
Abstract
The number and scale of island invasive species eradications is growing, but quantitative evidence of the conservation efficacy of passive recovery is limited. We compare relative abundances of breeding birds on Hawadax Island (formerly named Rat island), Aleutian Archipelago, Alaska, pre- and post- rat eradication to examine short-term (<1 year post-eradication) changes due to rodenticide application, and medium-term (5 years post-eradication) changes due to the absence of invasive rats. In the short term, Bald Eagle (Haliaeetus leucocephalus) numbers decreased from 24 individuals pre-eradication to two individuals <1 year post-eradication, but recovered to 10 individuals (42 % of pre-eradication) 5 years post-eradication, with all individuals nesting (63 % of the pre-eradication nesting). Five years post-eradication relative abundances of most terrestrial birds surveyed using point counts either significantly increased [Gray-crowned Rosy Finch (Leucosticte tephrocotis), Lapland Longspur (Calcarius lapponicus), Snow Bunting (Plectrophenax nivalis), Song Sparrow (Melospiza melodia)] or did not differ [Pacific Wren (Troglodytes troglodytes)]. Shorebirds also increased 5 years post-eradication with Black Oystercatchers (Haematopus palliates) increasing fivefold, and Rock Sandpiper (Calidris ptilocnemis) nesting increasing from one to five nests. We confirmed two species of ground nesting seabirds [Tufted Puffin (Fratercula cirrhata) and Leach’s Storm-petrel (Oceanodroma leucohoa)] as nesting (puffin) or engaged in courtship behavior (Storm-petrel) 5 years post-eradication. Our results indicate that despite the short-term impact on Bald Eagles, and without further human intervention, most terrestrial and marine birds have newly-colonized, re-colonized, or increased in abundance following the eradication of invasive rats.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Islands make up roughly 5 % of the Earth’s land area, yet host a disproportionate amount of global biodiversity including 19 % of all bird species (Tershy et al. 2015). Unfortunately, island breeding birds are disproportionately threatened, accounting for 95.3 % of recent historical (since 1500) avian extinctions (Loehle and Eschenbach 2012). A primary cause of insular avian extinctions is introduced alien species (Clavero et al. 2009), with invasive rodents in the genera Rattus among the most damaging invasive animals to island-breeding birds (Towns et al. 2006; Howald et al. 2007). Invasive rodents have invaded ~90 % of the world’s archipelagos (Jones 2010), but can be eradicated from many islands, with over 700 successful operations globally (Keitt et al. 2011; DIISE 2015).
While there is compelling evidence that invasive alien species have a broad range of direct and indirect impacts on island ecosystems (e.g. Croll et al. 2005; Jones et al. 2008; Kurle et al. 2008; Towns et al. 2006), and the number and scale of eradication efforts is growing (Keitt et al. 2011), quantitative evidence of the efficacy of eradication as a tool for avian conservation on islands is surprisingly limited (Jones 2010). Regional reviews document benefits for native species on French (Lorvelec and Pascal 2005) and New Zealand (Bellingham et al. 2010) islands, and a review by Lavers et al. (2010) demonstrated that bird demographic parameters respond positively to rodent eradications. In addition, Towns et al. (2006) documented plant and animal recovery from eradications, but commented that the evidence was complicated by lack of pre-eradication data, confounding effects of not eradicating all invasive alien species on an island, and limited post-eradication monitoring. Moreover, Kappes and Jones (2014), recently suggested the need to evaluate the necessity of active versus passive restoration programs following invasive mammal eradication. In this paper we document passive recovery of bird communities following a rodent eradication in autumn 2009 on Hawadax Island (formerly Rat Island), Aleutian Archipelago, Alaska.
The Aleutian Archipelago has 299 bird species, 80 of which (27 %) are residents or migratory breeders, including 26 seabird species (Gibson and Byrd 2007). The archipelago had no terrestrial mammals west of Umnak Island until Arctic foxes (Alopex lagopus) were intentionally introduced by fur ranchers and Norway rats (Rattus norvegicus) were accidentally introduced on several islands, including Hawadax Island, in the 1700s (Bailey 1993). In the Aleutians, fox and rat predation reduces abundance of nesting birds (particularly seabirds) (Bailey 1993; Ebbert and Byrd 2002; Byrd et al. 2005; Major et al. 2006), potentially decreases abundance of non-breeding birds, and can indirectly alter community processes and structure (e.g. intertidal and plant communities) (Croll et al. 2005; Kurle et al. 2008). On Hawadax Island, the combined impacts of introduced foxes and rats were likely significant, due to the synergistic effects from a carnivore and an omnivore as well as the potential for apparent competition between rats and native birds mediated by fox hyperpredation (the presence of an alternate prey leads to an impact of the predator on the prey via an elevated density of the predator) on native birds (reviewed in Russell 2011; Towns et al. 2011). Foxes were successfully eradicated in 1984 (Bailey 1993), but invasive rats persisted and apparently limited avian recovery compared to surrounding islands from which foxes were eradicated (J. Williams, pers. obs.). Rats were eradicated using an aerial broadcast of cereal grain pellets containing rodenticide (Brodifacoum) in September–October 2008 with an overall goal of “improving habitat quality for native species” (US Fish and Wildlife Service 2007). Details on the eradication and eradication confirmation are presented in Buckelew et al. (2011). Here we compare relative abundances of Hawadax breeding birds pre- and post-rat eradication to examine (1) short-term changes that may be due to the application of rodenticide (<1 year post-eradication), and (2) medium-term changes that may be due to the absence of invasive rats (5 years post-eradication).
Methods
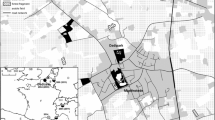
Hawadax Island (51.80°N, 178.30°E) (known as Rat Island until renamed in 2012; U.S. Board on Geographic Names 2012) is located in the Rat Islands group in the western Aleutian Islands and part of the Alaska Maritime National Wildlife Refuge (Fig. 1). The 2780 ha island has steep coastal cliffs along the majority of the coastline backed by rolling hills and plateaus, with nine small clusters of lakes and no human inhabitants. The interior contains a small range of mountains with a peak elevation of 400 m. There are more than 30 offshore rock stacks and islets, the largest of which is ~4 ha and <1 km offshore. Rat eradication was conducted after the end of seasonal bird breeding activities, when food resources for rats were limited (Buckelew et al. 2011).

The location of Hawadax Island in the Rat Islands Group, central Aleutian Island Archipelago, Alaska, USA
To evaluate the impacts of the rat eradication on island bird populations we conducted a variety of surveys in May–June to coincide with seasonal breeding activities of native birds. The range of survey types utilized were selected to provide a robust index of changes in abundance of all major groups of birds that commonly occur on the island, and to detect changes in presence of rare species. For some species we censused entire island populations (e.g. Bald Eagles (Haliaeetus leucocephalus), Peregrine Falcons (Falco peregrinus), Glaucous-winged Gull (Larus glaucescens) nests). However, due to difficulties inherent in measuring absolute abundance in highly mobile species with incomplete detectability (Elzinga et al. 2001), we measured relative rather than absolute abundance for most taxa. Here we present results of data gathered in 2008 (pre-eradication), 2009 (<1 year post-eradication), and 2013 (5 years post-eradication). Data from 2009 were used to examine short-term (<1 year post-eradication), potentially negative impacts of the eradication operations. Data from 2013 were used to examine longer-term (5 years post-eradication) recovery from invasive rat impacts.
We conducted all visual surveys (point count, beach, seabird, coastline surveys) during favorable weather: wind less than 15 knots, visibility greater than 250 m, and little or no precipitation. Analyses of point and beach surveys conducted on pilot data collected in 2007 indicated that there was no effect of time of day (morning vs. afternoon) on number of birds detected during surveys conducted under comparable weather conditions (Buckelew et al. 2008). To avoid confounding results from excessive zero count data, we performed interannual comparisons of survey data using nonparametric Van der Waerden tests, followed by post hoc comparisons using Wilcoxon tests. Alpha level for all tests was 0.05.
Point count surveys
For passerine bird population indices, we selected random points (44 points in 2008, 57 points in 2009, and 75 points in 2013) from a sampling grid of points at least 200 m apart over the entire island. At each point, observers conducted bird point counts as a relative measure of landbird abundance on the island. During a 5-min sampling period between 0600 and 1000 HADT, one observer recorded all species seen or heard in two distance categories: within 50 m of the point and beyond 50 m. We calculated relative abundance as the mean number of birds (±SE) observed per point by species.
For our point count analyses, we estimate relative, rather than absolute densities of terrestrial birds. While there is the potential for bias by not accounting for variability in detection across species, we believe this would have limited effect on our results for several reasons: (1) we restricted the radius of all surveys to 50 m to assure all individuals within the observation field were recorded, (2) we limited observations to favorable viewing conditions, (3) we conducted surveys during the same months (May, June) each year, (4) we limit our comparisons to changes within species across years, all observers (total = 20 across all years) were experienced, (5) the vegetation of Hawadax Island is highly homogeneous consisting almost exclusively of Leymus grass and Empetrum shrub which was below waist height with no trees, and (6) the terrestrial bird diversity is low, and we restricted our comparisons to a limited, more common, subset of species. Regardless, there is a potential for error resulting from not accounting for detection probability. In addition, we assume that the detectability of individual species did not change pre- versus post-eradication (e.g. behavioral changes). Because most species included in our study regularly migrate between islands with different conditions, we feel that significant behavior-related changes in detectability are unlikely.
Beach surveys
We conducted beach surveys along the entire length of all accessible (n = 16) beaches to assess relative abundance of bird species that concentrate along the coastal periphery [Pacific Wren (Troglodytes troglodytes), Song Sparrow (Melospiza melodia), Black Oystercatcher (Haematopus palliates), Glaucous-winged Gull (Larus glaucescens), Rock Sandpiper (Calidris ptilocnemis)] and are under-sampled using randomized standard point counts across the island. For each survey, an observer walked along the storm line for each beach transect, counting all birds seen or heard from the storm line to the water’s edge and the storm line to 50 m inland. Both sound and sight were used for detections. Start and end location of each transect was recorded using GPS, and we measured the length of each transect using ArcGIS 10.2 (ESRI 2013). Total count of birds detected on a beach transect was divided by length of beach to yield a relative measure of bird abundance (number km−1). To minimize effects of variation in time and conditions of sampling, we conducted five replicate surveys of 16 fixed beach transects on the larger beaches located around the island. For each year, the five replicate surveys for a beach were averaged and used as a sample for that beach for that year (i.e. averaged counts for each beach in a year was considered a sample).
Surface-nesting seabird surveys
We recorded locations and counted all active nest sites in all ground nesting seabird colonies (e.g. Glaucous-winged Gull) encountered during other survey activities. We considered nests active if they had fresh nest material.
Coastline surveys
An island-wide census of birds of prey was generated by a coastline visual survey each year where the entire coastal periphery of Hawadax Island was traversed. An observer followed the coastal contour and scanned for raptor nests or displays of territorial or nesting behavior. Shorelines that could not be accessed on foot were scanned using binoculars. We recorded the number of eggs or chicks in each nest when visible. Completely inaccessible sections of the coastline (e.g. cliff faces and sea stacks) were examined from a small skiff, and these nests were added to the coastline survey. In addition to nest contents, we recorded number of territorial or nesting adults.
Acoustic surveys
To detect recruitment of new species and measure changes in distribution and acoustic activity of birds, we deployed passive acoustic sensors. Passive acoustic surveys provide a measure of target species occupancy (presence/absence) at independent survey points, and increases the probability of detecting nocturnal cryptic-nesting species (i.e., Leach’s Storm-petrel) and recruitment events which are difficult to detect with visual surveys or are rare in time and space (Borker et al. 2014). We established 23 acoustic monitoring stations: 17 focused on detecting terrestrial birds, and six focused on seabirds.
For terrestrial birds (Lapland Longspur, Pacific Wren, Rock Ptarmigan (Lagopus muta), Snow Bunting, and Song Sparrow), acoustic sensors were randomly deployed using the same technique used for point counts at each of 17 stations during the month of June at the same locations each survey year (2008, 2009, 2013). Each sensor recorded a 4-h period just after local sunrise (0700–1100 HADT), and each site was sampled for three or four consecutive mornings.
In 2009 (<1 year post-eradication) and 2013 (5 years post-eradication), we deployed six additional sensors at potential seabird breeding habitat (based on slope, coastal proximity, and soil and vegetation characteristics) to monitor recruitment of breeding seabird at the same locations across years. The sensors recorded 1 min of every 10 min for the duration of deployment. In 2009, each sensor was deployed for 5–10 consecutive nights. In 2013, each sensor was deployed for over 70 consecutive nights at each location. Seabird acoustic surveys were only carried out post-eradication (2009, 2013); thus pre- and post-eradication comparisons of acoustic activity were not possible.
We used Autonomous Recording Units (ARU’s) (Bioacoustics Research Program, Cornell Lab of Ornithology) for acoustic surveys conducted in 2008 and 2009. In 2013, we deployed Song Meter 2 acoustic sensors (Wildlife Acoustics, Inc.). Borker et al. (2014) found no significant difference in call rates measured using the two sensor types. Calls were detected using XBAT (Bioacoustics Research Program, Lab of Ornithology, Cornell University). Briefly, we classified calls using spectrogram cross-correlation between spectral qualities of a template sound (the calls of target species) and sounds on audio files recorded in the field. We employed the same species templates and correlation thresholds to detect and quantify call rates for all field seasons to ensure consistency between years. We quantified call detections for Lapland Longspur, Pacific Wren, Snow Bunting, and Song Sparrow. Recordings were not examined for Gray-crowned Rosy Finches (Leucosticte tephrocotis) because suitable template calls were not available. For seabirds, we analyzed recordings for Leach’s Storm-petrel (Oceanodroma leucorhoa), a species known to be nesting on nearby Ayugadak Island. For each location we classified a bird species as present if a vocalization of that species was detected. Due to differences in recording protocols in 2009 vs. 2013, we restricted analyses to the 4 nights with the same lunar conditions in each year. We calculated Leach’s Storm-petrel mean calls min−1 ± SE and used a paired t test to compare 2009 vs. 2013 call rates (alpha level 0.05).
Incidental visual observations
We recorded opportunistic observations of bird species of special interest (e.g. raptors, burrow-nesting seabirds), or those not recorded in standard surveys. For each opportunistic observation, number of individuals, age class (when possible), date and time, and GPS position of any nests were recorded.
Results
Terrestrial bird point counts
Relative abundances of all five terrestrial birds surveyed using point counts were either not different (Pacific Wren) or significantly increased (Gray-crowned Rosy Finch, Lapland Longspur, Snow Bunting, Song Sparrow) post- versus pre-eradication (Fig. 2). Snow Buntings and Song Sparrows were not recorded in point counts either pre- or <1 year post-eradication, but were commonly observed 5 years post-eradication.

Annual changes in passerine abundance (mean ± SE) on Hawadax Island pre- versus post-eradication (2008 vs. 2009 and 2013) from point count surveys; median values are shown in parentheses. Comparison conducted using Van der Waerden test (Lapland Longspur, Pacific Wren) or one-sample t test (0 hypothesized pre-eradication mean; Gray-crowned Rosy Finch, Snow Bunting, Song Sparrow). A–C indicate significantly different data, p < 0.05
Beach surveys
Terrestrial bird beach survey numbers for Pacific Wrens were not different pre- vs. <1 year post-eradication, but were significantly reduced (compared to pre- and <1 year post) 5 years post-eradication (Fig. 3). Song Sparrows were not observed in either pre- or <1 year post-eradication surveys, but were found in surveys 5 years post-eradication (Fig. 3). For seabird and shorebird counts, Glaucous-winged Gulls were significantly more abundant both <1 and 5 years post-eradication, Black Oystercatcher abundance increased significantly 5 years post-eradication, and Rock Sandpiper abundance was unchanged pre- versus post-eradication (Fig. 4).

Annual changes in passerine abundance (mean ± SE) on Hawadax Island pre- versus post-eradication (2008 vs. 2009 and 2013) from beach surveys; median values are shown in parentheses. Pacific Wren comparisons conducted using Van der Waerden test. Song Sparrow compared using one-sample t test (0 hypothesized pre-eradication mean). A, B indicate significantly different data, p < 0.05

Annual changes in shorebird and seabird abundance (mean ± SE) on Hawadax Island pre- versus post-eradication (2008 vs. 2009 and 2013) from beach surveys; median values are shown in parentheses. Comparisons conducted using Van der Waerden test. A, B indicate significantly different data, p < 0.05
Surface-nesting seabird survey
Two Glaucous-winged Gull colonies with active nests were identified pre- and post-eradication. Total number of active nests in both colonies was greater in both post-eradication surveys compared to pre-eradication (Table 1).
Coastline survey
Nesting Bald Eagle numbers declined drastically <1 year post-eradication, but 5 years post-eradication recovered to 63 % of pre-eradication numbers respectively (Table 2). While 12 Bald Eagle chicks were observed in nests in pre-eradication surveys (2008), chicks were not observed post-eradication until 2013, when three were observed. The number of territorial pairs of Peregrine Falcons was uniformly low and variable across the three periods (Table 2).
Acoustic surveys
The proportion of acoustic survey locations detecting Pacific Wrens and Snow Buntings increased post-eradication (Table 3). Similar to point and beach surveys, Song Sparrow songs were not recorded at any location either pre- nor <1 year post-eradication, but were recorded at 53 % of locations 5 years post-eradication.
Leach’s Storm-petrel mean call rates did not differ significantly when comparing four equivalent survey nights in 2009 and 2013 (2009 = 0.003 calls per minute ± 0.003 SE, 2013 = 0.170 ± 0.098 SE, t = 1.03, df = 5, p = 0.18). However, proportion of sites with Leach’s calling activity increased from 17 % (1) to 50 % (3) of seabird survey sites from <1 year post-eradication to 5 years post-eradication (Table 3); although this difference was not significant (and seabirds were not acoustically surveyed pre-eradication). Finally, although comparable acoustic data are not available for 2009, acoustic survey data from 2013 (with >70 survey nights per survey point) documented Leach’s Storm-petrel calling activity at all 6 seabird survey points, including calling on 88 % of survey nights and consistent courtship vocalizations at one survey point on the southeastern tip of the island. Together these data suggest increasing Leach’s Storm-petrel presence on Hawadax Island.
Incidental observations
During incidental observations and systematic surveys we recorded 20 bird species pre-eradication (2008) versus 27 species post-eradication (2013), including 63 nests from 10 species (Table 4). Pre-eradication (2008), we encountered one Black Oystercatcher and one Rock Sandpiper nest; post-eradication (2013) we encountered six Black Oystercatcher and five Rock Sandpiper nests. Tufted Puffins (Fratercula cirrhata) were reported as likely not breeding on the island prior to the eradiation (US Fish and Wildlife Service 2007), and we found no burrows during survey activities pre-eradication, but found six active burrows in 2013 in an area that had been thoroughly searched in previous years. An egg was visible in one of the burrows, and adults were found in the other five. Finally, we incidentally observed one Common Eider (Somateria mollissima) and two Song Sparrow nests with chicks 5 years post-eradication, confirming their nesting presence on the island. Three non-Aleutian breeding species incidentally observed pre-eradiation were not observed post-eradication [Semipalmated Plover (Charadrius semipalmatus), Brambling (Fingilla motifringilla), Sandhill Crane (Grus canadensis); Table 4].
Discussion
Rats have both direct and indirect impacts on a variety of native taxa. For birds, direct effects include predation of eggs, chicks, and adults (Jones et al. 2008). Within the Aleutian Islands, rat predation on intertidal-feeding marine birds also has significant indirect effects on intertidal invertebrate communities (Kurle et al. 2008). Reduced seabird abundance has also been shown to significantly change plant community structure throughout the Aleutian Archipelago (Croll et al. 2005).
Introduced vertebrates, especially rats and foxes, have severely reduced Aleutian avian populations, particularly seabirds (Ebbert and Byrd 2002; Byrd et al. 2005; Major et al. 2006; Buxton et al. 2013). As a result they have been the focus of an intensive removal program: foxes have been eradicated from over 40 islands since 1949 (Ebbert and Byrd 2002). Artic foxes were removed from Hawadax Island in 1984 (USFWS 2007), and the eradication of rats from Hawadax Island was one of several potential rodent eradications in the region. While fourfold to fivefold increases in seabirds and 35-fold increases in Aleutian cackling geese have been reported after fox removal (Ebbert and Byrd 2002), detailed quantitative evidence of post-eradication recovery for other Aleutian species is limited (with the exception of recent models based on acoustic data showing recovery of burrow nesting Aleutian seabirds; Buxton et al. 2013). Moreover, Kappes and Jones (2014) suggested the long-term ability of seabirds to passively recover on islands following eradications may be limited due low reproductive rates/fecundity, intermittent breeding, and high natal philopatry.
Short-term impacts
Some negative short-term (<1 year post eradication) impacts occurred. Brodifacoum (an anti-coagulant) is toxic to many vertebrates at varying doses, but is generally highly toxic to mammals and birds when an exposure pathway exists (Howald et al. 2000). Because there are no native terrestrial amphibians, reptiles, or mammals on Hawadax Island, mortality of native birds was the primary non-target concern (US Fish and Wildlife Service 2007). Brodifacoum has potential impacts on herbivorous and omnivorous birds via direct ingestion of cereal bait and to predatory and scavenging birds via secondary, indirect, ingestion of brodifacoum-contaminated carcasses (Donlan et al. 2003; Howald et al. 2009). Buckelew et al. (2011) reported non-target mortality of Glaucous-winged Gulls and Bald Eagles resulting from scavenging of rat and secondarily-poisoned gull carcasses on Hawadax Island. They recovered 320 gull and 46 eagle carcasses during beach carcass surveys and opportunistic recoveries in 2009, nine to 11 months after the eradication activities ended. In post-mortem tissue analysis 24 of 34 (66 %) of gull carcasses tested positively for brodifacoum while 12 of 16 (75 %) of eagle carcasses had potentially lethal brodifacoum levels (Paul and Salmon 2010).
Coastline surveys for nesting eagles conducted <1 year post-eradication reflect this mortality, with number of eagles decreasing from a pre-eradication maximum of 24 to a post-eradication minimum of two. However, 5 years post-eradication the number had recovered to 42 % of pre-eradication counts. All individuals were nesting, thus nesting effort recovered to 63 % of pre-eradication level. Peregrine Falcon numbers appeared unaffected both short- and long-term by the eradication, but their low and variable numbers make it difficult to discern clear patterns.
Although there was some non-target Glaucous-winged Gull mortality from the eradication operation, the breeding population may have benefited more from release of rat impacts. Only five gull nests were observed pre-eradication, but increased to 13 nests (160 %) <1 year post-eradication and 27 nests (440 %) 5 years post-eradication. Glaucous-winged Gulls counted in beach transects likewise increased 250 % <1 year post-eradication and remained at levels 150 % greater 5 years post-eradication. The delayed age of first reproduction aspect of gull life history suggests that these increases in nest counts and abundance resulted from recruitment of gulls from nearby islands. With time, increased nesting on Hawadax should lead to further increases in gull numbers. We did not detect declines in any other breeding bird species monitored on Hawadax indicating limited short-term non-target negative impacts of the eradication on these species.
Medium-term recovery
Most monitored bird populations showed positive changes 5 years post-eradication of rats from Hawadax Island. Non-vagrant species richness increased by seven species pre- vs. 5 years post-eradication, and several terrestrial, shorebird, and seabird species increased in relative abundance. In addition, several species were confirmed breeding on the island for the first time 5 years post-eradication, presumably after having been extirpated.
For terrestrial birds, point and acoustic surveys showed no significant change in Pacific Wren abundance (although beach surveys showed a decline in wren abundance 5 years post-eradication). Two terrestrial species (Snow Buntings and Song Sparrows) historically present on Hawadax island but not encountered in pre-eradication or <1 year post-eradication surveys were commonly encountered 5 years post-eradication, with visual confirmation of renewed Song Sparrow nesting on the island in incidental observations.
For shorebirds, the relative abundance of Black Oystercatchers recorded in beach transects increased fivefold pre- versus 5 years post-eradication. While only one Black Oystercatcher nest and no Rock Sandpiper nests were encountered pre-eradication, by 5 years post-eradication, six and five Black Oystercatcher and Rock Sandpiper nests were encountered, respectively.
The majority of terrestrial, waterfowl, and shorebird species regularly observed in surveys thus appear to have benefitted from rat eradication. Most of these species have life histories (younger age of first reproduction, greater fecundity) indicating that these increases are partially due to increases in Hawadax Island populations released from rodent impacts.
Seabirds generally exhibit life history characteristics (delayed reproduction, low fecundity) that lead to delayed responses to release from rodent impacts (Jones et al. 2008; Howald et al. 2009; Jones 2010; Lavers et al. 2010; Towns et al. 2011). Nonetheless, in addition to increased Glaucous-winged Gull nesting effort and relative abundance, two species of burrowing seabird (Tufted Puffin and Leach’s Storm-petrel) likely not breeding on the island prior to rat eradication (US Fish and Wildlife Service 2007) were first confirmed as nesting (puffin) or engaged in courtship behavior (storm-petrel) post-eradication likely due to the close proximity of source populations.
Evidence from animal bones recovered from archeological studies on Hawadax Island indicate that a range of burrow-nesting seabirds bred on the island including Ancient Murrelets (Synthliboramphus antiquus), Parakeet Auklets (Aethia psittacula), Crested Auklets (Aethia cristatella), and Whiskered Auklets (Aethia pygmaea; Funk 2012). Their recolonization is likely constrained by low fecundity, proximity of source populations and breeding behavior (e.g. coloniality, Allee effects). As a result, more complete recovery will likely require more time. Buxton et al. (2013) suggested that recolonization by extirpated species depends on the distance to a source population (within 25 km) and local diversity of other seabirds. Kappes and Jones (2014) suggested that, for islands >25 km from source populations, passive recovery of seabirds may not occur and suggested managers consider active seabird restoration programs for remote islands. However, Hawadax Island is less than 2 km from potential source populations of seabirds, including Tufted Puffins and Leach’s Storm-petrels (Williams, pers. obs.). In addition, populations of these other seabird species occur on other islands in the Rat Island group, less than 25 km from Hawadax. The relatively rapid appearance of two of six seabird species extirpated from the island support both Buxton and Kappes and Jones’ hypotheses, and it is likely that active recovery is not necessary for seabirds on Hawadax Island.
Bald Eagle nests have only recovered to 63 % of pre-eradication numbers. However, they have relatively slow life histories, and it will take time for their numbers to return to pre-eradication levels. In the long term, seabirds are the primary prey of Aleutian Bald Eagles and Peregrine Falcons and their densities are generally greater on islands with larger concentrations of seabirds (Anthony et al. 2008). As seabird populations increase on Hawadax Island, we predict Bald Eagle and Peregrine Falcon numbers will eventually exceed pre-eradication numbers.
Acknowledging that there is likely interannual variability in avian population abundances on Hawadax island, our results indicate that despite the negative initial impact on Bald Eagles, the goal of the Hawadax Island eradication: “improving habitat quality for native species” (US Fish and Wildlife Service 2007) has been achieved for avian species. A range of terrestrial and marine birds have newly-colonized, re-colonized, or increased in abundance following the eradication of invasive rats.
Moving forward, we anticipate the effects of rat eradication will extend beyond benefits to target species and lead to restoration of important ecosystem functions as avian populations on Hawadax continue to recover. There are no native terrestrial mammals in the Aleutians, and insect communities are impoverished at higher latitudes. As a result, functions that birds provide (e.g., seed dispersal, pollination, scavenging, nutrient subsidy, and ecosystem engineering) may be particularly important on Hawadax Island. In particular, we anticipate the recovery of important bird-mediated ecosystem processes, such as nutrient subsidies and trophic interactions, described for the Aleutian Archipelago (Croll et al. 2005; Maron et al. 2006; Kurle et al. 2008). Due to time lags inherent in recovery from indirect effects, it will likely take longer (decades) for these functional changes to be fully manifested on Hawadax Island (Jones et al. 2008; Towns 2008; Russell 2011; Towns et al. 2011).
References
Anthony RG, Estes JA, Ricca MA, Miles AK, Forsman ED (2008) Bald eagles and sea otters in the Aleutian Archipelago: indirect effects of trophic cascades. Ecology 89:2725–2735
Bailey E (1993) Introduction of Foxes to Alaskan Islands—history, effects on avifauna, and eradication. US Fish and Wildlife Service Resource Publication, Washington
Bellingham PJ, Towns DR, Cameron EK, Davis JJ, Wardle DA, Wilmshurst JM, Mulder CPH (2010) Feathers to fur New Zealand island restoration: seabirds, predators, and the importance of history. N Z J Ecol 34:115–136
Borker AL, McKown MW, Ackerman JT, Eagles-Smith CA, Tershy BR, Croll DA (2014) Vocal activity as a low cost and scalable index of seabird colony size. Conserv Biol 28:1100–1108
Buckelew S, Howald G, MacLean S, Byrd V, Daniel L, Ebbert S, Meeks W (2008) Rat Island habitat restoration project: operational report. Report to USFWS. Island Conservation, Santa Cruz
Buckelew S, Byrd V, Howald G, MacLean S, Sheppard J (2011) Preliminary ecosystem response following invasive Norway rat eradication on Rat Island, Aleutian Islands, Alaska. In: Veitch CR, Clout MN, Towns DR (eds) Island invasives: eradication and management. International Union for the Conservation of Nature, Gland, pp 275–279
Buxton RT, Major HL, Jones IL, Williams JC (2013) Examining patterns in nocturnal seabird activity and recovery across the Western Aleutian Islands, Alaska, using automated acoustic recording. Auk 130:331–341
Byrd GV, Renner HM, Renner M (2005) Distribution patterns and population trends of breeding seabirds in the Aleutian Islands. Fish Oceanogr 14:139–159
Clavero M, Brotons L, Pons P, Sol D (2009) Prominent role of invasive species in avian biodiversity loss. Biol Conserv 142:2043–2049
Croll DA, Maron JL, Estes JA, Danner EM, Byrd GV (2005) Introduced predators transform subarctic islands from grassland to tundra. Science 307:1959–1961
DIISE (2015) The database of island invasive species eradications. Developed by Island Conservation, Coastal Conservation Action Laboratory UCSC, IUCN SSC Invasive Species Specialist Group, University of Auckland and Landcare Research New Zealand. http://diise.islandconservation.org. Accessed 20 Oct 2015
Donlan CJ, Howald GR, Tershy BR, Croll DA (2003) Evaluating alternative rodenticides for island conservation: roof rat eradication from the San Jorge Islands, Mexico. Biol Conserv 114:29–34
Ebbert SE, Byrd GV (2002) Eradications of invasive species to restore natural biological diversity on Alaska Maritime National Wildlife Refuge. In: Veitch CR, Clout MN, Towns DR (eds) Island Invasives: eradication and management. International Union for the Conservation of Nature, Gland, Switzerland, pp 102–109
Elzinga CL, Salzer DW, Willoughby JW, Gibbs JP (2001) Measuring and monitoring plant and animal populations. Blackwell Science Inc, Malden
ESRI (Environmental Systems Research Institute) (2013) ArcMap version 10.2. ESRI, Redlands, California
Funk C (2012) Rat Islands archaeological research 2003 and 2009: working toward an understanding of regional cultural, and environmental histories. Arctic Anthropol 48:25–51
Gibson DD, Byrd GV (2007) Birds of the Aleutian Islands. Nuttal Ornithological Club, American Ornighologists’ Union, Cambridge
Howald GR, Mineau P, Elliott JE, Cheng KM (2000) Brodifacoum poisoning of avian scavengers during rat control on a seabird colony. Ecotoxicology 8:431–447
Howald G, Donlan CJ, Galvan JP, Russell JC, Parkes J, Samaniego A, Wang Y, Veitch D, Genovesi P, Pascal M, Saunders A, Tershy B (2007) Invasive rodent eradication on islands. Conserv Biol 21:1258–1268
Howald G, Donlan CJ, Faulkner KR, Ortega S, Gellerman H, Croll DA, Tershy BR (2009) Eradication of black rats Rattus rattus from Anacapa Island. Oryx 44:30–40
Jones H (2010) Seabird islands take mere decades to recover following rat eradication. Ecol Appl 20:2075–2080
Jones HP, Tershy BR, Zavaleta ES, Croll DA, Keitt BS, Finkelstein ME, Howald GR (2008) Severity of the effects of invasive rats on seabirds: a global review. Conserv Biol 22:16–26
Kappes PJ, Jones HP (2014) Integrating seabird restoration and mammal eradication programs on islands to maximize conservation gains. Biodivers Conserv 23:503–509
Keitt B, Campbell K, Saunders A, Clout M, Wang Y, Heinz R, Newton K, Tershy B (2011) The Global Islands Invasive Vertebrate Eradication Database: a tool to improve and facilitate restoration of island ecosystems. In: Veitch DR, Clout MN, Towns DR (eds) Island invasives: eradication and management. Internation Union for the Conservation of Nature, Gland, pp 74–77
Kurle CM, Croll DA, Tershy BR (2008) Introduced rats indirectly change marine rocky intertidal communities from algae- to invertebrate-dominated. Proc Natl Acad Sci USA 105:3800–3804
Lavers JL, Wilcox C, Donlan CJ (2010) Bird demographic responses to predator removal programs. Biol Invasions 12:3839–3859
Loehle C, Eschenbach W (2012) Historical bird and terrestrial mammal extinction rates and causes. Divers Distrib 18:84–91
Lorvelec O, Pascal M (2005) French attempts to eradicate non-indigenous mammals and their consequences for native biota. Biol Invasions 7:135–140
Major HL, Jones IL, Byrd GV, Williams JC (2006) Assessing the effects of introduced Norway rats (Rattus norvegicus) on survival and productivity of Least Auklets (Aethia pusilla). Auk 123:681–694
Maron JL, Estes J, Croll DA, Danner EM, Elmendorf SC, Buckelew SL (2006) An introduced predator alters Aleutian Island plant communities by thwarting nutrient subsidies. Ecol Monogr 76:3–24
Paul E, Salmon T (2010) The Rat Island eradication project: a critical evaluation of nontarget mortality. The Ornithological Council, Bethesda
Russell JC (2011) Indirect effects of introduced predators on seabird islands. In: Mulder CP, Anderson WP, Towns DR, Bellingham PJ (eds) Seabird islands ecology, invasion, and restoration. Oxford University Press, New York, pp 261–279
Tershy BR, Shen KW, Newton KM, Holmes ND, Croll DA (2015) The importance of islands for the protection of biological and linguistic diversity. Bioscience 65:592–597
Towns DR (2008) Eradications as reverse invasions: lessons from Pacific rat (Rattus exulans) removals on New Zealand islands. Biol Invasions 11:1719–1733
Towns DR, Atkinson IAE, Daugherty CH (2006) Have the harmful effects of introduced rats on islands been exaggerated? Biol Invasions 8:863–891
Towns DR, Byrd GV, Jones HP, Rauzon MJ, Russell JC, Wilcox C (2011) Impacts of introduced predators on seabirds. In: Mulder CP, Anderson WP, Towns DR, Bellingham PJ (eds) Seabird islands ecology, invasion, and restoration. Oxford University Press, New York, pp 56–90
US Board on Geographic Names (2012) Geographic Names Post Phase I Board/Staff Revisions. 10 May 2012
US Fish and Wildlife Service (2007) Restoring wildlife habitat on Rat Island environmental assessment. Alaska Maritime National Wildlife Refuge, Aleutian Islands Unit, Alaska
Acknowledgments
We gratefully acknowledge field assistance provided by S. Abel, E. Bishop, A. Chateau, K. Cunningham, R. Dingler, S. Ebbert, C. Eggelston, R. Federer, J. Giffard, C. Hanson, R. Heinz, J. Helm, E. McCreless, A. McInturff, M. Mumm, K. Outten, R. Shaening-Pokrasso, and R. Stansbury. Alaska Maritime National Wildlife Refuge Staff, particularly V. Byrd, A. Sowls, S. Ebbert, G. Siekaniec and the crew of the R/V Tiglax provided invaluable research support. We were supported in part by the National Science Foundation (OPP-9985814), the National Fish and Wildlife Foundation (0101.12.030733) and the David and Lucile Packard Foundation. The views and conclusions contained in this document are those of the authors and should not be interpreted as representing the opinions or policies of the National Fish and Wildlife Foundation. Mention of trade names or commercial products does not constitute their endorsement by the National Fish and Wildlife Foundation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Croll, D.A., Newton, K.M., McKown, M. et al. Passive recovery of an island bird community after rodent eradication. Biol Invasions 18, 703–715 (2016). https://doi.org/10.1007/s10530-015-1042-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-015-1042-9