
Abstract
Eradications of kiore or Pacific rats (Rattus exulans) from islands around New Zealand have been followed by responses from resident species of coastal plants, invertebrates, reptiles and seabirds. These responses are compared with an invasion by ship rats (Rattus rattus), which devastated populations of invertebrates, birds and bats. Post-eradication responses only approximate the effects of invasions because recovery is limited to the residual pool of native species. Greater effects of kiore are indicated by adding incompatible species confined to rat-free locations. The extended list includes at least 15 species of invertebrates, two species of frogs, tuatara (Sphenodon punctatus), 11 species of lizards and 9 species of seabirds. The analyses indicate direct and indirect effects of kiore similar to those reported after ship rat invasions. This is despite indications from the literature that kiore are the least damaging of the three commensal rat species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Invasive species are those exotic organisms that establish in new locations and threaten native biodiversity, ecosystem functioning, animal and plant health, and human economies (Myers et al. 2000). Such species have been particularly damaging on islands, where two thirds of recorded animal extinctions have been attributed to introduced species (Cole et al. 2005). Rats are quintessential invaders that have been spread to at least 80% of the world’s island groups (Atkinson 1985), but information about their effects is surprisingly weak, patchy, geographically scattered and biased towards vertebrates, especially birds (Towns et al. 2006).
Differences between the fauna of islands invaded by rats and those that are rat free were noted in New Zealand early in the twentieth century. The naturalist Herbert Guthrie-Smith lamented differences between the birdlife of the rat-free islands in the Muttonbird (Titi) Islands in 1913, and those elsewhere that had been invaded by rats introduced from Europe (Guthrie-Smith 1925). More commonly, the link between rats and declines of indigenous biota was never made. Had the link been known, Joseph Banks would perhaps have been less prosaic in his description of the song of bellbirds (Anthornis melanura) at Ship Cove at the northern end of the South Island, New Zealand. On the morning of 17 January 1770 he wrote from the ship Endeavour: “…the numbers of them were certainly very great who seemed to strain their throats with emulation perhaps; their voices were certainly the most melodius wild musick I have ever heard, almost imitating small bells but with the most tuneable silver sound imaginable…” (Begg and Begg 1969). The day previously, expedition leader James Cook had careened Endeavour. The ship was aground or moored to the shore for 3 weeks (Reed and Reed 1969) providing a perfect opportunity for some of its cargo of “vermin” or Norway rats (Rattus norvegicus) to jump ship. The loss of “wild musick” may well have begun at that point. Subsequently, for 3 weeks in March 1773, Cook tied the Resolution so close to shore in the south-western South Island, that “the vermin could walk ashore over the bridge” (Beaglehole 1961). If the first invasion failed, the second most likely succeeded.
The impacts of these early invasions remained unclear because the locations visited by early explorers were already inhabited by kiore or Pacific rats (Rattus exulans). These south-east Asian rats were spread by Polynesian voyagers throughout the Pacific and had reached New Zealand at least 500 years before European contact (Atkinson and Towns 2005; Wilmshurst et al. 2008). Furthermore, with European settlement, mice (Mus musculus), ship rats (Rattus rattus), mustelids, domestic cats and other mobile predators were added to the main islands. It was not until ship rats invaded Big South Cape Island off southern New Zealand that the devastating effects of rats on some naive indigenous species were demonstrated (Atkinson and Bell 1973; Ballance 2007).
Invasions are uncontrolled experiments, and because of their unpredictable occurrence, are rarely documented in detail. As an alternative to using invasions, the impacts of rats may be revealed when rats are removed and the response of resident species is measured (Campbell et al. 1984; Veltman 1996). Since 1960, three species of rats have been removed at least once from about 80 islands around New Zealand and on 34 of these rats were the only introduced mammal (Towns and Broome 2003; Clout and Russell 2006). Here, I review the information gained about the invasive effects of rats from these eradications.
I focus on kiore, partly because the history of eradications is longest for this species, but also because its effects have been intensively studied in New Zealand. Kiore are widespread elsewhere in the Pacific, but I have been unable to find published accounts of post eradication responses by native species in the Pacific islands.
To answer whether rat eradications are in effect reversed invasions, it is necessary first to describe a rat invasion. There are few accounts of the effects of a kiore invasion into complex island systems. The one exception is the invasion of 1 ha Lizard Island in 1977, but the kiore were eradicated within 12 months (McCallum 1986). I therefore begin with the ship rat invasion of the Big South Cape archipelago, because it provides insights into likely general effects of rat invasions (e.g. Courchamp and Caut 2005). I then illustrate the range of effects of kiore demonstrated after eradications using selected island case studies and discuss the validity of using eradications as a means of determining the effects of invasions.
The invasion of Big South Cape and neighbouring islands
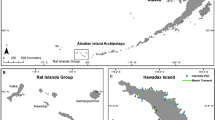
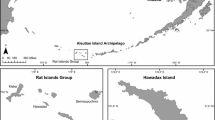
Big South Cape Island (Taukihepa)—hereafter referred to as Big South Cape Island—is the largest of the Muttonbird (Titi) Islands which are privately owned and used for traditional harvesting of sooty shearwaters (Puffinus griseus). The islands, which are off southwestern Stewart Island/Rakiura (Fig. 1), include Big South Cape Island (939 ha), Rerewhakaupopo (Solomon) Island (26 ha) and Pukeweka Island (1.5 ha). In about 1962 ship rats invaded the three islands, presumably over stern lines from fishing boats (Atkinson and Bell 1973; Bell 1978; Atkinson 1989) which, based on analyses of mtDNA, were carrying rats from outside the Stewart Island archipelago (R. Fewster, pers. comm., 2006). Following the invasion, two groups of plants showed the effects of severe browsing, some to ground level; one species of weevil was extirpated, flightless crickets were all but eliminated and nine species of birds and both resident species of bats were either greatly reduced in numbers or extirpated. Of the extirpated birds and bats, one species of bird and bat were total extinctions, as were two subspecies of bird (Table 1). There was no immediate evidence of detrimental effects on burrowing seabirds (Bell 1978), and some native passerines including tui (Prosthemadera novaeseelandiae), silvereyes (Zosterops lateralis), grey warblers (Gerygone igata), tomtits (Petroica m. macrocephala) and at least three species of introduced passerines appeared to be more abundant after the invasion (Atkinson and Bell 1973; Bell 1978).

Locations in New Zealand mentioned in the text
The invasion has many implications. First, the spread of rats and loss of sensitive species was extremely rapid. The time between the earliest recorded rat presence in August 1962 and the peak of the irruption in February 1965, when all 939 ha of the island were occupied, was only 2.5 years (Atkinson and Bell 1973). Within 5 years, five species of birds were extirpated or functionally so (reduced to one sex) and three had shown substantial declines.
Second, damage to some native species was greatest during the invasion phase, which implies that the spreading rats overshot their food supply. For example, megaherbs were initially gnawed to near ground level, but were recovering within 5 years (Atkinson and Bell 1973).
Third, the recorded effects were wide-ranging and included plants, invertebrates, birds and mammals. However, with the exception of bats, the effects were also selective within each group. Large flightless invertebrates were heavily affected. Furthermore, ten of the eleven species of affected vertebrates nest or roost either on or near the ground and in tree, ground and rock cavities. All of them were within or less than the body weight (100–180 g) of ship rats (Innes 2005). Other species appeared unaffected and some apparently became more abundant.
Fourth, the effects were apparently direct and indirect. The loss of the large flightless weevil may have been direct through predation and indirect due to loss of its host plant during the period of extensive browsing. However, the increase of some species of birds is likely to have been an indirect response to the loss of others. This implies that invasions can have unpredictable knock-on effects.
Fifth, for most species the mechanism for decline is unknown. However, at least some species were directly affected by predation of roosting females, as revealed by a sex bias to males in the last of the surviving saddlebacks (Blackburn 1965). Subsequent video camera studies indicate that ship rats frequently feed on eggs and chicks in the nest (Brown 1997).
Finally, although heralded as the best account of an invasion in New Zealand, there are complicating effects. A native flightless rail, the Stewart Island weka (Gallirallus australis scotti) is present on Big South Cape Island. Weka were introduced to many southern islands (probably including Big South Cape), most likely as an alternative food supply for mutton-birders (Atkinson and Bell 1973). Weka are omnivores that eat fruit, carrion, invertebrates, lizards, eggs, chicks and adults of burrowing seabirds and ground-dwelling terrestrial birds (Beauchamp et al. 1999). Releases of weka elsewhere onto islands in the Stewart Island archipelago were followed by declines of fairy prions (Pachyptila turtur), diving petrels (Pelecanoides urinatrix), mottled petrels (Pterodroma inexpectata) and Cook’s petrels (Pterodroma cookii) (Atkinson and Bell 1973; Imber et al. 2003). The first three of these species were present on Big South Cape Island (Blackburn 1965). Weka were reported on Solomon Island in 1913, as were banded rails (Guthrie-Smith 1925, p. 118). However, Bell (1978) considered weka to be responsible for the later absence of banded rails from Rerewhakaupopo Island and the restricted range of wren and snipe on Big South Cape Island. Recent analyses of δC13 isotope signatures in rats and weka on these islands indicate a stronger marine component in the diet of weka than for ship rats. This evidence, and the experimental local removal of rats and weka, indicates that weka feed more frequently on seabirds than do rats, whereas ship rats feed more frequently on terrestrial passerines than do weka (Harper 2007). Comments by Bell (1978) that seabirds did not appear to have been detrimentally affected by the ship rat invasion are therefore probably correct, because weka may have already severely depleted the more vulnerable species. However, there are local reports of the loss of prions and diving petrels since the rat invasion (G. Harper, pers. comm., 2006). Whether there were additive or synergistic effects between weka and ship rats is unknown but cannot be discounted.
Such complications bedevil most studies of the effects of invasive rodents, including those based on responses after rats were eradicated. Accordingly, most of the examples used below are from locations where the effects of kiore are unlikely to be complicated by synergistic effects of other introduced predatory birds or mammals.
The eradication of rats from islands provides an opportunity to investigate three of the above effects indicated for Big South Cape and neighbouring islands: the range and life history of the species most likely to be affected; the extent to which the effects are direct or indirect; and, at least for some species, the likely mechanisms for decline. Below, I use four sets of examples to illustrate these for kiore.
Selective and successional effects of kiore on plants
The possible effects of kiore on island-inhabiting plants were reviewed by Atkinson (1986), who identified the nikau palm and the trees karo and milktree (Table 2) as vulnerable to the effects of kiore based on distributional evidence. This hypothesis was tested when kiore were removed from several islands off northeastern New Zealand. Increased seedling recruitment in the absence of kiore confirmed the hypotheses for all three species, but revealed that the effects had been under-estimated. In fact, at least 11 plant species showed significant increases in seedling recruitment when kiore were removed (Table 2), when compared with locations still with kiore and those never invaded. Additional species with large fruit showed patchy responses or tended to be most common on the larger islands, thereby restricting the range of sites for comparison. For example, karaka (Corynocarpus laevigatus) showed consistent seedling responses on some islands following removal of kiore but gave conflicting data on others (Campbell and Atkinson 1999). Furthermore, some of the effects of kiore were underestimated for resident populations. For example, Campbell and Atkinson (2002) found dioecious species that showed hitherto unrecognised sex imbalances, species that survived only where seeds lodged in rock falls, and species that were propagating vegetatively by layering.
The species of plants suppressed by kiore have a range of fruit sizes and reproductive modes. Studies of seedling recruitment imply that kiore are seed predators. However, impaired seedling recruitment may also reflect destruction of flowers, flower buds or developing fruit on the tree (Campbell and Atkinson 1999). Increased recruitment of seedlings after kiore are removed therefore represents the sum of these effects. For example, of two species with large fruit (Table 2), tawapou appears heavily affected by kiore, which eat the flesh and seeds, whereas kohekohe is less obviously affected, probably because kiore often only eat the fruit (Campbell and Atkinson 1999). Nonetheless, the strong response by kohekohe after kiore are removed suggests other effects of kiore, perhaps on the flowers, which are formed into large panicles that protrude from the trunk.
At least 6 additional species of plants have been identified as sensitive to kiore using experimental cages and exclosures (Campbell and Atkinson 2002; Towns et al. 2006), but whether they similarly respond when kiore are removed is yet to be determined.
Many of the species detrimentally affected by kiore produce copious fruit and four of the 11 species in Table 2 produce very large fruit. Some of these are favoured food species of the native fruit pigeon or kereru (Hemiphaga novaeseelandiae), which is the only species capable of transporting the ingested seeds in their gut. Furthermore, trees such as kohekohe, tawapou and milktree are canopy species which, on some islands without rats, dominate the mixed coastal forest. Campbell and Atkinson (2002) suggest that historic forest clearance on islands, coupled with the presence of kiore, has limited the recruitment pool of species into coastal forest. Without kiore, kereru are attracted to numerous fruiting trees, the forest may become dominated by species vulnerable to kiore, and these often bear the large fruit dispersed by kereru. Where kiore are present, kereru are less likely to visit the few fruiting trees, the forest is more likely to be dominated by species resistant to kiore, and species vulnerable to rats become rare or locally extinct. Such effects can be modified by site differences but, overall, the recovery of a wide range of species after removal of kiore indicates that, while present, these rats can fundamentally change successional pathways (Campbell and Atkinson 2002).
Indirect effects of kiore on an invertebrate
The honey dew scale insect Coelostomidia zealandica (Hemiptera: Margarodidae) inhabits northeastern islands. Immature scale insects or crawlers settle in cracks or on the branches of its host plants, which include karo and ngaio (Myoporum laetum). The larvae insert their mouthparts and produce a long tube to exude sugary waste. Infested trees have distinctive black trunks caused by sooty moulds growing on the secretions. The honeydew is highly sought after by nectivorous birds and geckos (Towns 2002a) and can be a major energy source in some forest ecosystems (Beggs 2001).
Since karo often becomes rare on islands inhabited by kiore (Campbell and Atkinson 1999), the scale insects decline as well. Invaded islands usually lack honey dew scale on the remaining karo but also ngaio, even though the latter species does not appear to be negatively affected by kiore. Towns (2002a) found that the scale insects occasionally recover and slowly reinfest karo and ngaio trees after rats (and other browsing mammals) are removed. Karo appears to be a vital component of the life cycle of the scale insects because it is a shade-tolerant species. Without karo, ngaio that die lose their scale populations and new plants are not reinfested because there is no mechanism for transfer between the scattered light-hungry ngaio trees. Furthermore, because female scale insects are flightless, the rate of re-infestation is very slow, and may cease altogether if source populations are lost. Consequently, honeydew scale in some archipelagos can be confined to tiny rat-free islets, and be absent from their larger, forested but kiore-inhabited neighbours (Towns 2002a). In this example, loss of one host species has resulted in the collapse of a parasite transfer pathway that, in turn, can destroy an energy transfer source.
More direct effects of kiore have been reported for three species each of terrestrial snails, cockroaches, flightless crickets and beetles, a species of earwig and a species of large spider (Table 3).
Direct and indirect effects of kiore on a large reptile
The endemic tuatara (Sphenodon punctatus) is a large iguana-like sphenodontid reptile that can weigh up to 1,300 g (Gill and Whitaker 1996). By comparison, kiore are unusually large if they reach 120 g (Atkinson and Towns 2005). Tuatara were unknown to eighteenth century European explorers, when kiore were the only rats on the mainland, and were not revealed until about 1840 when populations were discovered on offshore islands (Robb 1986). Nonetheless, that tuatara had previously been present on the mainland is demonstrated by extensive Holocene deposits throughout the North and South Islands (Worthy and Holdaway 2002). Today, tuatara have a disjunctive distribution on islands off the northeastern North Island and in Cook Strait between the North and South Islands (Gaze 2001).
The potential role of kiore in the range contraction of tuatara was identified by Crook (1973) and Whitaker (1978). They found lower densities of tuatara and populations with little juvenile recruitment on islands where kiore were present. These findings were confirmed by Cree et al. (1995), who concluded that juvenile recruitment was impaired on 8 of the 9 islands where kiore and tuatara co-existed and, of these, the tuatara populations were near extinction on five. Cree et al. (1995) proposed possible direct and indirect effects of kiore. Direct effects were likely to be predation of eggs and young, which would explain low or failed recruitment. Indirect effects could include competition for food, as a result of dietary overlap between the two species (e.g. Ussher 1999).
Removal of kiore was proposed as one way to test hypotheses about their effects on tuatara (Cree et al. 1995; Veltman 1996), with the appearance of hitherto missing cohorts of juvenile tuatara as the predicted outcome. These predictions were tested on three islands in the Marotere group, with neighbouring Taranga Island retaining kiore as control (Towns et al. 2007). The removal of kiore was followed by significant increases in juvenile recruitment on the three treatment islands. However, the proportional increase varied by island and by sites within islands, indicating location-specific responses. These may be a response to localised tuatara nesting sites from which juveniles gradually disperse to other locations.
An unpredicted response was increased body condition of tuatara on two of the three islands after kiore were removed. This indicated location-specific indirect effects of kiore, either through interference competition or competition for food.
Other species of herpetofauna with detrimental effects attributed to kiore include two species of frogs (now extinct), three species of geckos, and seven species of nocturnal or presumed nocturnal skinks (one now extinct). Two species of diurnal skinks have showed variable responses to kiore removal, with measurable effects at some (but not all) sites (Table 4).
Effects of kiore on ecosystem drivers
Coastal ecosystems around New Zealand were almost certainly influenced by the nutrients imported and activities of vast numbers of burrowing seabirds now largely confined to offshore islands (Worthy and Holdaway 2002). On islands, seabirds can form extremely dense colonies with burrow of densities at least 1/m2. As a result, soil geochemical properties can be strongly modified, with low pH but high nutrient concentrations (Mulder and Keall 2001). The burrows are also used as retreats for lizards and tuatara (e.g. Newman 1987). Burrowing seabirds are therefore viewed as ecosystem drivers because their activity has a multitude of direct and indirect effects. These effects can be measured in below-ground food webs, above-ground vegetation composition and structure (Fukami et al. 2006; Mulder et al. 2008), and in biomass of other dependent groups such as lizards and tuatara (Markwell and Daugherty 2002; Towns 2002b).
Following the eradication of kiore from islands, at least four species of burrowing seabirds <300 g in weight have shown positive responses either through increased fledging success or increased colony size (Table 5). One such example is Cook’s petrel (Pterodroma cookii). Only two island populations of this species remain although evidence indicates that it was previously widespread on the mainland (Imber et al. 2003). The smallest population is on Whenuahou (Codfish) Island, off Stewart Island/Rakiura where, over the 10 years before 1998, there was periodic heavy predation of chicks by kiore at a ridge-top site. The percentage of chicks successfully fledging at this site ranged from 8 to 69%. However, when kiore were removed in 1998, fledging success increased from 8% (immediately before the eradication) to 54% the following breeding season, and by 2003 reached 88%. The control for this was the second population on Hauturu (Little Barrier Island) which, where predation by kiore was high, had fledging success ≤22% (Imber et al. 2003). Immediately before kiore were removed from Hauturu in 2004, fledging success was as low as 5%, but this increased to an average 60% when kiore were removed (Rayner et al. 2007).
The effects of such declines following invasions by kiore were summarised by Holdaway (1999). At least 10 species of seabirds have become locally or totally extinct in New Zealand following invasions by kiore, and most of them were less than 200 g and with an egg length ≤60 mm. In addition, at least 13 species or subspecies of terrestrial birds declined to extinction soon after the arrival of kiore in New Zealand (Tennyson and Martinson 2006).
Discussion
Rats have been eradicated from New Zealand islands at the behest of government conservation agencies in order to: (i) develop new eradication technologies (Towns and Broome 2003); (ii) protect resident species under threat, such as tuatara (see earlier); (iii) enable the ex situ management of threatened species such as the flightless parrot or kakapo Strigops habroptilus (Elliot et al. 2001); (iv) enable recolonisation by extirpated species (natural or assisted); and to (v) increase the ecological integrity of nature reserves penetrated by exotic species (Reserves Act 1977). None of the eradications were undertaken as experiments to measure the effects of rats on native species, which is understandable given the expense and infrastructure needed as support (Cromarty et al. 2002; Towns and Broome 2003). Nonetheless, considerable data have been gathered as species respond to rat removals, and these provide a useful basis for comparison with invasions.
Post eradication responses confirmed that the effects of rats can be direct or indirect. In response to direct effects, there were rapid responses such as increased fledging success of seabirds, and the appearance of new cohorts of juveniles, such as those measured for tuatara. By comparison, indirect effects can be difficult to detect and may involve long term and subtle responses. The impacts of kiore on karo trees and suppression of the plants as hosts of honeydew scale is such an example.
However, the effects identified after eradications of kiore only partly reflect those revealed from the ship rat invasion of Big South Cape and neighbouring islands; eradications should be seen as invasions in reverse only within limits. The first problem is that invasion can result in local extirpation or total extinctions, which cannot be revealed from responses to rodent eradications. Many extirpated species cannot recolonise and it becomes necessary to use circumstantial evidence within archipelagos to indicate the likely effects of rats while present. For example, if invertebrates apparently incompatible with kiore are included in the list of vulnerable species, the number increases from 11 to 15, all of which are ground-dwelling, nocturnal and have body lengths >12 mm. Furthermore, extinctions or dramatic range declines that coincide with the arrival of kiore on the main islands of New Zealand are reported for an additional six species of large invertebrates (Gibbs 2008). Consequently, the total number of invertebrates incompatible with kiore as revealed from eradications is only 52% of the total revealed from total or local extinctions.
In addition to invertebrates, lists that include detrimentally affected and incompatible species of vertebrates include two species of frogs, tuatara, and 11 species of lizards. Almost all of these species are nocturnal and the tuatara and nine species of affected skinks are ground dwelling. The latter include six species of Cyclodina, representing all of the larger members of the genus. The avifauna includes nine species of small-bodied seabirds of less than 300 g. However, kiore were identified as the most likely cause of extinction for one additional species of seabird and 13 species of terrestrial birds from the New Zealand archipelago (Holdaway 1999; Tennyson and Martinson 2006). Whether rats were the sole cause either directly or indirectly is difficult to assess for all of these species because some, such as seabirds and ducks, may also have been harvested by people. Furthermore, predation by feral dogs cannot be discounted as a contributing factor (Towns and Daugherty 1994), something dismissed by others (e.g. Tennyson and Martinson 2006).
A second problem is that resident species need to have residual populations that are sufficiently large and reproductively viable for responses to be measured when rats are eradicated. For example, plants may be present but represented by reproductively constrained populations (Campbell and Atkinson 2002), in which case, a lack of a response may not necessarily indicate a lack of sensitivity to rats.
How ever they are revealed, the effects of kiore discussed to this point are largely at species level. Furthermore, published records of vertebrate declines indicate that kiore are probably the least-damaging of the three commensal rat species (Towns et al. 2006). Nonetheless, when plants and invertebrates are added to vertebrates, the effects of kiore can likely accumulate into pervasive impacts on whole ecosystems. On seabird islands around New Zealand, kiore have been demonstrated to directly affect all four components of the islands’ unique seabird-reptile-invertebrate-plant communities (Fig. 2). These effects are probably greatest in coastal forest where there are three pathways of interaction: direct predation of seeds, seedlings and flowers; indirect effects through suppression of seabirds; and the modification of successional pathways. Invertebrates may also be affected directly and indirectly. Indirect effects may stem from interruption of host-parasite pathways, but also, presumably, through modification of forest vegetation. Because of the loss of honeydew sources, geckos can also be indirectly affected by kiore. The fact that tuatara showed shifts in body condition on some islands after removal of kiore indicates that there are site-specific indirect effects on these species as well. Finally, since burrowing seabirds provide refuges and food for tuatara and lizards, the loss of seabirds must inevitably affect reptiles. Although for most species of lizards, the direct effects of kiore on reptiles are probably sufficiently strong to mask most indirect effects through the loss of seabirds. Therefore, it would not be surprising if kiore have effects on marine derived nutrients and below ground organisms similar to those described on islands invaded by ship and Norway rats (Fukami et al. 2006).

Diagrammatic representation of the effects of kiore on islands around New Zealand dominated by small seabirds. Based on examples from post-eradication recovery, with solid lines showing direct effects and broken lines showing indirect effects. Additional data for affected species and those with exclusive ranges are given in Towns et al. (2006)
It is unclear to what extent lessons from temperate New Zealand can be transferred to kiore populations in the tropical Pacific. Suggestions that kiore were responsible for major forest transformations in the Hawaiian islands (Athen et al. 2002) and loss of palms from Easter Island (Hunt 2007) at present have little contemporary data in support from the region. There is some evidence that at least four species of seabirds responded to kiore eradication from Kure Atoll (Alton 2000), and there are anecdotal reports of rapid responses of lizards after recent eradications of kiore in Fiji (C. Morley and R. Chappell, pers. comm., 2007). Much could be revealed about the past effects of kiore from intensive studies at such sites.
The final lesson from New Zealand eradications is that the interactive and indirect effects of rats can be exceedingly complex, are very difficult to disentangle, and as a result, may be underestimated. This does not mean that our lessons from eradications are of dubious value. It may mean that the hypotheses and models proposed have to be quite general and perhaps site-specific (e.g. Towns and Atkinson 2004). It also means that we will need to be patient and prepared for the unpredicted. Of course, if these long-running experiments are to be of use, they will need regular and long-term measurement. Most of all, they will need constant vigilance to ensure that rats do not recolonise.
References
Alton H (2000) Nature’s sovereignty rules Northwestern Hawaiian isles. Honolulu Star Bulletin, 14 October 2000
Athen JS, Tuggle HD, Ward JV, Welch DJ (2002) Avifaunal extinctions, vegetation change, and Polynesian impacts in prehistoric Hawai’i. Archaeol Ocean 37:57–78
Atkinson IAE (1985) The spread of commensal species of Rattus to oceanic islands and their effects on island avifaunas. In: Moors PJ (ed) Conservation of island birds. ICBP Technical Publication No. 3, pp. 35–81
Atkinson IAE (1986) Rodents on New Zealand’s northern offshore islands: distribution, effects and precautions against further spread. In: Wright AE, Beever RE (eds) The offshore islands of northern New Zealand. New Zealand Department of Lands and Survey Information Series 16, Wellington, pp 13–40
Atkinson IAE (1989) Introduced animals and extinctions. In: Western DC, Pearl MC (eds) Conservation for the twenty-first century. Oxford University Press, New York, pp 54–75
Atkinson IAE, Bell BD (1973) Offshore and outlying islands. In: Williams GR (ed) The natural history of New Zealand. A.H. and A.W. Reed, Wellington, pp 372–392
Atkinson IAE, Towns DR (2005) Kiore. In: King CM (ed) The handbook of New Zealand mammals, 2nd edn. Oxford University Press, Melbourne, pp 159–174
Ballance A (2007) Don Merton the man who saved the black robin. Reed, Auckland
Beaglehole JC (ed) (1961) The journals of Captain James Cook on his voyages of discovery. The voyage of resolution and adventure, vol 2. Cambridge University Press for the Hakluyt Society, Cambridge, pp 1772–1775
Beauchamp AJ, Butler DJ, King D (1999) Weka (Gallirallus australis) recovery plan. Department of Conservation, Wellington 90
Begg AC, Begg NC (1969) James Cook and New Zealand. Government Printer, Wellington
Beggs J (2001) The ecological consequences of social wasps (Vespula sp.) invading an ecosystem that has an abundant carbohydrate resource. Biol Conserv 99:17–28
Bell BD (1978) The Big South Cape islands rat irruption. In: Dingwall PR, Atkinson IAE, Hay C (eds) The ecology and control of rodents in New Zealand nature reserves. Department of Lands and Survey Information Series 4, Wellington, pp 33–40
Blackburn A (1965) Muttonbird islands diary. Notornis 12:191–207
Brook FJ (1999) Changes in the landsnail fauna of Lady Alice Island, northeastern New Zealand. J R Soc N Z 29:135–157
Brown KP (1997) Predation at nests of two New Zealand endemic passerines; implications for bird community restoration. Pac Conserv Biol 3:91–98
Campbell JD, Atkinson IAE (1999) Effects of kiore (Rattus exulans) on recruitment of indigenous coastal trees on northern offshore islands of New Zealand. J R Soc N Z 29:265–290
Campbell JD, Atkinson IAE (2002) Depression of tree recruitment by the Pacific rat (Rattus exulans Peale) on New Zealand’s northern offshore islands. Biol Conserv 107:19–35
Campbell JD, Moller H, Ramsay GW, Watt JC (1984) Observations on foods of kiore (Rattus exulans) found in husking stations on northern offshore islands of New Zealand. N Z J Ecol 7:131–138
Clout MN, Russell JC (2006) The eradication of introduced mammals from New Zealand islands. In: Koike F, Clout MN, Kawamichi M, de Poorter M, Iwatsuki K (eds) Assessment and control of biological invasion risks. Shoukadoh Book Sellers, Kyoto and IUCN, Gland, pp 127–141
Cole NC, Jones CG, Harris S (2005) The need for enemy-free space: the impacts of an invasive gecko on island endemics. Biol Conserv 125:467–474
Courchamp F, Caut S (2005) Use of biological invasions and their control to study the dynamics of interacting populations. In: Cadotte MW, McMahon SM, Fukami T (eds) Conceptual ecology and invasions biology. Springer, New York, pp 253–279
Cree A, Daugherty CH, Hay JM (1995) Reproduction of a rare reptile, the tuatara Sphenodon punctatus, on rat-free and rat-inhabited islands in New Zealand. Conserv Biol 9:373–383
Cromarty PL, Broome KG, Cox A, Empson RA, Hutchinson WM, McFadden I (2002) Eradication planning for invasive alien animal species on islands—the approach developed by the New Zealand Department of Conservation. In: Veitch CR, Clout MN (eds) Turing the tide; the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, IUCN Gland and Cambridge, pp 85–91
Crook IG (1973) The tuatara, Sphenodon punctatus Gray, on islands with and without populations of the Polynesian rat, Rattus exulans (Peale). Proc N Z Ecol Soc 20:115–120
Crowe A (2002) Which New Zealand insect?. Penguin Books, Auckland
Department of Conservation (2002) The penguin guide to New Zealand wildlife. Penguin Books, Auckland
Elliot GP, Merton DV, Jansen PW (2001) Intensive management of a critically endangered species: the kakapo. Biol Conserv 99:121–133
Fukami T, Wardle DA, Bellingham PJ, Mulder CPH, Towns DR, Yeates GW, Bonner KI, Durrett MS, Grant-Hoffman MN, Williamson WM (2006) Above- and below-ground impacts of introduced predators in seabird-dominated island ecosystems. Ecol Lett 9:1299–1307
Gaskin CP, Baird KA (2005) Observations of black and white storm petrels in the Hauraki Gulf, November 2003 to June 2005; were they New Zealand storm petrels? Notornis 52:181–194
Gaze P (2001) Tuatara recovery plan 2001–2011. Threatened species recovery plan 47. Department of Conservation, Wellington
Gibbs GW (2008) The end of an 80-million year experiment: a review of evidence describing the impact of introduced rodents on New Zealand’s ‘mammal-free’ invertebrate fauna. Biol Invas. doi:10.1007/s10530-008-9408-x
Gill BJ, Whitaker AH (1996) New Zealand frogs and reptiles. David Bateman, Auckland
Green CJ (2002) Recovery of invertebrate populations on Tiritiri Matangi Island, New Zealand following eradication of Pacific rats (Rattus exulans). In: Veitch CR, Clout MN (eds) Turing the tide; the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, IUCN Gland and Cambridge, p 407
Guthrie-Smith H (1925) Bird life on island and shore. William Blackwood and Sons, Edinburgh and London
Harper GA (2007) Detecting predation of a burrow-nesting seabird by two introduced predators, using stable isotopes, diet analysis, and experimental removals. Wildl Res 34:443–453
Heather BD, Robertson HA (2000) The field guide to the birds of New Zealand. Penguin, Auckland
Hoare JM, Pledger S, Nelson NJ, Daugherty CH (2007) Avoiding aliens: behavioural plasticity in habitat use enables large, nocturnal geckos to survive Pacific rat invasions. Biol Conserv 136:510–519
Holdaway RN (1999) Introduced predators and avifaunal extinctions in New Zealand. In: MacPhee RDE (ed) Extinctions in near time. Kluwer Academic/Plenum Publishers, New York, pp 189–238
Hunt TL (2007) Rethinking Easter Island’s ecological catastrophe. J Archaeol Sci 34:485–502
Imber MJ, West JA, Cooper WJ (2003) Breeding biology of Cook’s petrel (Pterodroma cookii) on Hauturu (Little Barrier Island) and Whenua Hou (Codfish Island). Notornis 50:221–230
Innes J (2005) Ship rat. In: King CM (ed) The handbook of New Zealand mammals, 2nd edn. Melbourne, Oxford University Press, pp 187–203
Kuschel G, Worthy TH (1996) Past distribution of large weevils (Coleoptera:Curculionidae) in the South Island, New Zealand, based on Holocene fossil remains. N Z Entomol 19:15–22
Lloyd B (2005a) Lesser short-tailed bat. In: King CM (ed) The handbook of New Zealand mammals, 2nd edn. Melbourne, Oxford University Press, pp 110–126
Lloyd B (2005b) Greater short-tailed bat. In: King CM (ed) The handbook of New Zealand mammals, 2nd edn. Melbourne, Oxford University Press, pp 127–129
Markwell T, Daugherty CH (2002) Invertebrate and lizard abundance is greater of seabird-inhabited islands than on seabird-free islands in the Marlborough Sounds, New Zealand. Ecoscience 9:293–299
McCallum J (1986) Evidence of predation by kiore upon lizards from the Mokohinau Islands. N Z J Ecol 9:83–87
McCallum J, Bellingham PJ, Hay JR, Hitchmough RA (1984) The birds of the Chickens Islands, northern New Zealand. Tane 30:105–124
McGuinness CA (2001) The conservation requirements of New Zealand’s nationally threatened invertebrates. Threatened Species Occasional Publication 20. Department of Conservation, Wellington
McIntyre M (2001) The ecology of some large weta species in New Zealand. In: Field LH (ed) The biology of wetas, king crickets and their allies. Wallingford, CAB International, pp 225–242
Meads M (1990) Forgotten fauna. DSIR Publishing, Wellington
Mulder CPH, Keall SN (2001) Burrowing seabirds and reptiles: impacts on seeds, seedlings and soils in an island forest in New Zealand. Oecologia 127:350–360
Mulder CPH, Grant-Hoffman MN, Towns DR, Bellingham PJ, Wardle DA, Durrett MS, Fukami T, Bonner KI (2008) Direct and indirect effects of rats: does rat eradication restore ecosystem functioning of New Zealand seabird islands? Biol Invas. doi:10.1007/s10530-008-9396-x
Myers JH, Simberloff D, Kuris AM, Carey JR (2000) Eradication revisited: dealing with exotic species. Trends Ecol Evol 15:316–320
Newman DG (1987) Burrow use and population densities of tuatara (Sphenodon punctatus) and how they are influenced by fairy prions (Pachyptila turtur) on Stephens Island, New Zealand. Herpetologica 43:336–344
Parrish GR, Sherley GH (1993) Invertebrates of Moutopao Island, Northland, New Zealand. Tane 34:45–52
Pierce RJ (2002) Kiore (Rattus exulans) impact on breeding success of Pycroft’s petrels and little shearwaters. DOC Science Internal Series 39. Department of Conservation, Wellington
Poole AL, Adams NM (1990) Trees and shrubs of New Zealand. DSIR Publishing, Wellington
Reserves Act (1977) New Zealand Government, Wellington
Rayner M, Hauber ME, Imber MJ, Stamp RK, Clout MN (2007) Spatial heterogeneity of mesopredator release within an oceanic island system. Proc Natl Acad Sci USA 104:20862–20865
Reed AH, Reed AW (1969) Captain Cook in New Zealand. Reed, Wellington
Robb J (1986) New Zealand amphibians and reptiles in colour. Collins, Auckland Revised edition
Tennyson A, Martinson P (2006) Extinct birds of New Zealand. Te Papa Press, Wellington
Towns DR (1991) Response of lizard assemblages in the Mercury Islands, New Zealand, to removal of an introduced rodent: the kiore (Rattus exulans). J R Soc N Z 21:119–136
Towns DR (1996) Changes in habitat use by lizards on a New Zealand island following removal of the introduced Pacific rat Rattus exulans. Pac Conserv Biol 2:286–292
Towns DR (1999) Cyclodina spp. skink recovery plan. Threatened Species Recovery Plan No. 27. Department of Conservation, Wellington
Towns DR (2002a) Interactions between geckos, honeydew scale insects and host plants revealed on islands in northern New Zealand, following eradication of introduced rats and rabbits. In: Veitch CR, Clout MN (eds) Turing the tide; the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, IUCN Gland and Cambridge, pp 329–335
Towns DR (2002b) Korapuki Island as a case study for restoration of insular ecosystems in New Zealand. J Biogeogr 29:593–607
Towns DR, Atkinson IAE (2004) Restoration plan for Korapuki Island (Mercury Islands) 2004–2024. Department of Conservation, Wellington
Towns DR, Broome KG (2003) From small Maria to massive Campbell: 40 years of rat eradications from New Zealand islands. N Z J of Zool 30:377–398
Towns DR, Daugherty CH (1994) Patterns of range contractions and extinctions in the New Zealand herpetofauna following human colonisation. N Z J of Zool 21:325–339
Towns DR, Simberloff D, Atkinson IAE (1997) Restoration of New Zealand islands: redressing the effects of introduced species. Pac Conserv Biol 3:99–124
Towns DR, Parrish GR, Westbrooke I (2003) Inferring vulnerability to introduced predators without experimental demonstration: case study of Suter’s skink in New Zealand. Conserv Biol 17:1361–1371
Towns DR, Atkinson IAE, Daugherty CH (2006) Have the harmful effects of introduced rats on islands been exaggerated? Biol Invasions 8:863–891
Towns DR, Parrish GR, Tyrrell CL, Ussher GT, Cree A, Newman DG, Whitaker AH, Westbrooke I (2007) Responses of tuatara (Sphenodon punctatus) to removal of Pacific rats from islands. Conserv Biol 21:1021–1031
Ussher GT (1999) Tuatara (Sphenodon punctatus) feeding ecology in the presence of kiore (Rattus exulans). N Z J of Zool 26:117–125
Veltman C (1996) Investigating causes of population decline in New Zealand plants and animals: introduction to a symposium. New Zealand Journal of Ecology 20:1–5
Watt JC (1986) Beetles (Coleoptera) of the offshore islands of northern New Zealand. In: Wright AE, Beever RE (eds) The offshore islands of northern New Zealand. Department of Lands and Survey Information Series 16, Wellington, pp 221–228
Whitaker AH (1978) The effects of rodents on reptiles and amphibians. In: Dingwall PR, Atkinson IAE, Hay C (eds) The ecology and control of rodents in New Zealand nature reserves. . Department of Lands and Survey Information Series 4, Wellington, pp 75–86
Wilmshurst JM, Anderson AJ, Higham TFG, Worthy TH (2008) Dating the late prehistoric dispersal of Polynesians to New Zealand using the commensal Pacific rat. Proc Natl Acad Sci USA 105:7676–7680
Worthy TH (1987) Osteology of Leiopelma (Amphibia: Leiopelmatidae) and descriptions of three new subfossil Leiopelma species. J R Soc N Z 17:201–251
Worthy TH (1991) Fossil skink bones from Northland, New Zealand, and description of a new species of Cyclodina, Scincidae. J R Soc N Z 21:329–348
Worthy TH, Holdaway RN (2002) The lost world of the moa. Canterbury University Press, Christchurch
Acknowledgments
I thank Ian Atkinson, Grant Harper and James Russell for their useful comments on the manuscript, the organisers of the conference on Rats, Humans and their Impacts on Islands for their invitation to present this paper and two anonymous reviewers for their thoughtful comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Towns, D.R. Eradications as reverse invasions: lessons from Pacific rat (Rattus exulans) removals on New Zealand islands. Biol Invasions 11, 1719–1733 (2009). https://doi.org/10.1007/s10530-008-9399-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-008-9399-7