just a pretty place. Oryx 40:259–260
Weber E (2003) Invasive plant species of the world: a reference guide to environmental weeds. CABI Publishing, Wallingford
Whitmore TC (1991) Invasive woody plants in perhumid tropical climates. In: Ramakrishnan PS (ed) Ecology of biological invasions in the tropics. International Scientific Publications, New Delhi, pp 35–40
Wijesundra S (1999) Alien invasive species in Sri Lanka and their history of introduction. In: Marambe B (ed) Proceedings of the first national workshop on alien invasive species. Ministry of Forestry and Environment, Sri Lanka, pp 25–27
Williamson MH (1996) Biological invasions. Chapman & Hall, London
Williamson MH, Fitter A (1996) The characters of successful invaders. Biol Conserv 78:163–170
Acknowledgements
This work is part of the Darwin Initiative project “Combating alien invasive plants threatening the East Usambara mountains in Tanzania” (162/13/033) and authors are grateful for financial support from Defra and NERC. The authors would also like to thank the staff of the Amani Nature Reserve, especially Mr. Corodius Sawe, and the Tropical Biology Association, especially Dr. Rosie Trevelyan, for logistical support.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dawson, W., Mndolwa, A.S., Burslem, D.F.R.P. et al. Assessing the risks of plant invasions arising from collections in tropical botanical gardens. Biodivers Conserv 17, 1979–1995 (2008). https://doi.org/10.1007/s10531-008-9345-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-008-9345-0