
Abstract
A Lake Michigan Ecosystem Model (LM-Eco) that includes a detailed description of trophic levels and their interactions was developed for Lake Michigan. The LM-Eco model constitutes a first step toward a comprehensive Lake Michigan ecosystem productivity model to investigate ecosystem-level responses and effects within the lower food web of the lake. The effect of the invasive species Bythotrephes longimanus on individual zooplankton species was investigated based upon extensive field data collected at multiple locations in Lake Michigan during the 1994–1995 Lake Michigan Mass Balance Study. Field data collected at 15 sampling stations within Lake Michigan over a series of 8 sampling cruises throughout a 2 year period demonstrated that over 65% of zooplankton species exhibited a decline with the occurrence of Bythotrephes in the sample. The LM-Eco model was successfully applied to simulate the trends of Bythotrephes and zooplankton abundance as observed in the collected field data. Model simulations allowed for examination of interactions between the invader Bythotrephes and native zooplankton groups on a 5 km by 5 km resolution throughout Lake Michigan. Analysis was completed as a time series specific to individual field sampling locations within the lake, and also on a lake-wide scale.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Understanding the temporal and spatial distribution of nonindigenous aquatic nuisance species, as well as their effects on other native species within the Great Lakes ecosystem, is essential for future management (Hassan et al. 2005; Great Lakes Commission 2006; International Joint Commission 2006). In the Great Lakes Commission’s Strategic Plan, the Aquatic Nuisance Species issue is a key objective calling for “healthy ecosystems where new introductions of nonindigenous aquatic nuisance species are prevented and adverse ecological and economic impacts of species already present are minimized (Great Lakes Commission 2006).”
The Great Lakes Water Quality Agreement of 1978 enacts the maintenance of chemical, physical, and biological integrity of the Great Lakes via monitoring and management. The Lake Michigan Mass Balance Study (LMMBS) was conducted by the United States Environmental Protection Agency (USEPA) during the 1994 and 1995 time period to gain better understanding of Lake Michigan through monitoring and modeling (US Environmental Protection Agency 1997a, b; Richardson et al. 1999). As part of this study, a thorough sampling of Lake Michigan was made with respect to organisms within the lower food web. We used data obtained from the LMMBS, and developed a Lake Michigan Ecosystem Model (LM-Eco), which includes a detailed description of ecosystem dynamics within the lower food web of the water column of the lake. This modeling framework contains multiple compartments, including compartments for two zooplankton groups, herbivorous zooplankton and carnivorous zooplankton, as well as a compartment for the nonindigenous aquatic nuisance species Bythotrephes longimanus. The present analysis is unique, in that the model development utilized an extensive data set that comprises an unprecedented lake-wide data collection effort consisting of data from over 100 stations sampled regularly over the 2 year LMMBS period. The LM-Eco model was constructed using high spatial resolution, in that the resolution of the model consists of a 5 km grid containing 44,042 cells applied to Lake Michigan. The LM-Eco model construct comprises biological and physical components, as both the hydrodynamics of the lake system and the kinetics corresponding to the organismal compartments operate within the grid.
In the current study, the LM-ECO model was applied specifically to simulate the effect of the invasive, nonindigenous species Bythotrephes on zooplankton abundance. Bythotrephes is a predatory cladoceran that is native to northern Europe and Asia, but invaded the Laurentian Great Lakes in the early 1980s (Johannsson et al. 1991). While the distribution of this species currently includes all five Great Lakes, the species was first reported in Lake Michigan in 1986 (Lehman 1987; Evans 1988; Pothoven et al. 2003). Transport of Bythotrephes to the Laurentian Great Lakes has been hypothesized as occurring through contaminated ship ballast water (Jin and Sprules 1990; Pothoven et al. 2001). Bythotrephes has become established in Lake Michigan, and as a result has impacted the composition of the zooplankton community within the lake, both on an annual basis and in terms of long term trends (Lehman 1988; Lehman and Caceres 1993; Burkhardt and Lehman 1994; Barbiero and Tuchman 2004). We examined the trend of Bythotrephes with herbivorous and carnivorous zooplankton species during an annual cycle for 1994 and 1995 using field data collected from the LMMBS. We used the LM-Eco model to simulate trends observed from analysis of the field data. In applying the LM-Eco model, spatial and temporal trends were examined for Bythotrephes and zooplankton relative to a high resolution (within a 5 km by 5 km grid cell), site specific basis within the lake, whereby they were compared to field samples taken at sampling stations specific to the LMMBS. In addition, spatial and temporal patterns were analyzed for Bythotrephes and zooplankton groups on a lake wide scale.
Methods
Model description
The LM-Eco model was designed as a combination of physical and biological components. The physical model was developed as a high resolution model, consisting of a 5 km × 5 km grid applied to the surface of Lake Michigan and containing 19 vertical sigma layers. Each vertical sigma layer contains 2,318 cells for a total of 44,042 modeled grid cells. Hydrodynamics in the LM-Eco model are calculated externally using output from a version of the Princeton Ocean Model (POM) developed originally by Blumberg and Mellor (1987), and later configured for Lake Michigan geometry (Beletsky and Schwab 1998). The POM was driven using observed meteorological forces, which include winds and heat flux (Liu and Schwab 1987; Beletsky and Schwab 1998). Output from the POM consisted of water temperature, horizontal and vertical dispersion, and horizontal and vertical currents for each grid cell (Liu and Schwab 1987; Beletsky and Schwab 1998). The hydrodynamics from the POM are for the time period of April 1, 1994 through December 21, 1995. The LM-Eco model utilized a transport method based upon the “ULTIMATE QUICKEST” transport scheme developed by Leonard (1991) and subsequently augmented for use with variable grid sizes (Cerco and Cole 1995; Chapman et al. 1997). The LM-Eco model calculations included numerical integration of spatially varying particle concentrations using quadratic interpolation of the concentration to infer a value at flow faces, as well as analytic integration over space and time to account for changes in the concentration at the grid cell wall during each time step.
The biological model is constructed based upon field studies of the observed lower food web as it existed within the water column of Lake Michigan during the time period of 1994 through 1995 coinciding with the LMMBS (Madenjian et al. 2002). The biological model is based upon a proposed model framework for the LMMBS (Richardson et al. 1999) which includes a series of mathematical equations for construct of a lower food web ecosystem model for Lake Michigan (LimnoTech 2000), but this framework has been enhanced to address the impact of the invasive species Bythotrephes on the lower food web, and specifically the zooplankton community structure, of Lake Michigan. The equations describing the biomass (g/m3) of herbivorous zooplankton, carnivorous zooplankton, and Bythotrephes, respectively, are given in terms of the basal metabolic rates, BMHZ, BMCZ, and BMB (/day), upper trophic level predation rates, UHZ, UCZ, and UB (g/m3/day), grazing rates, GRHZ, GRCZ, and GRB (/day), and grazing efficiencies, GREFHZ, GREFCZ, and GREFB:
Herbivorous zooplankton (HZ):
Carnivorous zooplankton (CZ):
Bythotrephes (B)
Model application
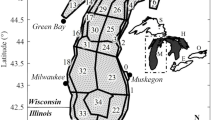
The LM-Eco model was applied to examine population dynamics of interacting compartments within the lower food web of the water column of Lake Michigan for the time period of 1994 and 1995, coinciding with data collected from the LMMBS. A specific emphasis was placed in the current study upon interactions between the invasive species Bythotrephes and zooplankton. The LMMBS included data collected within Lake Michigan during eight sampling cruises occurring from April 24, 1994 through October 13, 1995 and spread throughout the lake (Table 1). A detailed description of sampling techniques and sample analysis can be found in the Lake Michigan Mass Balance Project Methods Compendium (US Environmental Protection Agency 1997a). All data collected as part of the LMMBS were subjected to water quality assurance procedures (US Environmental Protection Agency 1997a, 2001). Bythotrephes was sampled at 15 sample stations located throughout Lake Michigan (Fig. 1), and was found only in the last two sample cruises of each year in the study (Table 1). The occurrence of Bythotrephes in the samples taken in the later portion of the year is consistent with other studies which have found that Bythotrephes does not occur until water temperatures reach 17–20°C, and is dominant with populations rising rapidly starting in late July and lasting throughout the later portion of the year (Pothoven et al. 2001). A total of 48 species of zooplankton were also sampled at these 15 sampling stations (Table 3). The zooplankton species were compartmentalized into herbivorous zooplankton (HZ) and carnivorous zooplankton (CZ) to be consistent with the LM-Eco model construct. The predator to prey ratio (CZ to HZ ratio) was examined using the field data during the time period in which Bythotrephes was sampled in the annual cycle at these 15 stations.

The invasive species Bythotrephes was sampled at 15 field sampling sites within Lake Michigan as part of the Lake Michigan Mass Balance Study
The LM-Eco model simulations were conducted over the time period of April 1, 1994 through December 21, 1995, which coincides with the simulation timeframe of the Princeton Ocean Model as applied to Lake Michigan. The initial distributions of biological variables within the lake were constructed based upon field data collected from Cruise 1 of the LMMBS (Table 1). Output from the model was tabulated using a daily time step for comparison between modeled values and field data. Model parameterization was completed based upon field studies specific to Lake Michigan or one of the other Great Lakes (LimnoTech 2000). Differences in grazing efficiency and temperature preferences between Bythotrephes, carnivorous zooplankton, and herbivorous zooplankton were captured in model parameterization, based upon documented field studies with the Great Lakes Region (LimnoTech 2000; Table 2). The model was calibrated following the methods of Thomann (1982), whereby field data versus model plots and techniques including regression of field data on model output were used to guide a fit of the model to the field data.
The output from the LM-Eco model simulation was compared to field data on a high resolution, site specific basis in the lake. For a given sample station, trends for Bythotrephes, herbivorous zooplankton, and carnivorous zooplankton were examined over the 21 month simulation of the LMMBS study period and compared to field collected data. In addition, predator to prey ratio (CZ to HZ ratio) was predicted by the model corresponding with the presences of Bythotrephes during the annual cycle on a high resolution, site specific basis in the lake, and this ratio was compared to the predator to prey ratio observed in the field corresponding to the given sampling locations. Further, lake wide trends in Bythotrephes, herbivorous zooplankton, and carnivorous zooplankton were produced by the LM-Eco model, enabling spatial and temporal patterns of these interacting compartments to be estimated over the course of the LMMBS study period.
Results
The invasive species Bythotrephes was collected at 15 sampling stations during the field study for the LMMBS, and 48 species of zooplankton were also found at these 15 sites (Fig. 1). In addition, Bythotrephes was found in samples collected during the later two cruises of each sampling year (Table 1). As Bythotrephes exhibits a cyclic pattern in abundance, trends corresponding to each of the 48 species of zooplankton were analyzed specifically to determine if there was an effect on a given species abundance corresponding with the presence of Bythotrephes during the annual cycle. For each species of zooplankton, we compared the median value of its distribution among the 15 stations in the absence of the invasive species Bythotrephes to that in the presence of Bythotrephes within an annual cycle. Using this approach, 32 species of zooplankton showed a decrease in abundance and 16 zooplankton species showed an increase in abundance corresponding with the presence of the invasive species Bythotrephes in the sample within an annual cycle (Table 3). Further, we applied the nonparametric Wilcoxin signed ranks test (α = 0.05) to compare distributions of each zooplankton species in the absence of Bythotrephes to those in the presence of Bythotrephes during an annual cycle. It was determined that nine zooplankton species exhibited a statistically significant decline with the presence of Bythotrephes (Table 3). No zooplankton species were found to exhibit a statistically significant increase corresponding to the presence of Bythotrephes in the sample (Table 3). Trends were also observed by compartmentalizing the zooplankton species, as they are grouped into herbivorous zooplankton and carnivorous zooplankton within the LM-Eco modeling framework. We applied the nonparametric Wilcoxin signed ranks test (α = 0.05) to compare distributions of the predator to prey ratio (carnivorous zooplankton to herbivorous zooplankton ratio) at each sampling station in the absence of the invasive species Bythotrephes to the same ratio in the presence of Bythotrephes at these stations during an annual cycle. It was determined that there was a statistically significant decline in the ratio of carnivorous zooplankton to herbivorous zooplankton corresponding with the occurrence of Bythotrephes during an annual cycle (P < 0.01). There was also a decreasing trend in the median carnivorous to herbivorous zooplankton ratio recorded during the time period in which Bythotrephes existed in samples throughout the later part of the year encompassing the later two sampling cruises annually (median value of decrease in the carnivorous to herbivorous ratio was 8.6% over the 15 stations sampled). Other notable trends include application of the Wilcoxin signed ranks test (α = 0.05) resulting in determination of a statistically significant decrease in abundance of herbivorous zooplankton (P < 0.01) and carnivorous zooplankton (P < 0.05) during the time period in which the invasive species Bythotrephes occurred in the annual cycle.
We used the LM-Eco model, and produced simulations to examine trends for the interaction of the invasive species Bythotrephes with carnivorous and herbivorous zooplankton. Each simulation began on April 1, 1994 and ended on December 21, 1995, and output from the LM-Eco model was compared to field data. The LM-Eco model produced output for trends associated with the invasive species Bythotrephes, herbivorous zooplankton, and carnivorous zooplankton specific to any selected 5 km by 5 km grid cell. This, in turn, allowed comparison between trends predicted by the LM-Eco model within a 5 km by 5 km model grid cell and field data collected at a particular sampling station in Lake Michigan during the LMMBS, given that the longitude and latitude of the sampling station placed it within the corresponding 5 km by 5 km grid cell. The LM-Eco model was capable of capturing field data collected at a particular sample site in Lake Michigan with respect to interacting compartments (the invasive species Bythotrephes, carnivorous zooplankton, herbivorous zooplankton), as well as predicting trends between sampling events of these interacting groups over time at that site. As an illustration, the LM-Eco model simulated trends as compared to field collected data for Bythotrephes, carnivorous zooplankton, and herbivorous zooplankton at grid cell number 859, which contains the LMMBS field sampling station number 240, located at a latitude of 43.349° and a longitude of −87.170° (Fig. 2). The LM-Eco model output for the ratio of carnivorous zooplankton to herbivorous zooplankton (predator to prey ratio) was compared to the ratio obtained from field data collected among the 15 sample stations. The LM-Eco model simulated a general pattern of decline in carnivorous to herbivorous zooplankton (predator to prey ratio) that occurred among the 15 sampling locations with the presence of Bythotrephes in the sample (Fig. 3).

LM-Eco model simulated trends compared with field collected data for herbivorous zooplankton (a), carnivorous zooplankton (b), and the invasive species Bythotrephes (c) at model grid cell 859/44,042 which contains the LMMBS field sampling station 240 located at latitude of 43.349° and a longitude of −87.170°

LM-Eco model simulated carnivorous zooplankton to herbivorous zooplankton (predator to prey) ratio compared to the ratio of carnivorous zooplankton to herbivorous zooplankton recorded at 15 field sampling locations in which the invasive species Bythotrephes was present
Further, the LM-Eco model was applied to examine lake-wide trends for the interaction of the invasive species Bythotrephes with herbivorous and carnivorous zooplankton. This application of the model allows for examining spatial and temporal patterns associated with interactions occurring among all 44,042 segments within the model. As an example, LM-Eco model output for lake wide spatial patterns of the interacting compartments (Bythotrephes, herbivorous zooplankton, and carnivorous zooplankton) are illustrated that corresponded to June 29, 1994 and August 1, 1994 (Fig. 4). Sequential spatial and temporal plots were used to examine general patterns among interacting compartments, such as the decline of both herbivorous and carnivorous zooplankton with the presence of Bythotrephes, and an increase in abundance of Bythotrephes beginning in late June with a continued increase throughout July and August, followed by declines in November and December. A decrease in both herbivorous and carnivorous zooplankton coinciding with an increase of Bythotrephes is evident in comparison of the spatial patterns for these groups corresponding to June 29, 1994 (Fig. 4a) with those from August 1, 1994 (Fig. 4b).

LM-Eco Lake Michigan model simulation for herbivorous zooplankton, carnivorous zooplankton, and the invasive species Bythotrephes abundances. Output is shown for the simulation time step corresponding to the date of June 29, 1994 (a) and the simulation time step corresponding to the date August 1, 1994 (b). Lake wide simulation output of the abundances within the model’s 5 km × 5 km grid cells allows for examination of spatial and temporal patterns between interacting trophic levels on a scale corresponding to the entire lake
Discussion
An “ecosystem approach” to investigating the effects of aquatic invasive species could be defined as an approach that addresses ecosystem functions and attributes that support water quality and aquatic biodiversity in both open and near-shore areas of water bodies. A measure of the health of a given ecosystem and its attendant functions can be expressed in terms of the species and communities present, and most importantly the processes and energy flows between them (Cardinale et al. 2006). Aquatic invasive species are hypothesized to be the leading cause of biodiversity change in lakes and to play a major role in extinctions of species within North American freshwater ecosystems in the coming century (Ricciardi and Rasmussen 1999; Sala et al. 2000). Thus, there is a need to be able to monitor spatial and temporal patterns associated with a given invasive species, and to use these patterns as a tool for understanding the impacts of the exotic species on the pre-invaded community. The current study provides a practical example of how field collected data can be used within an ecosystem productivity modeling framework to support prediction of the spatial and temporal distribution of an invasive species, Bythotrephes, and also the ecological impacts of the invasive species on the pre-invaded zooplankton community structure of a large lake ecosystem.
A major advantage in development of the LM-Eco model presented here is the use of the Lake Michigan Mass Balance Study data set. The LMMBS constitutes a thorough and unusual monitoring effort of Lake Michigan over the 1994 and 1995 time period, and represents the most spatially and temporally comprehensive lake-wide data set ever collected over a 2 year period within the lake (US Environmental Protection Agency 1997b). This type of monitoring is essential in developing a high resolution model containing a grid system with thousands of cells, and provides a sound scientific base of information to guide model formulation, as well as evaluation of model output. Our examination of field data collected at 15 sampling stations within the lake over a series of 8 sampling cruises throughout the 2 year LMMBP study period revealed a majority (over 65%) of zooplankton species exhibited a decline with the presence of Bythotrephes within the annual cycle, and this is consistent with several other studies in which the invader is hypothesized to have caused changes in distribution of abundance of zooplankton species (Lehman 1987, 1988, 1991; Sandgren and Lehman 1990; Lehman and Caceres 1993; Pothoven et al. 2003; Barbiero and Tuchman 2004). Utilizing the measurements for Bythotrephes and zooplankton abundances at these sites within the development of a large-scale high resolution mathematical model (the LM-Eco model) allowed for the estimation of site specific and lake-wide spatial and temporal patterns occurring between zooplankton groups and the invader over the 640 days simulation period. This application is unique in its spatial aspects, as the LM-Eco model allows for the ability to examine trends for interactions between zooplankton groups and the invasive species Bythotrephes on a resolution of 5 km by 5 km locations throughout the entire lake. Further, the LM-Eco model allows for examination of temporal patterns between zooplankton groups and the invader at any given location in Lake Michigan over an annual cycle, thereby encouraging the collection of daily or weekly field samples for use in comparison between model and field applications from site specific locations in the lake. As demonstrated in the results of this study, comparisons between the ratio of interacting compartments (predator and prey) can be projected by the model to summarize trends for interactions over time, and to formulate patterns that might be expected resulting from disruption to the food web (Figs. 3, 4).
Several advancements in technology may allow for future data collection efforts that will interface well with the high resolution LM-Eco model developed here. The Great Lakes Observing System (GLOS), the Great Lakes node of the national Integrated Ocean Observing System (IOOS), is an interagency cooperative effort based on a sustained network of buoys, ships, satellites, underwater vehicles, and other platforms that routinely collect real time data and store information (Great Lakes Observation System 2007). The GLOS has been proposed as capable of enhancing the LMMBS study, by allowing for provision of GLOS monitoring programs so that modeling tools can, in turn, continue to improve our understanding of the Lake Michigan ecosystem. In addition to the GLOS, other applications of remote sensing of lake water quality parameters including new and developing hyperspectral plane-based and boat-based sensors will allow for data collection on a high resolution basis in Lake Michigan (Heiskary et al. 2006). A laser optical plankton counter (LOPC) has recently been applied by the USEPA Mid-Continent Ecology Division for monitoring and assessment of the lower food web of Lake Superior (Yurista et al. 2005, 2009).
In summary, the LMMBS included an unprecedented data collection effort that allowed for an appropriate data base from which to construct a lower food web ecosystem model for the water column of Lake Michigan. The Lake Michigan Ecosystem Model constitutes a first step in utilizing a high resolution model to investigate invasive species dynamics within the lower food web of the lake. Thorough monitoring of the Great Lakes would be necessary to continue to observe new impacts as they occur within the lower food web of the lake. A comprehensive lake wide survey for Lake Michigan, comparable to the LMMBS, is planned to take place throughout 2010. The LM-Eco model as presented in this study represents the ecology of the Lake Michigan food web within the water column of the lake as it existed during the 1994–1995 LMMBS period (Madenjian et al. 2002; Vanderploeg et al. 2002). Thus, the model could serve as a starting point from which a more detailed model could be constructed that includes sediment dwelling organisms based upon future comprehensive field survey results. Refinement to this modeling framework, based upon data collected and our understanding of changes to the lower food web of Lake Michigan since the LMMBS, could be used as an investigative tool to forecast the spatial and temporal effects of invasive species and their impact on ecosystem level dynamics within the lower food web of the lake.
References
Barbiero RP, Tuchman ML (2004) Changes in the crustacean communities of Lakes Michigan, Huron, and Erie following the invasion of the predatory cladoceran Bythotrephes longimanus. Can J Fish Aquat Sci 61:2111–2125
Beletsky D, Schwab DJ (1998) Modeling thermal structure and circulation in Lake Michigan. In: Abstracts of the estuarine and coastal modeling fifth international conference, Alexandria, 22–24 Oct 1997
Bierman VJ, Dolan DM (1981) Modeling phytoplankton-nutrient dynamics in Saginaw Bay, Lake Huron. J Gt Lakes Res 7:409–439
Bierman VJ, Dolan DM (1986) Modeling of phytoplankton in Saginaw Bay: I. Calibration phase. J Environ Eng 112:415–429
Blumberg AF, Mellor GL (1987) A description of a three-dimensional coastal ocean circulation model. Coast Estuar Sci 4:1–6
Burkhardt S, Lehman JT (1994) Prey consumption and predatory effects of an invertebrate predator (Bythotrephes: Cladocera, Cercopagidae) based on phosphorus budgets. Limnol Oceanogr 39:1007–1019
Canale RP (1976) Modeling biochemical processes in aquatic ecosystems. Ann Arbor Science, Ann Arbor
Canale RP, Depalma LM, Vogel AH (1975) A plankton based food web model for Lake Michigan. In: Canale RP, Depalma LM, Vogel AH (eds) Modeling biochemical processes in aquatic ecosystems. Sea Grant Program, Ann Arbor
Cardinale BJ, Srivastava DS, Duffy JE, Wright JP, Downing AL, Sankaran M, Jouseau C (2006) Effects of biodiversity on the functioning of trophic groups and ecosystems. Nature 443:989–992
Cerco CF, Cole T (1995) Three dimensional eutrophication model of Chesapeake Bay. J Environ Eng 119:1006–1025
Chapman RS, Cole TM, Gerald TK (1997) Development of a hydrodynamic/water quality (POM-IPXMT) linkage for the Lake Michigan mass balance project, final report. U.S. Environmental Protection Agency, Office of Research and Development, ERL-Duluth, Large Lakes Research Station, Grosse Ile
Evans MS (1988) Bythotrephes cederstroemi: its new appearance in Lake Michigan. J Gt Lakes Res 14:234–240
Great Lakes Commission (2006) Great Lakes aquatic nuisance species. http://www.glc.org/ans/. Accessed 08 Aug 2007
Great Lakes Observation System (2007) About GLOS—The Great Lakes node of the National Integrated Ocean Observing System. http://glos.us/. Accessed 10 Dec 2007
Hassan R, Scholes R, Ash N (2005) Ecosystems and human well-being: current state and trends, vol 1, findings of the condition and trends working group of the millennium ecosystem assessment. Island Press, WA
Heiskary S, Olmanson L, Gitelson A, Chipman J (2006) Remote sensing: does it have a role? Lakeline 26:27–35
International Joint Commission (2006) Thirteenth Biennial report. http://www.ijc.org/en/publications/pdf/13br_e.pdf. Accessed 28 May 2008
Jin EH, Sprules WG (1990) Distribution and abundance of Bythotrephes cederstroemii (Cladocera: Cercopagidae) in the St. Lawrence Great Lakes. Verh Int Ver Limnol 24:383–385
Johannsson OE, Mills EL, O’Gorman R (1991) Changes in the near shore and offshore zooplankton communities in Lake Ontario. Can J Fish Aquat Sci 48:1546–1557
Lehman JT (1987) Palearctic predator invades North American Great Lakes. Oecologia 74:478–480
Lehman JT (1988) Algal biomass unaffected by food web changes in Lake Michigan. Nature 332:537–538
Lehman JT (1991) Causes and consequences of cladoceran dynamics in Lake Michigan: implications of species invasion by Bythotrephes. J Gt Lakes Res 17:437–445
Lehman JT, Caceres CE (1993) Food-web responses to species invasion by a predatory invertebrate Bythotrephes in Lake Michigan: implications of species invasion by Bythotrephes. Limnol Oceanogr 38:879–891
Leonard B (1991) The ULTIMATE conservative difference scheme applied to unsteady one-dimensional advection. Comp Methods Appl Mech Eng 88:17–74
LimnoTech (2000) Ecosystem model for the Lake Michigan mass balance study. U.S. Environmental Protection Agency, Office of Research and Development, ERL-Duluth, Large Lakes Research Station, Grosse Ile
Liu PC, Schwab DJ (1987) A comparison of methods for estimating U* from given Uz and air-sea temperature differences. J Geophys Res 92:6488–6494
Madenjian C, Fahnenstiel G, Johengen T, Nalepa T, Vanderploeg H, Fleischer G, Schneeberger P, Benjamin D, Smith E, Bence J, Rutherford E, Lavis D, Robertson D, Jude D, Ebener M (2002) Dynamics of the Lake Michigan food web, 1970–2000. Can J Fish Aquat Sci 59:736–753
Pothoven SA, Fahnenstiel GL, Vanderploeg HA (2001) Population dynamics of Bythotrephes cederstroemii in south-east Lake Michigan 1995–1998. Freshw Biol 46:1491–1501
Pothoven SA, Fahnenstiel GL, Vanderploeg HA (2003) Population characteristics of Bythotrephes in Lake Michigan. J Gt Lakes Res 29:145–156
Ricciardi A, Rasmussen JB (1999) Extinction rates of North American freshwater fauna. Conserv Biol 3:1220–1222
Richardson WL, Endicott DD, Kreis RG Jr, Rygwelski KR (1999) The Lake Michigan mass balance project quality assurance plan for mathematical modeling. US Environmental Protection Agency, Office of Research and Development, Mid-Continent Ecology Division, Large Lakes Research Station, Grosse Ile, EPA/600/R-04/018
Sala OE, Chapin FS III, Armesto J, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leeman RS, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Sandgren CD, Lehman JT (1990) Response of chlorophyll a, phytoplankton, and microzooplankton to the invasion of Lake Michigan by Bythotrephes. Verh Int Ver Limnol 24:386–392
Scavia D, Lang GA, Kitchell JF (1988) Dynamics of Lake Michigan plankton: a model evaluation of nutrient loading, competition, and predation. Can J Fish Aquat Sci 45:165–177
Thomann RV (1982) Verification of water quality models. J Environ Eng 108:933–940
US Environmental Protection Agency (1997a) Lake Michigan mass balance study (LMMB) methods compendium, vol 1: sample collection techniques. U.S. Environmental Protection Agency, Great Lakes National Program Office, Chicago, EPA/905/R-97/017
US Environmental Protection Agency (1997b) Lake Michigan mass budget/mass balance work plan. US Environmental Protection Agency, Great Lakes National Program Office, Chicago, EPA/905/R-97/018
US Environmental Protection Agency (2001) The Lake Michigan mass balance study quality assurance report. US Environmental Protection Agency, Great Lakes National Program Office, Chicago, EPA/905/R-01/013
Vanderploeg HA, Nalepa TF, Jude DJ, Mills EL, Holeck KT, Liebig JR, Grigorovich IA, Ojaveer H (2002) Dispersal and emerging ecological impacts of Ponto-Caspian species in the Laurentian Great Lakes. Can J Fish Aquat Sci 59:1209–1228
Yurista PM, Kelly JR, Miller S (2005) Evaluation of optically acquired zooplankton size-spectrum data as a potential tool for assessment of condition in the Great Lakes. Environ Manag 35:34–44
Yurista PM, Kelly JR, Miller S (2009) Lake Superior zooplankton biomass: alternate estimates from a probability-based net survey and spatially extensive LOPC surveys. J Gt Lakes Res 35:337–346
Acknowledgments
This study was conducted in support of the Invasive Species Initiative in the Office of Research and Development (ORD) of the USEPA. We thank Peder Yurista and Henry Vanderploeg for providing a valuable review of this manuscript. The authors also wish to acknowledge the Great Lakes National Program Office for their efforts in the Lake Michigan Mass Balance Study, as well as the many cooperators during the study. In addition, the authors wish to acknowledge NOAA, GLERL, and LimnoTech for their cooperation and collaboration. The views expressed in this article are those of the authors and do not necessarily reflect the views or policies of the U.S. Environmental Protection Agency. Mention of trade names or commercial products does not constitute endorsement or recommendation for use.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Miller, D.H., Kreis, R.G., Huang, WC. et al. Application of a lower food web ecosystem productivity model for investigating dynamics of the invasive species Bythotrephes longimanus in Lake Michigan. Biol Invasions 12, 3513–3524 (2010). https://doi.org/10.1007/s10530-010-9748-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-010-9748-1