
Abstract
Food web models are powerful tools to inform management of lake ecosystems, where top-down (predation) and bottom-up (resource) controls likely propagate through multiple trophic levels because of strong predator–prey links. We used the Ecopath with Ecosim modeling approach to assess these controls on the Lake Huron main basin food web and the 2003 collapse of an invasive pelagic prey fish, alewife (Alosa pseudoharengus). We parameterized two Ecopath models to characterize food web changes occurring between two study periods of 1981–1985 and 1998–2002. We also built an Ecosim model and simulated food web time-dynamics under scenarios representing different levels of top-down control by Chinook salmon (Oncorhynchus tshawytscha) and of bottom-up control by quagga mussels (Dreissena rostriformis bugensis) and nutrients. Ecopath results showed an increase in the relative importance of bottom-up controls between the two periods, as production decreased across all trophic levels. The production of non-dreissenid benthos decreased most, which could cause decreases in production of pelagic prey fishes feeding on them. Ecosim simulation results indicated that the alewife collapse was caused by a combination of top-down and bottom-up controls. Results showed that while controls by Chinook salmon were relatively constant before alewife collapse, controls by quagga mussels and nutrients increased jointly to unsustainable levels. Under current conditions of low nutrients and high quagga mussel biomass, simulation results showed that recovery of alewives is unlikely regardless of Chinook salmon biomass in Lake Huron, which implies that the shrinking prey base cannot support the same level of salmonine predators as that prevailed during the 1980s.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Understanding the relative importance of top-down (predation) and bottom-up (resource) controls on ecosystem structures is a key to successful ecosystem management. In lake ecosystems, controls are more likely to propagate through whole food webs than in other ecosystems as they are characterized by stronger species interactions (Borer and others 2005). This characteristic makes lake ecosystems especially vulnerable to human activities that alter top-down controls such as fisheries (Pauly and others 2002) or bottom-up controls such as watershed agricultural practices (Smith 2003). However, the same characteristic also makes manipulating controls an applicable management tool (for example, Shapiro and Wright 1984).
The Laurentian Great Lakes have been continuously affected by invasive species, overexploitation, habitat alterations, and management practices that have altered top-down and bottom-up controls on food web dynamics (Gaden and others 2012). In the early twentieth century, invasive sea lamprey (Petromyzon marinus) and alewives (Alosa pseudoharengus) reached Lake Huron from the Atlantic Ocean through the Welland Canal that allowed them to bypass the Niagara Falls (Ebener and others 1995). Overfishing and mortality imposed by parasitic sea lamprey caused sharp declines in commercial fishery harvests and the abundance of lake trout (Salvelinus namaycush), the only dominant native predator in pelagic waters, around 1950 (Berst and Spangler 1972), which facilitated the establishment of the planktivorous prey fish alewife (Miller 1957). In response, the USA and Canada management agencies started to control sea lamprey in the late 1950s mainly through the application of chemical lampricides targeting sedentary larval stages in streams (Smith and Tibbles 1980). In the 1960s, management agencies started to stock exotic predators including coho and Chinook salmon (Oncorhynchus kisutch and O. tshawytscha) to create recreational fisheries (Tody and Tanner 1966). Since the mid-1990s, there has been a general decreasing trend in prey fish biomass in Lake Huron, and the biomass of alewives abruptly decreased by more than 90% (“collapsed”) between 2002 and 2003 (Riley and others 2008). However, it is unclear if the collapse of alewives was caused by top-down control because alewives also were affected by bottom-up controls from nutrient reduction and invasive dreissenid mussel (zebra and quagga mussels, Dreissena polymorpha and D. rostriformis bugensis) filtration. Starting in the late 1990s, dreissenid filtration reduced phytoplankton biomass, increased water clarity (Vanderploeg and others 2002), and sequestered nutrients that would be otherwise available to the offshore planktonic food web in nearshore zones (Hecky and others 2004), thus causing changes in zooplankton and benthos community structures (Higgins and Vander Zanden 2010). In the same period, nutrient loads and concentrations were reduced by phosphorus abatement programs initiated in the 1970s (Dolan and Chapra 2012).
In this study, we assessed relative importance of top-down control imposed by Chinook salmon and other top predators and bottom-up controls from dreissenid filtration and nutrient reduction on the 2003 collapse of alewife population in Lake Huron. Previous studies have investigated top-down or bottom-up controls in Lake Huron in relation to trophic shifts in the main basin. For example, He and others (2015) quantified piscivory patterns and showed that salmonine consumption might have exceeded prey production soon after 2000, while Nalepa and others (2007) showed changes in benthos community, with increases in the abundance of quagga mussel and decreases in abundances of amphipod Diporeia spp. and oligochaetes that were important food to fishes. However, there is a lack of understanding of the relative importance of these controls on the alewife collapse, which could interact in complex ways (McQueen and others 1989) and occurred simultaneously in the Lake Huron food web.
Given the collapse of alewife in Lake Huron in 2003, Dettmers and others (2012) pointed out that resource managers face an important dilemma: whether to manage for economically important recreational fisheries that rely on stocked exotic species or for native species that may better adapt to ongoing ecosystem changes. Recreational harvests of stocked Pacific salmonines in Lake Huron generally decreased with decreases in prey fish biomass after 2000 (Su and He 2013). However, the collapse of the alewife population may benefit recruitment of native fishery species including lake trout, walleye (Sander vitreus), and yellow perch (Perca flavescens) that were negatively affected by alewives (Madenjian and others 2008). To address this management dilemma, it is crucial for resource managers to understand how food web dynamics may be affected by management actions given an evolving ecosystem state where importance of top-down (such as predator stocking) and bottom-up (such as nutrient loading) controls can be altered.
Assessing relative importance of top-down and bottom-up controls in a large ecosystem like Lake Huron may only be achieved in a timely fashion by using ecological models (Jørgensen and others 2012). These models integrate process knowledge and data collected in focal ecosystems. Thus, they are powerful tools that can be used to untangle evolution of effects among concurrent factors in ecosystems where manipulative experiments are not feasible and statistical analyses are limited by temporal lags between effects and responses. In this study, we used the Ecopath with Ecosim (EwE) modeling approach (Christensen and Walters 2004), which consisted of components for understanding trophic interactions among food web groups (Ecopath) and simulating time-dynamics under designed scenarios (Ecosim). We configured and implemented Lake Huron EwE models to: (1) characterize changes in trophic interactions among food web groups between the reference period 1981–1985 and the period 1998–2002 before alewives collapsed; and (2) simulate food web time-dynamics under top-down and bottom-up control scenarios.
Methods
Study Area
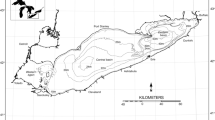
Lake Huron is shared by USA’s State of Michigan and Canada’s Province of Ontario (Figure 1) and is comprised of four subbasins: North Channel, Georgian Bay, Saginaw Bay, and the main basin. We modeled the food web in the main basin, which is deep and oligotrophic with an area of 3.78 × 104 km2 (63% lake surface) and mean and maximum depths of 73 and 229 m (Beeton and Saylor 1995). Major water inflow is from Lake Superior via St. Mary’s River but the most important nutrient input is the Saginaw Bay outflow (Dolan and Chapra 2012). The basin is connected to Lake Michigan via the Straits of Mackinac and to Lake Erie via the St. Clair River–Detroit River corridor.

Lake Huron map.
Ecopath with Ecosim (EwE) Modeling Approach
The EwE modeling approach was developed based on conservation of biomass. Details for model derivation, program software, and software documentation are available at http://www.ecopath.org/. We used the software EwE version 6.3.
Lake Huron Ecopath Models
We parameterized 1984 and 2002 Ecopath models to represent Lake Huron food webs during the 1981–1985 and 1998–2002 periods, respectively. Available biomass data (as described Supplementary Material) suggested that food webs in both periods were in relatively steady-state condition, which is an assumption of the Ecopath analysis (Christensen and Pauly 1992).
In Ecopath, food web groups are connected via feeding links using a system of linear equations:
where B i is the biomass (g/m2 in wet weight) of group i, (P/B) i is annual average production to biomass ratio, EE i is ecotrophic efficiency, B j and (Q/B) j are the biomass and annual average consumption to biomass ratio of group j, DC ij is the proportion of group i in diet of group j, BA i is annual biomass accumulation, Y i is annual fishery yield, and E i is the annual net migration (emigration–immigration). EE i represents the proportion of production of group i that is lost to predation or exported through fishing and migration. The second equation in Ecopath represents energy balance for each consumer group i:
where (P/Q) i , (R/Q) i , and (U/Q) i are proportions of consumption represented by production, respiration, and unassimilated food, respectively. Energy balance of producer groups was ensured at parameterization as their production equals net primary production (Christensen and others 2008).
Food Web Configuration, Model Inputs, and Data Sources
We configured 52 groups in the 1984 Ecopath model and 55 in the 2002 model (Tables 1 and 2) based on information and community structure data described in Supplementary Material. Three additional groups into the 2002 model represent species that invaded Lake Huron in the 1990s: zebra mussel, quagga mussel, and round goby (Neogobius melanostomus). To represent trophic ontogeny and selectivity to fisheries, we parameterized 10 out of the 15 fish taxa as multistanza groups (Christensen and others 2008) with up to four age stanzas. For example, “Lake trout 0–0.5″ stands for larvae that are vulnerable to alewife predation, “Lake trout 0.5–1″ for fingerlings whose biomass is supplemented by stocking, and “Lake trout 5+” for age 5 and older, the only stanza vulnerable to fisheries. Food web groups can be categorized into eight functional components (Table 1): Top predators (17 groups), Lake whitefish (3 stanza groups), Pelagic prey fishes (6 groups), Benthic prey fishes (10 groups), Zooplankton (6 groups), Benthos (7 groups), Microplankton (four groups including cyanophytes, other phytoplankton, protozoa, and pelagic bacteria), and Detritus (2 groups).
Input parameters required for these Ecopath models included B, P/B for groups with only one stanza, total mortality rate Z for multistanza groups, Q/B, U/Q, DC, and Y. We detail data sources and estimation for input parameters in Supplementary Material. P/B and Q/B inputs were the same for most groups in the 1984 and 2002 models (Table 2) but diet composition inputs were modified in the 2002 model to reflect changes in food web structure (Tables 3 and 4). We set BA for all groups to zero as the food web was in a relatively steady-state in both periods. We also set E to zero. To represent temporary presence in the main basin for double-crested cormorant (Phalacrocorax auritus), sea lamprey, some salmonine species, and walleye, we estimated input parameters representing the predator–prey interactions exclusively in the main basin. For example, the Q/B of Double-crested cormorants was based on consumption estimates only during the breeding season when they feed in the main basin (Ridgway and Fielder 2012). For Chinook salmon 5+ and Other salmonines 5+ that migrate into streams during spawning runs and die, we set their diets as 100% “Import” and detritus fates as 100% “Export” so that consumption and detritus generation would not affect model biomass balance (Christensen and others 2008).
Ecotrophic efficiency (EE) and respiration to consumption ratio (R/Q) were estimated by Ecopath through solving a set of linear equations (1) and (2). We slightly modified biomass and diet composition inputs to ensure that biomass balance was achieved (EE < 1) for all groups. We also ensured that energy balance was achieved (R/Q > 0) at parameterization.
Characterizing Changes in Food Web Structure
We first summarized changes in production, biomass, and ecological size of the Lake Huron food web between 1981–1985 and 1998–2002 periods. We used total system throughput (TST) as measure of food web ecological size based on Finn (1976):
where Q T is total consumption, R T is total respiration, FD T is total flow into detritus, and Ex T is total food web exports. Finn (1976) suggested that proportion of TST components Q T , Ex T , R T , and FD T can be used to characterize food web structure. Ecopath tool “Statistics” automatically calculates food web total production, total biomass, and TST (Christensen and others 2008). Note that detritus group biomass is excluded in total biomass calculation. Because detritus is nonliving organic matter, detritus biomass stands for an Ecopath model parameter, not a real biomass in ecological context.
Characterizing Changes in Interactions Among Food Web Groups
To characterize Lake Huron food web trophic interactions during 1981–1985 and 1998–2002 periods, we summarized production, biomass, and ecotrophic efficiencies for trophic levels I–IV. Change in production by trophic level is an indicator of change in relative importance of top-down and bottom-up controls in a food web. If the relative importance of top-down control increases, trophic cascade theory (Carpenter and others 1985) suggests that production changes will vary among trophic levels in an alternating manner, with increases in production for trophic levels IV and II, and decreases for levels III and I. If relative importance of bottom-up control increases, production should decrease across all trophic levels (McQueen and others 1986).
We calculated biomass by trophic level using “Trophic level decomposition” routine outputs in the Ecopath tool “Network analysis”, in which each group biomass is apportioned into discrete trophic levels based on diet composition inputs (Christensen and others 2008). We calculated production P and EE by trophic level l (l = I, II, III, and IV):
where B i,l is the biomass of a non-detritus group i that is apportioned into trophic level l.
To further characterize trophic interactions in Lake Huron food web in both periods, we summarized production, biomass, ecotrophic efficiency, and allocation of production by functional component. We also quantified changes in production, biomass, and ecotrophic efficiency and identified major changes in production allocation for each functional component between the two periods.
Lake Huron Ecosim Dynamic Modeling
We used Ecosim to simulate time-dynamics of the Lake Huron food web under scenarios of Chinook salmon and quagga mussel biomass and of nutrient levels. The Ecosim master equation is the derivative form of Ecopath master equation (1), which represents how biomass B of group i changes with time:
where G i is dimensionless gross conversion efficiency, Q ji is consumption on group j by group i, Q ij is predation on group i by group j, E i is net migration rate, M0 i is nonpredatory natural mortality rate, and F i is the fishing mortality rate.
Consumption in Ecosim is modeled based on the foraging arena theory (Walters and others 1997):
where p ij is predation rate on prey group i by unit biomass of predator group j that can be adjusted by forcing functions S ij and mediation functions M ij ; v ij is vulnerability parameter; and f 1 , f 2 , and f 3 are functions representing effects of feeding time and handling time on consumption (Christensen and Walters 2004).
We used the 1984 Ecopath model as initial conditions of Ecosim, with slight modifications related to inclusion of invasive groups Round goby, Zebra Mussel, and Quagga mussel so that they could be incorporated in simulations, as did in Kao and others (2014). We set low biomass values (0.25 g/m2 for Round goby, and 0.60 g/m2 for Zebra and Quagga mussels) and used 2002 Ecopath model values for parameters P/B, Q/B, and U/Q (Table 2). Correspondingly, we modified diet composition inputs as described in Supplementary Material. We set biomass inputs for obtaining ecotrophic efficiencies of about 0.50 for Round goby and about 0.15 for Zebra mussel and Quagga mussel. Ecotrophic efficiencies were based on 2002 Ecopath outputs and the literature data on temporal changes in feeding preferences among groups in response to invasive species. For example, feeding preference of double-crested cormorants shifted from pelagic alewives and rainbow smelt (Osmerus mordax) to benthic round goby (Johnson and others 2010), while Lake whitefish 3+ preference shifted from soft-bodied benthos Diporeia spp. to zebra and quagga mussels (Pothoven and Nalepa 2006) and round goby (Pothoven and Madenjian 2013). Further details for Ecosim dynamic modeling are given in Supplementary Material.
Ecosim Calibration
To calibrate the Ecosim model, we estimated vulnerability parameters by fitting simulated biomass to available biomass time-series data. We included groups with more than 3 years of available data during 1984–2006. To judge goodness of fit among groups after calibration, we calculated root-mean-squared deviations (RMSD) between simulated and observed biomasses after natural logarithm transformation.
Simulation Scenarios
To assess the relative importance of top-down and bottom-up controls on the Lake Huron alewife collapse, we conducted simulations under scenarios that represented a range of historical levels of Chinook salmon biomass, quagga mussel biomass, and nutrient loads. For Chinook salmon scenarios, we used 21 biomass levels of Chinook salmon 1–4 ranging from 0.00 g/m2 to 0.20 g/m2. Biomass inputs of other Chinook salmon stanza groups were estimated by Ecopath. For quagga scenarios, we used five biomass levels ranging from 0 g/m2 to 80 g/m2. For nutrient scenarios, we used three total phosphorus (TP) load levels: 2575 tonnes/year (corresponding to 15.0 µg/l input concentration), 1803 tonnes/year (10.5 µg/l) and 1526 tonnes/year (8.9 µg/l). These levels represent average TP loads for the periods between 1984 and 1997, 1998 and 2002, and 2003 and 2006, respectively.
We designed the analysis as a factorial experiment and ran simulations under scenarios representing different levels of controls by each factor (that is, Chinook salmon, quagga mussel, and nutrients) and their combinations. We ran simulations for a 40-year period after 2006, last year of the model calibration period, within which the food web was expected to reach equilibrium conditions, and summarized changes in simulated equilibrium biomass of Alewife 1+. We considered alewives as collapsed when equilibrium biomass fell below 0.01 g/m2. In scenarios representing controls by one factor, we kept the other two factors unchanged: to simulate nutrient loading effects, we kept biomass of Chinook salmon and quagga mussel at 0 g/m2; to simulate effects of either Chinook salmon or quagga mussel, we kept the other group at 0 g/m2 and nutrients constant at the high level (2575 tonnes/year). In scenarios representing controls by two factors, we kept the third factor at 0 g/m2 for either Chinook salmon or Quagga mussel, or at the high level for nutrients. In simulations, we used stocking biomass for Lake trout 0.5–1 and Other salmonines 0 groups corresponding to reported 2006 levels. Stocking of Chinook salmon 0–0.5 in simulations was estimated by Ecopath in order to generate Chinook salmon biomass 1–4 defined for each of the 21 levels used in simulation scenarios. For groups subjected to fishery harvests, we used observed fishing mortality rates in 2006.
Results
Ecopath Analyses: Changes at the Food Web Level
Ecopath results showed decreases in productivity and ecological size, as measured by total system throughput—the sum of total consumption, total respiration, total flow into detritus, and total exports, of the Lake Huron food web between the 1981–1985 and 1998–2002 periods, and an increase in standing biomass. Total production decreased by 34% and total system throughput (TST) decreased by 32%, while total biomass increased by 14% (Table 5). All components in TST decreased. Total consumption and total respiration decreased by about 20% while total flow into detritus and total exports decreased by about 45%.
Ecopath analyses also showed decreases in production across all trophic levels in the Lake Huron food web (Table 6), which indicated an increase in the relative importance of bottom-up controls between the 1981–1985 and 1998–2002 periods. However, changes in productivity were not uniform across trophic levels. Production decreased by 38 and 23% for levels I and II and by 8% and 18% for levels III and IV. On the other hand, standing biomass decreased by 18% and 31% for trophic levels I and IV but increased by 23 and 9% for levels II and III. Decreases in production were not matched with decreases in biomass for levels II and III because biomass of invasive dreissenids, among the least productive species in the food web, was apportioned to these two levels based on diet. In the 2002 Ecopath model, dreissenid mussels comprised 33% and about 50% of biomass in levels II and III. Ecopath estimated ecotrophic efficiencies, which represented the proportion of prey production that is consumed by predators or harvested by fisheries in this study, increased by 34 and 18% for levels I and II but decreased by 6 and 13% for trophic levels III and IV.
Ecopath Analyses: Changes at the Functional Component Level
Results from Ecopath analyses for all functional components in the pelagic pathway (Microplankton → Zooplankton → Pelagic prey fishes → Top predator) of the Lake Huron food web showed decreases in production and biomass between the 1981–1985 and 1998–2002 periods (Table 7). Microplankton production and biomass decreased by 37 and 22%, as production and biomass of groups in this functional component generally decreased (Table 2). Zooplankton production and biomass decreased much less, by about 5%, with minor changes (<11%) among Cladocerans, Cyclopoids, and Calanoids groups that made up about 80% of total biomass of this functional component. Production and biomass of Pelagic prey fishes decreased by about 60%, the largest decrease among all pelagic functional components. The decrease reflected a 78% decrease in Rainbow smelt biomass. Production and biomass of Top predators decreased by about 30%, with decreases in biomass of all fish groups in this functional component except for Lake trout 5+ and Walleye.
In the pelagic pathway, ecotrophic efficiencies (EE) increased for Microplankton and Top predators but decreased for Zooplankton and Pelagic prey fishes between the two periods. Microplankton EE increased by 34% (Table 7) mainly because the proportion of their production consumed by Zooplankton increased from 38 to 56% (Figure 2). Zooplankton EE decreased by 10%, mainly because the proportion of their production consumed by Pelagic prey fishes decreased from 28 to 17% while the proportion of their production consumed by groups within the functional component increased only from 5 to 12%. Pelagic prey fishes EE decreased by 21% mainly because the proportion of their production consumed by groups within the functional component decreased from 38 to 19%. The proportion of Pelagic prey fishes production consumed by Top Predators actually increased from 8 to 13%. Top Predators EE increased by 63% mainly because the proportion of their production exported to fisheries increased from 10 to 20%.

Summary of Ecopath outputs for trophic level and allocation of production by functional component from the 1984 and 2002 Lake Huron food web models. Each pie chart shows proportions of production of a functional component that were allocated to predation by corresponding functional components, exported through fisheries, or accumulated as detritus.
Among functional components in the benthic pathway of the Lake Huron food web (Detritus → Benthos → Benthic prey fishes → Lake whitefish), results from Ecopath showed different direction changes in production and biomass between the 1981–1985 and 1998–2002 periods (Table 7). Detritus production decreased by 41% mainly because detritus that originated from Microplankton decreased by 57%. Biomass of the two Detritus groups decreased by about 45% (Table 2). Benthos production decreased by 14%, although biomass increased by 76% due to large additional biomass from dreissenid groups (Zebra mussel and Quagga mussel). This decrease in production resulted from decreases in biomass of Amphipods and Oligochaetes that are more productive than dreissenid groups. Production and biomass of Benthic prey fishes increased by 17 and 8%, mainly from additional production and biomass of invasive Round goby. Lake whitefish production and biomass increased by 140 and 97%, the largest increases between the two periods among all functional components.
EE in the benthic pathway increased for Detritus and Benthic prey fishes but decreased for Benthos and Lake whitefish between the two periods (Table 7). Detritus EE increased by 17% mainly because the proportion of Detritus production consumed by Microplankton increased from 30 and 37% (Figure 2). Benthos EE decreased slightly by 4%. However, the proportion of Benthos production consumed by Pelagic prey fishes decreased from 37 to 19% while the proportion consumed by Lake whitefish increased from 6 to 17%. Benthic prey fishes EE increased by more than 400%, reflecting increases in consumptive demands of all functional components that fed on Benthic prey fishes. Lake whitefish EE decreased by 38% mainly due to a decrease from 13 to 10% in the proportion of production exported through fisheries.
Ecosim Calibration
After calibration, time series of simulated biomass of the 22 groups selected for analysis generally tracked observed time series (Figure 3). Among fish groups, calibration fits were better for predator groups than for prey fish groups. Root-mean-square deviations (RMSD) between simulated and observed biomasses after natural logarithm transformation were 0.13–0.53 for predator groups and 0.63–0.96 for prey fish groups. For groups in lower trophic levels (zooplankton, benthos, and phytoplankton groups), observed versus simulated comparisons were as good as for prey fish groups. RMSD were 0.43–1.15 for zooplankton groups, 0.39–0.97 for benthos groups, and 0.71–0.79 for phytoplankton groups. Trends of simulated and observed biomass time series were very consistent for all groups. However, simulated biomass time series of groups in lower trophic levels did not have large inter-annual variations that were common to observed biomass time series.

Calibration results of the Lake Huron Ecosim model for groups with more than 3 years of available biomass data. The line represents the best-fit biomass, and circles are observed biomass time series in the calibration period from 1984 to 2006. RMSD is the root-mean-square deviation between simulated and observed biomasses after natural logarithm transformation.
Ecosim Simulations: Factorial Experiment on Alewife 1+ Biomass
Alewife 1+ biomass reached equilibrium within 20 simulation years under all scenarios that represented different levels of controls by each factor (Chinook salmon, quagga mussel, and nutrients) and their combinations. For example, simulated Alewife 1+ biomass, under selected Chinook salmon scenarios ranging from absence to 0.15 g/m2, reached equilibrium within 5 years when quagga mussel scenario was fixed at a low level (20 g/m2) and nutrient scenario at the high level (Figure 4).

Calibration and part of simulation results for Alewife 1+ biomass from Ecosim. The line from 1984 to 2006 represents the best-fit biomass and circles are observed biomass time series. Lines after 2007 represent 40-year simulations under scenarios of four different levels of Chinook salmon biomass, a low level of Quagga mussel biomass of 20 g/m2, and the high level of nutrient loads (2575 tonnes/year of total phosphorus). The 20-year biomass average after reaching equilibrium in each scenario corresponds to a point of the same scenario in Figure 5.
In one-factor analyses, equilibrium Alewife 1+ biomass decreased linearly with increases in Chinook salmon and quagga mussel biomass and with decreases in nutrient loads (Figures 5A–C). However, equilibrium Alewife 1 + biomass reached a constant level regardless of increases over a 0.12 g/m2 upper Chinook salmon biomass limit (CH UL ). The highest equilibrium Alewife 1+ biomass, 1.01 g/m2, occurred under scenarios of absence of Chinook salmon and quagga mussel and high nutrient level. Within Chinook salmon single-factor analysis, equilibrium Alewife 1+ biomass decreased from 1.01 to 0.81 g/m2 (Figure 5A) and within quagga mussel single-factor analysis, equilibrium biomass decreased to 0.50 g/m2 (Figure 5B). The lowest equilibrium Alewife 1+ biomass, 0.35 g/m2, occurred at the low nutrient level within nutrient single-factor analysis (Figure 5C).

Simulated Alewife 1+ biomass under all scenarios of Chinook salmon biomass, Quagga mussel biomass, and nutrient loads. The analysis was designed as a factorial experiment with three factors: Chinook salmon, quagga mussel, and nutrients. Each point represents a 20-year biomass average after reaching equilibrium under a scenario representing effects of one factor or a combination of two or three factors.
In two-factor analyses, Alewife 1+ equilibrium biomass decreased to below 0.01 g/m2 (that is, collapsed) in a scenario of high quagga mussel biomass (80 g/m2) and low nutrient level, CH UL varied with quagga mussel biomass and nutrient level, and the pattern of decreasing trend within quagga mussel scenarios varied with nutrient level (Figures 5D–F). Within Chinook salmon and quagga mussel analysis, equilibrium Alewife 1+ biomass decreased with increases in quagga biomass from 1.01 to 0.50 g/m2 under Chinook salmon absence scenarios and from 0.81 to 0.33 g/m2 under maximum Chinook salmon biomass scenarios (Figure 5D). Within Chinook salmon and nutrient analysis, equilibrium Alewife 1+ biomass decreased with decreases in nutrients from 1.01 to 0.35 g/m2 under Chinook salmon absence scenarios and from 0.81 to 0.16 g/m2 under maximum Chinook salmon biomass scenarios (Figure 5E). Within simulation ranges, CH UL decreased with increases in quagga mussel biomass from 0.12 to 0.08 g/m2, and with decreases in nutrients from 0.12 to 0.06 g/m2. However, before reaching CH UL , the decrease of Alewife 1+ equilibrium biomass with per unit increase in Chinook salmon biomass was intensified with increases in quagga mussel biomass and decreases in nutrient levels. Within quagga mussel and nutrient analysis, equilibrium Alewife 1+ biomass decreased with nutrient decreases from 1.01 to 0.35 g/m2 under quagga mussel absence scenarios and from 0.50 g/m2 to below 0.01 g/m2 under maximum quagga mussel biomass scenarios (Figure 5F). Further, the decrease of Alewife 1+ biomass with per unit increase in quagga biomass was higher at higher nutrient levels.
In three-factor analyses, Alewife 1+ equilibrium biomass decreased to below 0.01 g/m2 in several scenarios (Figure 5G). Among low nutrient level scenarios, Alewife 1 + equilibrium biomass decreased to below 0.01 g/m2 in scenarios where Chinook salmon biomass was greater than 0.02 g/m2 and quagga biomass was greater than 60 g/m2 (Figure 5G left panel). When quagga mussel biomass scenarios increased from 0 to 40 g/m2 at the low nutrient level, Alewife 1+ equilibrium biomass decreased from 0.16 to 0.08 g/m2 in scenarios of maximum Chinook salmon biomass and CH UL decreased from 0.05 to 0.03 g/m2. Among median nutrient level scenarios, Alewife 1+ equilibrium biomass decreased from 0.34 to 0.02 g/m2 in scenarios of maximum Chinook salmon biomass and CH UL decreased from 0.07 to 0.03 g/m2 with increases in quagga mussel biomass scenarios (Figure 5G central panel). Note that results for high nutrient level scenarios in three-factor analyses are the same as results for Chinook salmon and quagga mussel analysis in two-factor analyses described in previous paragraph (i.e., Figure 5G right panel is the same as Figure 5D).
Discussion
Overview and Synthesis
Although alewife production and biomass in Lake Huron changed little between 1981–1985 and 1998–2002 periods as indicated in our Ecopath analyses, an increase in relative importance of bottom-up controls in the food web portended the alewife collapse. An increase in relative importance of bottom-up controls in the Lake Huron food web can be inferred from our Ecopath analyses that showed (1) decreases in production across all trophic levels, (2) increases in efficiencies of utilizing primary production and recycling detritus, and (3) increases in ecotrophic efficiencies for lower trophic levels but not for higher trophic levels. These indicators of increasing bottom-up controls were associated with decline in nutrients and invasions of dreissenid mussels.
Ecopath analyses showed that the functional component Benthos formed a bottleneck for production transferring from lower to higher trophic levels after dreissenid invasions. Decreases in primary production did not result in major decreases in total production for Zooplankton and Benthos (Table 7). However, among benthos groups, production of non-dreissenid benthos (mainly Amphipods and Oligochates) decreased drastically between the 1981–1985 and 1998–2002 periods (Table 2). We estimated from Ecopath analysis that about half of non-dreissenid benthos production, on which most fish species feed, was replaced by dreissenid production. Consequently, almost all production of non-dreissenid benthos (94%) was consumed in the 1998–2002 period, while only 26% of dreissenid production was consumed. Limited non-dreissenid benthos availability likely caused decreases in production for pelagic prey fishes, in particular rainbow smelt (about 80% decrease in production), and their predators. Although a sharp decrease in alewife biomass had not occurred until 2002, results from Ecopath analyses imply that consumptive demand by predators on alewives would increase with the sharp decrease in rainbow smelt biomass.
Ecosim simulation scenarios shed further light on the relative importance of top-down (Chinook salmon) and bottom-up (quagga mussel and nutrient) controls that could have caused collapse of alewives in Lake Huron. Simulated alewife biomass nearly collapsed under a scenario representing conditions observed in 2003 and 2004: median biomass levels for Chinook salmon (0.07 g/m2) and quagga mussel (40 g/m2) and the low level of nutrients (1526 tonnes/year of total phosphorus loads). Therefore, although alewives were still abundant in 2002, our simulations showed that their population would be at risk if conditions during 2003–2004 persisted, which indeed happened. After 2004, low nutrient loads persisted (Dolan and Chapra 2012) and quagga mussel biomass increased to high levels corresponding to our simulation scenarios (Thomas Nalepa, University of Michigan, unpublished data). Under such conditions, our simulations showed that bottom-up controls from reduced nutrients and quagga mussel filtration were strong enough to cause alewives collapse and prevent their recovery even in Chinook salmon absent scenarios.
The bottom-up controls from nutrients in Lake Huron main basin revealed in our scenario simulations should be partially attributed to dreissenids. Cha and others (2011) showed that nutrient loads from Saginaw Bay to the main basin decreased by 40% after the invasion of zebra mussels around 1990. Thus, dreissenids should be considered partially responsible for the nutrient reduction in the main basin in conjunction with other factors including a series of years of low tributary inflow (Cha and others 2010).
Simulation results suggest that there was an upper limit to Chinook salmon predation on alewives in Lake Huron within the calibration period 1984–2006, and that this limit was expanded with increases in bottom-up controls caused by dreissenid consumption and reduction in nutrients. An upper limit of Chinook salmon predation on alewives probably is determined from limited spatial overlap owing to species’ temperature preferences. In the Lake Huron main basin, Chinook salmon were generally found in areas where temperatures were close to 13°C (Bergstedt, U.S. Geological Survey Hammond Bay Biological Station, Millersburg, Michigan, USA, unpublished data), but alewives were distributed in areas within a wider temperature range as they were found throughout the water column from nearshore to offshore depths of over 110 m (Adlerstein and others 2007).
Increases in Chinook salmon predation on alewives associated with bottom-up control possibly resulted from changes in the biomass proportion of available pelagic prey to Chinook salmon. Alewives and rainbow smelt were the two most important prey for Chinook salmon in Lake Huron (Dobiesz 2003) and simulations showed that rainbow smelt biomass decreased more than alewife biomass under quagga mussel or nutrients scenarios (Figure 6). Therefore, given increases in bottom-up controls that caused decreases in alternative prey, Chinook salmon were probably forced to expand their spatial distribution for feeding on alewives.

Simulated rainbow smelt 1+ biomass under all scenarios of Chinook salmon biomass, Quagga mussel biomass, and nutrient loads. The analysis was designed as a factorial experiment with three factors: Chinook salmon, quagga mussel, and nutrients. Each point represents a 20-year biomass average after reaching equilibrium under a scenario representing effects of one factor or a combination of two or three factors.
Studies have linked Lake Huron alewife collapse to single-factor control by salmonine predation (He and others 2015), dreissenid invasion (Nalepa and others 2007), or reduction in nutrients (Barbiero and others 2012), whereas our analyses showed a possible causality that integrates effects of these factors chronologically. Combined Ecopath analysis and Ecosim simulation results suggest that the alewife collapse in Lake Huron can be described sequentially as: (1) top-down controls surged from increases in Chinook salmon, lake trout, and other salmonine predators, which were initially driven by increases in stocking and then by increases in natural reproduction of Chinook salmon and reduction in sea lamprey mortality in Lake Huron, and then (2) bottom-up controls surged from production reduction caused by decreases in nutrient loads and increases in dreissenids, which caused (3) decreases in the biomass of non-dreissenid benthos, (4) decreases in rainbow smelt biomass, and (5) increases in the proportion of alewife in the diet of Chinook salmon and thus predation mortality, and finally resulted in the collapse of alewife population. The sequence of elements can be used to diagnose onset of alewife collapse in other Great Lakes.
Potential Model Biases
We caution that our conclusions were based on results of Ecopath and Ecosim models developed in this study, which is one of many approaches that could have been used to model the Lake Huron food web. To our knowledge, this is the first and only attempt to use a food web modeling approach to assess top-down and bottom-up controls on the collapse of alewives in Lake Huron. However, future studies may reach different conclusions by using different modeling approaches that can better capture spatiotemporal heterogeneity in species distributions and explicitly simulate temperature-dependent processes.
In addition, our Ecopath models have numerous input parameters, each of which has associated data and estimation uncertainty. Although we did our best to integrate available Lake Huron information, there are still potential biases associated with input parameter estimates in our Ecopath models that may affect results. Among data issues are biomass estimates for food web groups, which were not always representative of annual averages and full distribution ranges. For example, prey fish biomass might be underestimated by surveys conducted in fall after most predation mortality took place (He and others 2015). Further, zooplankton might be overestimated by surveys conducted in summer when primary production was the highest (Munawar and Munawar 1982) and also because samples were collected from stations between 50–140 m in depth, where zooplankton would be less vulnerable to predation by prey fishes that were most abundant in areas with depth less than 80 m (Adlerstein and others 2007). Therefore, our analyses might underestimate top-down controls on zooplankton by prey fishes.
Another potential bias is from calculating absolute biomass from survey index data, mostly for fish groups because there were no gear catchability estimates for most species. Further, biomass data are available in different units across trophic levels, such as carbon weight for phytoplankton, dry weight for zooplankton, and ash-free dry weight for benthos. We made assumptions and selected conversion factors from global databases for conversions into wet weight (see Supplementary Material for details). We are confident that assumptions were appropriate since we balanced both Ecopath models and obtain generally good fits in Ecosim calibration. Nevertheless, more comprehensive estimates for gear catchabilities and biomass conversion factors are needed for improving ecosystem related modeling in the Great Lakes in general.
Lake Huron Results Applicable to Understand Lake Michigan and Lake Ontario dynamics
Our results on Lake Huron food web dynamics under top-down and bottom-up controls are also informative for Lake Michigan and Lake Ontario given they have similar food web structures and were subjected to the same anthropogenic stressors and management as was Lake Huron before alewives collapsed (Adkinson and Morrison 2012; Bunnell 2012). An important message from our findings is that top-down controls on Lake Huron pelagic prey fishes by salmonine predators probably reached an upper limit in the mid-1980s, and would have not caused the alewife collapse without concurrent increases in bottom-up controls. Salmonines control on alewives was limited because their populations did not have a complete spatial overlap given temperature preferences: salmonines prefer 9–15°C and alewives prefer around 20°C (Coutant 1977). An overall upper limit of top-down controls is supported by the work of He and others (2015) showing that between 1988 and 2010 Lake Huron lakewide consumption on pelagic prey by salmonines was relatively constant despite fish community changes. Also Bunnell and others (2014) did not find correlations between predator biomass, mainly Chinook salmon and lake trout, and prey fish biomass in Lake Huron between 1998 and 2010. These results imply that there should also be upper limits of top-down controls on alewives by salmonine predators in Lake Michigan and Lake Ontario, but at different levels.
In Lake Michigan and Ontario, top-down controls by stocked salmonine predators on prey fishes have been widely reported. Before dreissenid invasions in the 1990s, many studies showed that consumptive demands by salmonine predators might exceed the production of prey fishes in Lake Michigan (Stewart and Ibarra 1991) and Lake Ontario (Jones and others 1993; Rand and Stewart 1998). However, there was no major collapse in pelagic prey fish populations in these two lakes. During last decade, top-down controls on pelagic prey fishes by salmonine predators might have been very strong but with bottom-up controls at levels still able to sustain alewife populations. Madenjian and others (2015) reported that Lake Michigan lakewide consumption on pelagic prey fishes by salmonines was relatively constant between 2000 and 2013 despite decreases in prey fish biomass in this period. On the other hand, Bunnell and others (2014) showed prey fish biomass significantly correlated with the biomass of non-dreissenid benthos between 1998 and 2010, which indicates strong bottom-up controls on prey fish. In Lake Ontario, the same study (Bunnell and others 2014) showed a positive correlation between predator biomass (mainly Chinook salmon and lake trout) and prey fish biomass, which indicates weak top-down controls on prey fish. On the other hand, Stewart and Sprules (2011) showed increases in bottom-up controls after dreissenid invasions as there was a synchrony between the decreases in primary production and majority of species biomass in the food web. We suspect that alewife populations in these two lakes will follow the Lake Huron population fate if dreissenids continue to increase and nutrients continue to decline and finally reach levels comparable to those causing the collapse in Lake Huron.
Management Implications
Our results suggest that recovery of alewives in Lake Huron is unlikely regardless of Chinook salmon biomass under current conditions of low nutrients and high quagga mussel biomass, which implies that the shrinking prey base cannot support the same level of salmonine predators as in the 1980s. Our results also have management implications for Lake Michigan and Ontario. Sustaining a prey base for predators that are a target of valuable fisheries in those lakes has been a concern for resource managers ever since the Lake Huron alewife population collapsed. The main difference among these Great Lakes ecosystems is that the Lake Huron main basin is oligotrophic and nutrient loads have been much lower than those in Lake Michigan and Lake Ontario. We cannot directly make predictions for Lake Michigan and Lake Ontario, because in these two lakes Chinook salmon and quagga mussel biomass (Rogers and others 2014; Stewart and Sprules 2011) and nutrient loads (Dolan and Chapra 2012) have been outside the range in our simulations, but we expect that conditions leading to alewife collapses would be similar as observed in Lake Huron. Based on observed food web dynamics (Table 8), it is reasonable to expect that alewives will collapse if consecutive years of low nutrient loads are in the horizon. Currently in both lakes, the biomass flow to Chinook salmon is through a simple pelagic pathway: phytoplankton → zooplankton → alewives → Chinook salmon. When nutrients become limited, both top-down and bottom-up controls on alewives would become very strong and almost certainly result in a population collapse.
References
Adlerstein S, Rutherford E, Riley SC, Haas R, Thomas M, Fielder D, Johnson J, He J, Mohr L. 2007. Evaluation of integrated long-term catch data from routine surveys for fish population assessment in Lake Huron, 1973–2004. Ann Arbor: Great Lakes Fishery Commission Project Completion Report.
Adkinson AC, Morrison BJ, Eds. 2012. The state of Lake Ontario in 2008. Ann Arbor: Great Lakes Fishery Commission Special Publication 14-01.
Barbiero RP, Lesht BM, Warren GJ. 2012. Convergence of trophic state and the lower food web in Lakes Huron, Michigan and Superior. J Gt Lakes Res 38:368–80.
Barbiero RP, Schmude K, Lesht BM, Riseng CM, Warren GJ, Tuchman ML. 2011. Trends in Diporeia populations across the Laurentian Great Lakes, 1997–2009. J Gt Lakes Res 37:9–17.
Beeton AM, Saylor JH. 1995. Limnology of Lake Huron. In: Munawar M, Edsall T, Leach J, Eds. The Lake Huron ecosystem: ecology, fisheries, and management. Amsterdam: SPB Academic Publishing. p 1–37.
Berst AH, Spangler GR. 1972. Lake Huron: Effects of exploitation, introductions, and eutrophication on salmonid community. J Fish Res Board Can 29:877–87.
Borer ET, Seabloom EW, Shurin JB, Anderson KE, Blanchette CA, Broitman B, Cooper SD, Halpern BS. 2005. What determines the strength of a trophic cascade? Ecology 86:528–37.
Bunnell DB, Ed. 2012. The state of Lake Michigan in 2011. Ann Arbor: Great Lakes Fishery Commission Special Publication 12-01.
Bunnell DB, Barbiero RP, Ludsin SA, Madenjian CP, Warren GJ, Dolan DM, Brenden TO, Briland R, Gorman OT, He JX, Johengen TH, Lantry BF, Lesht BM, Nalepa TF, Riley SC, Riseng CM, Treska TJ, Tsehaye I, Walsh MG, Warner DM, Weidel BC. 2014. Changing ecosystem dynamics in the Laurentian Great Lakes: bottom-up and top-down regulation. Bioscience 64:26–39.
Carpenter SR, Kitchell JF, Hodgson JR. 1985. Cascading trophic interactions and lake productivity. Bioscience 35:634–9.
Cha Y, Stow CA, Nalepa TF, Reckhow KH. 2011. Do invasive mussels restrict offshore phosphorus transport in Lake Huron? Environ Sci Technol 45:7226–31.
Cha Y, Stow CA, Reckhow KH, DeMarchi C, Johengen TH. 2010. Phosphorus load estimation in the Saginaw River, MI using a Bayesian hierarchical/multilevel model. Water Res 44:3270–82.
Christensen V, Pauly D. 1992. Ecopath II—a software for balancing steady-state ecosystem models and calculating network characteristics. Ecol Model 61:169–85.
Christensen V, Walters C, Pauly D, Forrest R. 2008. Ecopath with Ecosim version 6: user guide. Lenfest Ocean Futures Project.
Christensen V, Walters CJ. 2004. Ecopath with Ecosim: methods, capabilities and limitations. Ecol Model 172:109–39.
Claramunt RM, Warner DM, Madenjian CP, Treska TJ, Hanson D. 2013. Offshore salmonine food web. In: Bunnell DB, Ed. The State of Lake Michigan in 2011. Ann Arbor: Great Lakes Fishery Commission Special Publication 12-01. p 13–23.
Connerton MJ, Lantry JR, Walsh MG, Daniels ME, Hoyle JA, Bowlby JN, Johnson JH, Bishop DL, Schaner T. 2014. Offshore pelagic fish community. In: Adkinson AC, Morrison BJ, Eds. The State of Lake Ontario in 2008. Ann Arbor: Great Lakes Fishery Commission Special Publication 14-01. p 42–59.
Coutant CC. 1977. Compilation of temperature preference data. J Fish Res Board Can 34:739–45.
Dettmers JM, Goddard CI, Smith KD. 2012. Management of alewife using Pacific salmon in the Great Lakes: Whether to manage for economics or the ecosystem? Fisheries 37:495–501.
Dobiesz NE. 2003. An evaluation of the role of top piscivores in the fish community of the main basin of Lake Huron. Doctoral dissertation, Michigan State University.
Dolan DM, Chapra SC. 2012. Great Lakes total phosphorus revisited: 1. Loading analysis and update (1994–2008). J Great Lakes Res 38:730–40.
Ebener MP, Johnson JE, Reid DM, Payne NP, Argyle RL, Wright GM, Krueger K, Baker JP, Morse T, Weise J. 1995. Status and future of Lake Huron fish communities. In: Munawar M, Edsall T, Leach J, Eds. The Lake Huron Ecosystem: Ecology, Fisheries, and Management. Amsterdam: SPB Academic Publishing. p 125–69.
Finn JT. 1976. Measures of ecosystem structure and function derived from analysis of flows. J Theor Biol 56:363–80.
Gaden M, Goddard C, Read J. 2012. Multi-jurisdictional management of the shared Great Lakes fishery: transcending conflict and diffuse political authority. In: Taylor WW, Lynch AJ, Leonard NJ, Eds. Great Lakes Fisheries Policy and Management: a binational perspective. 2nd edn. East Lansing: Michigan State University Press. p 305–37.
He JX, Bence JR, Madenjian CP, Pothoven SA, Dobiesz NE, Fielder DG, Johnson JE, Ebener MP, Cottrill RA, Mohr LC, Koproski SR. 2015. Coupling age-structured stock assessment and fish bioenergetics models: A system of time-varying models for quantifying piscivory patterns during the rapid trophic shift in the main basin of Lake Huron. Can J Fish Aquat Sci 72:7–23.
Hecky RE, Smith REH, Barton DR, Guildford SJ, Taylor WD, Charlton MN, Howell T. 2004. The nearshore phosphorus shunt: A consequence of ecosystem engineering by dreissenids in the Laurentian Great Lakes. Can J Fish Aquat Sci 61:1285–93.
Higgins SN, Vander Zanden MJ. 2010. What a difference a species makes: a meta-analysis of dreissenid mussel impacts on freshwater ecosystems. Ecol Monogr 80:179–96.
Jacobs GR, Madenjian CP, Bunnell DB, Warner DM, Claramunt RM. 2013. Chinook salmon foraging patterns in a changing Lake Michigan. Trans Am Fish Soc 142:362–72.
Johnson JH, Ross RM, McCullough RD, Mathers A. 2010. Diet shift of double-crested cormorants in eastern Lake Ontario associated with the expansion of the invasive round goby. J Great Lakes Res 36:242–7.
Jones ML, Koonce JF, Ogorman R. 1993. Sustainability of hatchery-dependent salmonine fisheries in Lake Ontario: the conflict between predator demand and predator supply. Trans Am Fish Soc 122:1002–18.
Jørgensen SE, Tundisi JG, Matsumura-Tundisi T. 2012. Handbook of inland aquatic ecosystem management. London: CRC Press.
Kao YC, Adlerstein SA, Rutherford ES. 2014. The relative impacts of nutrient loads and invasive species on a Great Lakes food web: an Ecopath with Ecosim analysis. J Great Lakes Res 40:35–52.
Madenjian CP, Bunnell DB, Warner DM, Pothoven SA, Fahnenstiel GL, Nalepa TF, Vanderploeg HA, Tsehaye I, Claramunt RM, Clark RD Jr. 2015. Changes in the Lake Michigan food web following dreissenid mussel invasions: A synthesis. J Gt Lakes Res 41:217–31.
Madenjian CP, O’Gorman R, Bunnell DB, Argyle RL, Roseman EF, Warner DM, Stockwell JD, Stapanian MA. 2008. Adverse effects of alewives on Laurentian Great Lakes fish communities. North Am J Fish Manag 28:263–82.
McQueen DJ, Johannes MRS, Post JR, Stewart TJ, Lean DRS. 1989. Bottom-up and top-down impacts on freshwater pelagic community structure. Ecol Monogr 59:289–309.
McQueen DJ, Post JR, Mills EL. 1986. Trophic relationships in freshwater pelagic ecosystems. Can J Fish Aquat Sci 43:1571–81.
Miller RR. 1957. Origin and dispersal of the alewife, Alosa pseudoharengus, and the gizzard shad, Dorosoma cepedianum, in the Great Lakes. Trans Am Fish Soc 86:97–111.
Munawar M, Munawar IF. 1982. Phycological studies in Lakes Ontario, Erie, Huron, and Superior. Can J Bot 60:1837–58.
Nalepa TF, Fanslow DL, Lang GA. 2009. Transformation of the offshore benthic community in Lake Michigan: Recent shift from the native amphipod Diporeia spp. to the invasive mussel Dreissena rostriformis bugensis. Freshw Biol 54:466–79.
Nalepa TF, Fanslow DL, Pothoven SA, Foley AJIII, Lang GA. 2007. Long-term trends in benthic macroinvertebrate populations in Lake Huron over the past four decades. J Great Lakes Res 33:421–36.
Pauly D, Christensen V, Guenette S, Pitcher TJ, Sumaila UR, Walters CJ, Watson R, Zeller D. 2002. Towards sustainability in world fisheries. Nature 418:689–95.
Pothoven SA, Madenjian CP. 2013. Increased piscivory by lake whitefish in Lake Huron. N Am J Fish Manag 33:1194–202.
Pothoven SA, Nalepa TF. 2006. Feeding ecology of lake whitefish in Lake Huron. J Gt Lakes Res 32:489–501.
Rand PS, Stewart DJ. 1998. Prey fish exploitation, salmonine production, and pelagic food web efficiency in Lake Ontario. Can J Fish Aquat Sci 55:318–27.
Ridgway MS, Fielder DG. 2012. Double-crested cormorants in the Laurentian Great Lakes: issues and ecosystems. In: Taylor WW, Lynch AJ, Leonard NJ, Eds. Great Lakes fisheries policy and management: a binational perspective. 2nd edn. East Lansing: Michigan State University Press. p 733–64.
Riley SC, Roseman EF, Nichols SJ, O’Brien TP, Kiley CS, Schaeffer JS. 2008. Deepwater demersal fish community collapse in Lake Huron. Trans Am Fish Soc 137:1879–90.
Rogers MW, Bunnell DB, Madenjian CP, Warner DM. 2014. Lake Michigan offshore ecosystem structure and food web changes from 1987 to 2008. Can J Fish Aquat Sci 71:1072–86.
Shapiro J, Wright DI. 1984. Lake restoration by biomanipulation: Round Lake, Minnesota, the first 2 years. Freshw Biol 14:371–83.
Smith BR, Tibbles JJ. 1980. Sea Lamprey (Petromyzon marinus) in Lakes Huron, Michigan, and Superior: history of invasion and control, 1936–78. Can J Fish Aquat Sci 37:1780–801.
Smith VH. 2003. Eutrophication of freshwater and coastal marine ecosystems: a global problem. Environ Sci Pollut Res 10:126–39.
Stewart DJ, Ibarra M. 1991. Predation and production by salmonine fishes in Lake Michigan, 1978–88. Can J Fish Aquat Sci 48:909–22.
Stewart TJ, Sprules WG. 2011. Carbon-based balanced trophic structure and flows in the offshore Lake Ontario food web before (1987–1991) and after (2001–2005) invasion-induced ecosystem change. Ecol Model 222:692–708.
Su ZM, He JX. 2013. Analysis of Lake Huron recreational fisheries data using models dealing with excessive zeros. Fish Res 148:81–9.
Tody WH, Tanner HA. 1966. Coho salmon for the Great Lakes. Lansing: Michigan Department of Conservation Fish Division Fish Management Report No. p 1.
Vanderploeg HA, Nalepa TF, Jude DJ, Mills EL, Holeck KT, Liebig JR, Grigorovich IA, Ojaveer H. 2002. Dispersal and emerging ecological impacts of Ponto-Caspian species in the Laurentian Great Lakes. Can J Fish Aquat Sci 59:1209–28.
Walters C, Christensen V, Pauly D. 1997. Structuring dynamic models of exploited ecosystems from trophic mass-balance assessments. Rev Fish Biol Fish 7:139–72.
Weidel BC, Connerton MJ. 2012. Status of rainbow smelt in the U.S. waters of Lake Ontario, 2011. Oswego: U.S. Geological Survey Great Lakes Science Center Lake Ontario Biological Station.
Wilson KA, Howell ET, Jackson DA. 2006. Replacement of zebra mussels by quagga mussels in the Canadian nearshore of Lake Ontario: The importance of substrate, round goby abundance, and upwelling frequency. J Gt Lakes Res 32:11–28.
Acknowledgements
We are thankful to Adam Cottrill, Jixiang He, Thomas Nalepa, Stephen Riley, Jeffrey Schaeffer, and Zhenming Su for providing data and helping with data issues. We are also thankful to Jason Breck and Hongyan Zhang for programming and EwE modeling support. James Diana, Charles Madenjian, Earl Werner, Michael Wiley, David “Bo” Bunnell, and Randall Claramunt provided suggestions to an early draft of this article. This research was funded by award number DW-13-92359501 from the U.S. Environmental Protection Agency Great Lakes Restoration Initiative and award numbers GL-00E00604-0 and NA10NOS4780218 from the National Oceanic and Atmospheric Administration (NOAA) Center for Sponsored Coastal Ocean Research. This manuscript is NOAA GLERL Contribution No. 1801.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
All authors together designed the study, performed the research, and wrote the paper; YCK and SAA analyzed data.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Kao, YC., Adlerstein, S.A. & Rutherford, E.S. Assessment of Top-Down and Bottom-Up Controls on the Collapse of Alewives (Alosa pseudoharengus) in Lake Huron. Ecosystems 19, 803–831 (2016). https://doi.org/10.1007/s10021-016-9969-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-016-9969-y