
Abstract
Biological invasions are a fixture in our landscapes, with consequent losses in endemic biota and shifts in ecosystem function. Despite the historical recognition of exotic species success in novel environs, this phenomenon lacks a holistic-descriptive framework. Recent attempts to explain biological invasions are based largely on identifying the inherent invasive qualities of successful exotic species (i.e., invasiveness), or characterizing the susceptibility of a habitat to an introduced species (i.e., invasibility), with few studies examining their interaction or additional contributing factors (e.g., time since introduction). We propose unifying the ’points of entry’ into biological invasions with a state factor model that incorporates all contributing variables—not just species or habitats—into a quantifiable, factorial model amenable to hypothesis testing. State factors are phenomenological variables describing the state of a system—historically used in soil and vegetation science. Our state factor equation relates any quantifiable property of an invasion (i) as a function of propagule pressure (p), introduced habitat (h), invader autecology (a), source environment (s), and time since introduction (t). By manipulating state factors singly, or in interaction, targeted variation can be related to quantifiable properties of exotic species while controlling, or at minimum accounting for, remaining factors contributing variation to the system. This holistic factor-function paradigm extends research on invasions from beyond the limits imposed by current theory, fosters novel empirical approaches, elucidates knowledge gaps in our understanding of resident invasions, and allows for variable accounting via a factor matrix. Here we briefly outline the ontogeny of state factors in soil and vegetation science, detail our proposed ’phast’ framework for biological invasions, including notation, and examine a case study in state factor utility.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Despite the near ubiquity of invasive species across our landscapes, a universal model describing the success of exotic species remains remarkably elusive. Most efforts to explain the performance of invading plants and animals range from chronicling species traits leading to their invasiveness, to attempts to correlate habitat characteristics with susceptibility to invasion. This has historically been a piecemeal effort where (non)additivity of the components is unknown (Agrawal et al. 2007), because of their (mostly) singular focus. Some have attempted to unify the field by focusing on specific commonalities among biological invasions (e.g., Colautti et al. 2006; Hallett 2006), but lack broad explanatory power because of their limited inclusion. The progression from rare introduction to occasional exotic to ubiquitous nuisance is the exceptional result of historical contingencies among many interacting variables, each of which played a role in the invasion sequence—resulting in a complex system.
Historically, when faced with complex systems we as researchers often impose operationally defined boundaries and reduce a system to simpler components more readily accessible to experimentation. Whether defining an ecosystem, succession, or soil development, we draw artificial boundaries (usually spatial or temporal) through what is in actuality a continuum, thereby defining a unit of analysis, which can be observed or manipulated experimentally.
Vasilii Dokuchaev performed just this exercise in the 1870s when given the task of describing the structure, origin, and evolution of the Russian chernozem, a deep organic-rich soil common to western Russia (Krupenikov 1992). Prior to this, a coherent soil classification system had been elusive, largely a result of the historical view of soils as the product exclusively of "weathering" and not possessing emergent properties of their own. After traversing 10,000 km of the chernozem belt, Dokuchaev stated, "soil exists as an independent body with a specific physiognomy, has its own special origin, and properties unique to it alone" (Dokuchaev 1949–1961, p. 245). He further formulated that soils are "continuously being formed as a result of the constant interaction of the following agents: living and dead organisms (such as plants and animals), parent rock, climate, and relief of the locality" (Dokuchaev 1949–1961). Dokuchaev’s truly radical proposition that the earth’s surface is composed of a systematic matrix of soil types, which are a product of more than merely moisture and temperature—actually five soil-forming factors (climate, parent material, organisms, topography, and time)—would ultimately revolutionize not only soil science (earning him the title of ’Father of Soil Science’), but geology, geography, and ecology as well.
Regrettably, Dokuchaev’s idea languished in the Russian literature, largely unavailable to a western audience for decades. It was not until the publication of Hans Jenny’s seminal book Factors of Soil Formation: A System of Quantitative Pedology (Jenny 1941), that the concept of state factors, and the processes involved in soil genesis were brought to the fore. (Other soil scientists had adopted the soil-forming factor mindset pre-Jenny, but these are beyond the scope of this manuscript. Nevertheless, it was the presentation and quantitative examples in Factors that initiated the revolution in soil science.)
Jenny includes the same five state factors as Dokuchaev, but relates them into a quantitative equation:
where s is any quantifiable soil property (e.g., water holding capacity, pH, porosity) as a function of climate (cl), potential biota (o), topography (r), parent material (p), and time (t), with ellipses added for future addition of factors (Jenny 1941). Embracing the elegant, yet daunting prospect of using his ’factor-function paradigm,’ Jenny states that the "soil formation [equation] is of little value unless it is solved" (Jenny 1941). To this end, Jenny employs differential equations to visualize the state factor model for soil properties:
This representation allows each state factor to be isolated, and explicitly shows that a change in one factor results in changes in the dependent variable. The remainder of Factors is devoted to detailed, quantitative examples demonstrating the relationship between the state factors and soil properties, much as The Origin of Species (Darwin 1859) includes prolific examples to support the mechanism of natural selection.
One of Jenny’s doctoral students, Jack Major, went on to expand the 'clorpt’ model to ecosystems, concluding the same five state factors described properties of vegetation (Major 1951). Both Jenny and Major recognized that no state factor could be reduced to a single number representing the entire variation comprising that factor, nor did they propose this framework as a predictive model. Rather, by recognizing all variables that comprise each state factor, more efficient and informative experiments could be designed to elucidate the relationship between the state factors and the properties of interest. By manipulating specific components of the target state factor, while holding all other factors not being tested as invariable as possible, relationships between properties of the system and the state factor of interest can be empirically described. Both researchers were relegated to locating sequences in the landscape that varied in one factor, while the others remained negligibly variable (e.g., chronosequence). Soil and vegetation properties could now be investigated empirically, quantitatively, and holistically. By drawing operationally defined boundaries around their system and reducing it into component parts (i.e., state factors) via a ‘factor-function paradigm’, Jenny and Major brought the complexity of soils and vegetation into the quantitative realm that continues to inspire empirical research, even as the origins of the approach have been forgotten. Can this same paradigmatic factor-function template be applied to the complex field of biological invasions? To address this question we first need to survey the current ‘state of the field.’
Current hypotheses on biological invasions
In addition to inspiring notions of evolution’s underlying mechanism, Darwin’s Beagle voyage engendered this lament on exotic species success, “many species, naturalized through man’s agency, have spread with astonishing rapidity over new countries” (Darwin 1859). Despite the historical recognition of biological invasions, and the body of existing data on myriad aspects of introduced plants and animals, we have yet to synthesize this information into an integrated framework capable of describing, let alone predicting, biological invasions (e.g., Hallett 2006). The difficulty is rooted in the complex and varied factors that have been demonstrated to contribute to the success or failure of an introduced species, as well as the spatial and temporal scales at which these factors operate (Mack et al. 2000). Research on biological invasions has spanned aspects including species traits (e.g., Buckley et al. 2003), habitat properties (e.g., Huenneke et al. 1990), evolutionary processes (e.g., Blair and Wolfe 2004), biogeographical trends (e.g., Grigulis et al. 2001), propagule pressure (e.g., Ahlroth et al. 2003), as well as the interactions among several factors (e.g., Barney et al. 2005; Chong et al. 2006; Von Holle and Simberloff 2005). Despite the varied aspects contributing to invasion success, the overwhelming body of theory can be grouped into two broad categories—those premised on species autecology and those focusing on habitat characteristics (cogently reviewed in Hierro et al. 2005). Admittedly, there are exceptions (e.g., predictions of invasive species; Reichard and Hamilton 1997), but the majority of both theoretical and empirical work in the descriptive/mechanistic realm are either species or habitat-focused.
The leading hypotheses based on species autecology in the context of the introduced habitat are the Enemy Release Hypothesis (ERH) (Keane and Crawley 2002), Evolution of Increased Competitive Ability Hypothesis (EICA) (Blossey and Notzold 1995), and the Novel Weapons Hypothesis (NWH) (Callaway and Aschehoug 2000). ERH and EICA are premised on the existence of co-evolved herbivores and pathogens in the native range, which are absent or greatly reduced in the introduced range (Mitchell and Power 2003). This ‘release’ from specialist natural enemies in a new habitat results in a shift of defense resources into biomass (EICA), or a relative advantage of the invader against its resident competitors who are exposed to their natural enemies (ERH). NWH is grounded in the co-evolutionary history of communities—competitors in the native range are ‘immune’ to allelochemicals, while a naïve community may be susceptible to an exotic compound released by the introduced plant, conferring a competitive advantage to the exotic species. An additional hypothesis, not based on species autecology per se, but the dynamics of the introduction, posits that invasions are contingent on the number and magnitude of release events into a non-native range—propagule pressure (PP) (Lockwood et al. 2005). Each hypothesis is grounded in a biogeographic comparison within the context of the species’ autecology (ERH, EICA, NWH), or the flux of disseminules into the introduced range (PP).
The second cadre of hypotheses is concerned exclusively with the habitat into which the exotic species is introduced. These include the Biotic Resistance Hypothesis (BRH) (aka Species Richness, Elton 1958), the Fluctuating Resource Hypothesis (FRH) (Davis et al. 2000), and the Disturbance Hypothesis (DH) (Hobbs and Huenneke 1992) and deal with the susceptibility of a habitat to invasion by a non-native species—invasibility. BRH relates native species richness to invasibility, and has been reviewed extensively in the literature (e.g., Shea and Chesson 2002). Some studies have shown a positive correlation between native species richness and invasibility, while others have demonstrated a negative trend (Byers and Noonburg 2003)—a paradox attributed to the spatial scale of the experiment (Fridley et al. 2007). Unlike BRH, FRH is not focused on the resident vegetation, but rather on the availability of resources: invasibility increases proportionally with an increase in available resources. Lastly, DH posits that invasive species are better adapted to disturbance, natural or anthropogenic, with habitat invasibility increasing with increasing disturbance frequency or magnitude. In summary, this suite of hypotheses describes the susceptibility of a habitat to an introduced species based on resident vegetation, resources, or disturbance.
The merits of the above hypotheses, and their derivatives, are many, as indicated by the vast literature testing them. However, no single hypothesis captures the complexity of all contributing phenomena— largely a result of their singular focus. Evidence exists supporting and refuting each hypothesis (for example see Bossdorf et al. 2005 for review of studies testing EICA), which demonstrates that no ’theory of everything’ has yet been proposed. In addition, no system has been studied holistically. For example, research on the invasion of knapweed species (Centaurea spp.) into the western US has focused largely on plant community interactions mediated by chemical exudates (e.g., Callaway and Aschehoug 2000; Hierro and Callaway 2003), while island invasibility research has focused on physical or abiotic factors like island size and distance from source (e.g., MacCarthur and Wilson 1967; Lonsdale 1999). Thus, hypotheses based on habitat invasibility (BRH, FRH, DH) essentially ignore salient features of the introduced species, while hypotheses based largely on autecological traits (ERH, EICA, NWH) ignore characteristics of the habitat being invaded. We propose that a first step toward unifying these discrete approaches should be to view biological invasions as Jenny did with soils and Major with vegetation: a complex system with unique properties dependent on several contributing factors.
State factors in biological invasions
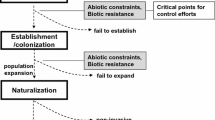
A successful invasion into previously uncolonized environs is the culmination of a contingent history integrating numerous variables, which contributed individually and in aggregate. As a consequence, the science investigating biological invasions is a post hoc enterprise, because an invasion is not discernable until the exotic species has reached invasive proportions or is causing noticeable environmental damage. Therefore, the hypotheses outlined above are directed at identifying and empirically investigating parcels of the conditional history—typically centered on the invading species or the invaded habitat. In contrast, we propose that each biological invasion—from a novel genotype (sensu Saltonstall 2002) to a novel species—should be viewed as a unique ‘entity’ whose properties depend on all interacting factors comprising the contingent history of the invasion, which can be represented nonetheless holistically instead of individually.
Using the precedence of the hypotheses outlined above, and the vast literature on biological invasions, we have categorized the interacting variables contributing to a biological invasion—plant or animal—into five broadly-defined state factors: propagule pressure (p), properties of the introduced habitat (h), invading species (or genotype) autecology (a), properties of the source habitat (s), and time since introduction (t). The state factors are related via the factor-function:
where i is any quantifiable property of an introduced population, and p, h, a, s, and t are operationally defined in Table 1.
Each state factor comprises manifold sub-variables, just as did Jenny’s and Major’s clorpt factor-function. For example, our introduced habitat factor (h) incorporates their climate (cl), biota (o), topography (r), and parent material (p) state factors among many other sub-variables (Table 1). Complexity is therefore built into the factor-function, but allows an elegant accounting of contributing variation without granting importance to any particular factor or sub-variable. Indeed, the contribution to i of some state factors may far outweigh that of others (e.g., propagule pressure in bird invasions; Cassey et al. 2005). However, such a weighting would only be meaningful with a robust dataset on species introductions (failed and successful), introduction loci, biotic and abiotic conditions, propagule number and events, source populations, and time of introduction. Such data are virtually non-existent (Lonsdale 1999). This fact however, does not relegate state factor utility to merely a qualitative academic exercise. Importantly, Jenny stressed that his state factor equation could be used quantitatively via clever site selection whereby only the state factors of interest varied and all others where nearly invariant (Jenny 1941). Unlike studies of soils pre-Jenny and vegetation pre-Major, research on biological invasions has been largely quantitative, and amenable to experimental manipulation. This raises the question: Is a state factor approach to biological invasions amenable to quantitative-based hypothesis testing?
As stated above, the descriptive/mechanistic hypotheses regarding biological invasions posited to date do account for a great deal of the variation in properties of biological invasions. However, most empirical research conducted under the framework of these hypotheses are limited in extent, leaving the many factors demonstrated in other studies to be important in biological invasions unaccounted (Table 2). For example, the Evolution of Increased Competitive Ability Hypothesis predicts that a reduction in biotic pressure in the introduced range (h) confers an advantage to genotypes that can reallocate unneeded defense resources (a) to yield more competitive phenotypes. However, there is no accounting of propagule pressure (p), the genetic variance of introduced propagules (a), whether the introduced propagules are environmentally tolerant of the novel environment (s,h), nor the number of generations (t) necessary for this novel phenotype to emerge. Likewise, the Fluctuating Resource Hypothesis states that habitat invasibility is proportional to resource availability in the introduced range (h), regardless of the species being introduced (a), its environmental tolerance (s), whether one or 10 million propagules are introduced (p), or the time elapsed (t). Mitchell et al. (2006) attempt to integrate all biotic-related features of the invasion process into a single working hypothesis, but important aspects of invasion success—abiotic factors, propagule pressure, and time—were not considered. Propagule pressure has been stated as being a primary determinant in the establishment of invasive species (Cassey et al. 2005; Colautti et al. 2006; Lockwood et al. 2005), but is not explicitly accounted for in any leading theory on biological invasions (Table 2). Perhaps the most neglected aspect in biological invasions is time since introduction. It is well documented that most invasions progress through a lag phase followed by exponential range expansion (Kowarik 1995). Therefore, properties of the invasion (i) are strongly dependent on the current stage of the invasion (t), which is almost never documented (though see Carpenter and Cappuccino 2005). A trend emerges when viewing the historical perspective on biological invasions through the lens of the proposed factor-function paradigm—a patent neglect in accounting for contributing variables.
The deficiency in general explanatory power of proposed theories is not their lack of utility or validity, but rather is rooted in their singularity and consequent lack of generalizability. These theories all generate testable hypotheses and make general predictions, but lack even tacit accounting of alternative mechanisms (i.e., state factors) that might be involved. It should be noted that the authors of several of these theories conclude their manuscripts with the recognition that other mechanisms may be important. For example, Davis et al. (2000) in their Fluctuating Resource paper state "whether or not invasion actually occurs in a particular environment depends also on propagule pressure and the attributes of the invading species." Yet they offer no way to incorporate this admission with their “quantitative, analytic, and systematic” theory (Davis et al. 2000). While we intend no disparagement of the authors or the explanatory power the various invasion hypotheses afford, we feel a need to go beyond conservative hedges like this if the field of invasion biology is to advance. We suggest that the use of the state factor model can unify current knowledge, and aid in the design of more effective experiments that explicitly address all contributing variables in the invasion process in a holistic manner.
Garlic mustard as a case study
To demonstrate how the proposed state factor framework can be used to organize research, elucidate knowledge gaps, and facilitate meta-analyses, we catalogued all studies (29 papers) investigating the biennial forest understory invader garlic mustard (Alliaria petiolata(M. Bieb.) Cavara and Grande) (Note: studies focused on management have been intentionally excluded as they do not investigate properties of the invasion per se). Garlic mustard was chosen as it has a widespread distribution in the Northeast and Midwestern US, is considered one of the primary threats to hardwood forest ecosystems (Nuzzo 1999), and a substantial body of research has and continues to be conducted on this species. This list encompasses both non-manipulative observational (e.g., demography of established populations) and experimental (e.g., light or nutrient manipulation) studies of both single and multiple state factors (Appendix 1). Through analysis of the objectives of the experiment and the methods used, we determined a posteriori which invasion properties were under investigation (i—dependent variables), and which state factors (phast) were manipulated. However, we were largely unable to determine which state factors were held constant and which were simply unknown (see below for proposed notation). Despite the potential limitations of this post-hoc analysis, we were able to determine which state factors have been deemed most important, and where gaps in knowledge occur (Table 3). The factor matrix in Table 3 shows that a vast majority of the studies reviewed focused on garlic mustard autecology (a) and habitat properties (h), which is not surprising considering the root of current hypotheses advanced to explain biological invasions (Table 2). Interestingly, despite the plethora of studies on this species, no single study has explicitly investigated propagule pressure (p) or time since introduction (t) (Table 3). Garlic mustard has been present in North America since at least the 1860s, with multiple introduction events (Nuzzo 1993; Durka et al. 2005). The number of introduction events, their spatial and temporal distribution (p), and the time since introduction (t) can profoundly influence the success and properties of a nascent invasion (e.g., Barney 2006). Without considering the influence of all state factors, information on garlic mustard biology, population dynamics, and plant-plant interactions is incomplete and difficult to reconcile.
This simple exercise on one the most researched invaders elucidates a need for a novel way of approaching biological invasions that is analytical and holistic. Lucid identification of ‘knowledge gaps’ in current understanding develops when using a factor matrix (e.g., Table 3), which can direct future research to address areas of important, but poorly studied, contributing variables. This post hoc analysis also distills current knowledge into a format that is easily manipulated for meta-analysis purposes to identify large-scale trends that can aid in management and invasion mitigation. Additionally, metrics in a robust factor matrix (i.e., one with many sub-variables known for each state factor) could be analyzed via multi-variant analyses to determine factor importance (e.g., Reichard and Hamilton 1997; Rejmanek and Richardson 1996). Here we addressed how factor-function analyses can be done post hoc on published research, next we address using this approach in designing experiments and integrating results.
State factor utility
Recent calls for investigating biological invasions in a biogeographic context (Meyerson and Mooney 2007; Hierro et al. 2005), and standardization and sharing of information (Rice 2007), are excellent starting points for advancing the science of invasion ecology in a global context. However, this advancement is hampered by issues of spatial scale (Mack et al. 2007), and an incomplete understanding of the biotic and abiotic factors that structure communities singly and in interaction (Agrawal et al. 2007). The state factor framework provides an opportunity to address biological invasions holistically by accounting for all variables involved (sensu Jenny 1980). Additionally, the factor-function framework allows the opportunity to draw our ‘research boundary’ at any spatial (from individuals to continents) or temporal (generations to centuries) scale we choose (Mack et al. 2007). However, the utility of this approach is not an attempt to address the infinite variables comprising the contingent history of the invasion resulting in a single value for i—a futile enterprise to be sure. Nor is it assumed that all state factors can be tested in a single factorial uberexperiment. Rather, the factorial model should be used as a tool to design efficient experiments that investigate contributing variables comprising the state factors, while accounting for other important, but not manipulated variables. For example, to examine the existence of a phenotype unique to those found in the native range (i.e., invasive phenotype), one experiment would be to compete individuals from populations originating in the native and introduced ranges (s) against a common competitor (h) in a common garden (thereby holding all other aspects of h invariable), which can be represented functionally as:
where i are the dependent variables, s and h are the state factors under factorial manipulation, a and p are the state factors held invariable, and t (subscript in brackets) is unknown. This equation is then followed with details of each variable, as shown in Appendix 1. This explicit, functional representation of the research undertaken will provide editors, reviewers, and readers a quick overview of the properties investigated in relation to specific variables, as well as those variables that were unknown or not considered. The factor-function approach allows clear relationships between manipulated state factors and properties of the invasion to be elucidated, much as Jenny envisioned with soils via generating "sequences" (Jenny 1941). Jenny was constrained to locating naturally occurring sequences to address relationships between soil properties and single state factors (e.g., Jenny 1946), while invasion biologists can design experimental systems virtually at will to address relationships of interest.
Potential interdependence of factors in Jenny’s clorpt equation plagued the pedologist (e.g., plants affect soil, and soil affects plants). For this reason Jenny stressed that identifying sequences in the landscape (e.g., toposequence) where one factor varied and the other four were nearly invariant, or where the magnitude of their influence was negligible (Jenny 1980). Our proposed ‘phast’ model is less prone to problems of interdependence because we can manipulate state variables factorially, while Jenny was constrained to single factor sequences. Additionally, the factors in phast are generally free of the circularity of cause-and-effect inherent in the factors involved in soil genesis. These are major advantages to the factor-function approach in the study of biological invasions that was unavailable to Jenny and his successors. Through factorial manipulation of the state factors of interest, their relationship (i.e., interaction) to the properties being quantified can be identified. In the example above, the researchers are able to identify the relationship between source (native versus introduced, and population), competitive ability, and fitness (whatever metrics they quantified). This experiment could then be complemented with investigations into the effects of local propagule pressure and exudation of allelopathic compounds on population dynamics of the invader, which is followed by a study of habitat disturbance, etc. Each experiment in the hierarchy complements and builds on the previous. Results from all experiments are entered into a factor matrix (sensu Table 3), which serves as an account sheet that directs future research while affording opportunity for meta-analyses for determining trends across factors and spatial and temporal scales. A holistic picture of the invasion emerges via the factor-function paradigm.
Summary
Complex systems, such as soils and vegetation, have been successfully reduced to component variables—state factors—which are determinants of the properties of the target system. Adoption of the state factor paradigm ushered soils and vegetation science into the realm of quantitative disciplines amenable to hypothesis testing. Our current understanding of biological invasions suffers from our inability to account for all contributing factors in a holistic perspective—partially a result of the leading hypotheses and their ontogeny. Therefore, we propose implementation of a state factor model of biological invasions that integrates all identified aspects of the invasion process—propagule pressure (p), introduced habitat characteristics (h), species autecology (a), source habitat environment (s), and time since introduction (t)—into a factorial, quantitative equation. Though we have focused on plant introductions, adoption of this approach will allow explicit recognition of all variables involved in describing the properties of an introduced species, plant or animal, elucidation of gaps in current knowledge, and the recognition of not being able to control for all variables (e.g., time since introduction), but acknowledging their involvement. A state factor approach is fully amenable to testing current theories on biological invasions, as well as for the generation of future hypotheses. The greatest benefit to the field of invasion biology is the experimental recognition of all contributing variables—through utilization of the state factor framework and accompanying factor matrix. Just as soil and vegetation science languished without identification and manipulation of the phenomenological state factors and their relation to system properties, the study of biological invasions cannot progress without making full use of this holistic approach that incorporates all contributing variation rather than focusing on singular aspects.
Abbreviations
- BRH:
-
Biotic Resistance Hypothesis
- DH:
-
Disturbance Hypothesis
- EICA:
-
Evolution of Increased Competitive Ability Hypothesis
- ERH:
-
Enemy Release Hypothesis
- FRH:
-
Fluctuating Resource Hypothesis
- NWH:
-
Novel Weapons Hypothesis
- PP:
-
Propagule Pressure
References
Agrawal AA, Ackerley DD, Adler F, et al (2007) Filling key gaps in population and community ecology. Front Ecol Environ 5:145–152
Ahlroth P, Alatalo RV, Holopainen A et al (2003) Founder population size and number of source populations enhance colonization success in waterstriders. Oecologia 137:617–620
Barney JN, DiTommaso A, Weston LA (2005) Differences in invasibility of two contrasting habitats and invasiveness of two mugwort (Artemisia vulgaris) populations. J Appl Ecol 42:567–576
Barney J (2006) North American history of two invasive plant species: phytogeographic distribution, dispersal vectors, and multiple introductions. Biol Invasions 8:703–717
Blair AC, Wolfe LM (2004) The evolution of an invasive plant: an experimental study with Silene latifolia. Ecology 85:3035–3042
Blossey B, Notzold R (1995) Evolution of increased competitive ability in invasive nonindigenous plants: a hypothesis. J Ecol 83:887–889
Bossdorf O, Augue H, Lafuma L et al (2005) Phenotypic and genotypic differentiation between native and introduced plant populations. Oecologia 144:1–11
Buckley YM, Downey PS, Fowler SV et al (2003) Are invasives bigger? A global study of seed size variation in two invasive shrubs. Ecology 84:1434–1440
Byers JE, Noonburg EG (2003) Scale dependent effects of biotic resistance to biological invasion. Ecology 84:1428–1433
Callaway RM, Aschehoug ET (2000) Invasive plants versus their new and old neighbors: a mechanism for exotic invasion. Science 290:521–523
Carpenter D, Cappuccino N (2005) Herbivory, time since introduction and the invasiveness of exotic plants. J Ecol 93:315–321
Cassey P, Blackburn TM, Duncan RP et al (2005) Lessons from the establishment of exotic species: a meta-analytical case study using birds. J Anim Ecol 74:250–258
Chong GW, Otsuki Y, Stohlgren TJ et al (2006) Evaluating plant invasions from both habitat and plant perspectives. West North Am Nat 66:92–105
Colautti RI, Grigorivich IA, MacIsaac HJ (2006) Propagule pressure: a null model for biological invasions. Biol Invasions 8:1023–1037
Darwin C (1859) On the origin of species. John Murray, London, England
Davis MA, Grime JP, Thompson K (2000) Fluctuating resources in plant communities: a general theory of invasibility. J Ecol 88:528–534
Dokuchaev VV (1949–1961) Socheneniya. in. Moskva, Izd-vo Akademii nauk SSSR, Moscow
Durka W, Bossdorf O, Prati D, Auge H (2005) Molecular evidence for multiple introductions of garlic mustard (Alliaria petiolata, Brassicaceae) to North America. Mol Ecol 14:1697–1706
Elton CS (1958) The ecology of invasions by animals and plants. Methuen, London, England
Fridley JD, Stachowicz JJ, Naeem S et al (2007) The invasion paradox: reconciling pattern and process in species invasion. Ecology 88:3–17
Grigulis K, Sheppard AW, Ash JE et al (2001) The comparative demography of the pasture weed Echium plantagineum between its native and invaded ranges. J Appl Ecol 38:281–290
Hallett SG (2006) Dislocation from coevolved relationships: a unifying theory for plant invasion and naturalization? Weed Sci 54:282–290
Hierro JL, Callaway RM (2003) Allelopathy and exotic plant invasion. Plant Soil 256:29–39
Hierro JL, Maron JL, Callaway RM (2005) A biogeographical approach to plant invasions: the importance of studying exotics in their introduced and native range. J Ecol 93:5–15
Hobbs RJ, Huenneke LF (1992) Disturbance, diversity, and invasion: implications for conservation. Conserv Biol 6:324–337
Huenneke LF, Hamburg SP, Koide R et al (1990) Effects of soil resources on plant invasion and community structure in a California serpentine grassland. Ecology 71:478–491
Jenny H (1941) Factors of soil formation: a system of quantitative pedology. McGraw-Hill, New York
Jenny H (1946) Arrangement of soil series and types according to functions of soil-forming factors. Soil Sci 61:375–391
Jenny H (1980) The soil resource: origin and behavior. Springer-Verlag, New York
Keane RM, Crawley MJ (2002) Exotic plant invasions and the enemy release hypothesis. Trends Ecol Evol 17:164–170
Kowarik I (1995) Time lags in biological invasions with regard to the success and failure of alien species. In: Pyesk P, Prach K, Rejmanek M, Wade M (eds) Plant invasions–general aspects and special problems. Academic Publishing, Amsterdam
Krupenikov IA (1992) History of soil science: from its inception to the present. Amerind Publishing Co, New Delhi
Lockwood JL, Cassey P, Blackburn T (2005) The role of propagule pressure in explaining species invasions. Trends Ecol Evol 20:223–228
Lonsdale WM (1999) Global patterns of plant invasions and the concept of invasibility. Ecology 80:1522–1536
MacCarthur RH, Wilson EO (1967) The theory of island biogeography. Princeton University Press, Princeton
Mack RN, Simberloff D, Lonsdale WM, et al (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10:689–710
Mack RN, Von Holle B, Meyerson LA (2007) Assessing invasive alien species across multiple spatial scales: working globally and locally. Front Ecol Environ 5:217–220
Major J (1951) A functional, factorial approach to plant ecology. Ecology 32:392–412
Meyerson LA, Mooney MA (2007) Invasive alien species in an era of globalization. Front Ecol Environ 5:199–208
Mitchell CE, Agrawal AA, Bever JD et al (2006) Biotic interactions and plant invasions. Ecol Lett 9:726–740
Mitchell CG, Power AG (2003) Release of invasive plants from fungal and viral pathogens. Nature 421:625–627
Nuzzo V (1993) Current and historic distribution of garlic mustard (Alliaria petiolata) in Illinois. Mich Bot 32:23–34
Nuzzo V (1999) Invasion pattern of herb garlic mustard (Alliaria petiolata) in high quality forests. Biol Invasions 1:169–179
Reichard SH, Hamilton CW (1997) Predicting invasions of woody plants introduced into North America. Conserv Biol 11:193–203
Rejmanek M, Richardson DM (1996) What attributes make some plant species more invasive? Ecology 77:1655–1660
Rice B (2007) Invasive species data applications and data sharing across the Americas. Front Ecol Environ 5:W15–W16
Saltonstall K (2002) Cryptic invasion by a non-native genotype of the common reed, Phragmites australis, into North America. Proc Nat Acad Sci 99:2445–2449
Shea K, Chesson P (2002) Community ecology theory as a framework for biological invasions. Trends Ecol Evol 17:170–176
Von Holle B, Simberloff D (2005) Ecological resistance to biological invasion overwhelmed by propagule pressure. Ecology 86:3212–3218
References to Appendix 1
Anderson RC, Dhillion SS, Kelley TM (1996) Aspects of the ecology of an invasive plant, garlic mustard (Alliaria petiolata), in Central Illinois. Rest Ecol 4:181–191
Bossdorf O, Prati D, Auge H, et al (2004a). Reduced competitive ability in an invasive plant. Ecol Lett 7:346–353
Bossdorf O, Schroder S, Prati D, et al (2004b) Palatability and tolerance to simulated herbivory in native and introduced populations of Alliaria petiolata (Brassicaceae). Am J Bot 91:856–862
Byers DL, Quinn JA (1998) Demographic variation in Alliaria petiolata (Brassicaceae) in four contrasting habitats. J Torrey Bot Soc 125:138–149
Cipollini D (2002) Variation in the expression of chemical defenses in Alliaria petiolata (Brassicaceae) in the field and common garden. Am J Bot 89:1422–1430
Cipollini D, Mbagwu J, Barto K, et al (2005) Expression of constitutive and inducible chemical defenses in native and invasive populations of Alliaria petiolata. J Chem Ecol 31:1255–1267
Dhillion SS, Anderson RC (1999) Growth and photosynthetic response of first-year garlic mustard (Alliaria petiolata) to varied irradiance. J Torrey Bot Soc 126:9–14
Durka W, Bossdorf O, Prati D, et al (2005) Molecular evidence for multiple introductions of garlic mustard (Alliaria petiolata, Brassicaceae) to North America. Mol Ecol 14:1697–1706
Lewis K, Bazzaz FA, Liao Q, et al (2006) Geographic patterns of herbivory and resource allocation to defense, growth, and reproduction in an invasive biennial, Alliaria petiolata. Oecologia 148:384–395
Meekins JF, McCarthy BC (1999) Competitive ability of Alliaria petiolata (garlic mustard, Brassicaceae), an invasive, nonindigenous forest herb. Inter J Plant Sci 160:743–752
Meekins JF, McCarthy BC (2000) Responses of the biennial forest herb Alliaria petiolata to variation in population density, nutrient addition and light availability. J Ecol 88:447–463
Meekins JF, McCarthy BC (2001) Effect of environmental variations on the invasive success of a nonindigenous forest herb. Ecol Appl 11:1336–1348
Meekins JF, McCarthy BC (2002) Effect of population density on the demography of an invasive plant (Alliaria petiolata, Brassicaceae) population in a Southeastern Ohio forest. Am Midl Nat 147:256–278
Murphy SD (2005) Concurrent management of an exotic species and initial restoration efforts in forests. Rest Ecol 13:584–593
Myers C, Anderson RC (2003) Seasonal variation in photosynthetic rates influences success of an invasive plant, garlic mustard (Alliaria petiolata). Am Midl Nat 150:231–245
Myers C, Anderson RC, Byers DL (2005) Influence of shading on the growth and leaf photosynthesis of the invasive non-indigenous plant garlic mustard [Alliaria petiolata (M Bieb) Cavara and Grande] grown under simulated late-winter to mid-spring conditions. J Torrey Bot Soc 132:1–10
Prati D, Bossdorf O (2004) Allelopathic inhibition of germination by Alliaria petiolata(Brassicaceae). Am J Bot 91:285–288
Rebek KA, O’Neil RJ (2005) Impact of simulated herbivory on Alliaria petiolata survival, growth, and reproduction. Biol Control 34:283–289
Rebek KA, O’Neil RJ (2006) The effects of natural and manipulated density regimes on Alliaria petiolata survival, growth and reproduction. Weed Res 46:345–352
Renwick JAA, Zhang W, Haribal M, et al (2001) Dual chemical barriers protect a plant against different larval stages of an insect. J Chem Ecol 27:1575–1583
Roberts KJ, Anderson RC (2001) Effect of garlic mustard [Alliaria petiolata (Bieb Cavara & Grande)] extracts on plants and arbuscular mycorrhizal (AM) fungi. Am Midl Nat 146:146–152
Smith GR, Dingfelder HA, Vaala DA (2003) Effect of plant size and density on garlic mustard reproduction. Northeast Nat 10:269–276
Stinson KA, Campbell SA, Powell JR, et al (2006) Invasive plant suppresses the growth of native tree seedlings by disrupting belowground mutualisms. PLoS Biol 4:727–731
Susko DJ, Lovett-Doust L (1998) Variable patterns of seed maturation and abortion in Alliaria petiolata (Brassicaceae). Can J Bot 76:1677–1686
Susko DJ, Lovett-Doust L (1999) Effects of resource availability, and fruit and ovule position on components of fecundity in Alliaria petiolata (Brassicaceae). New Phytol 144:295–306
Susko DJ, Lovett-Doust L (2000a) Patterns of seed mass variation and their effects on seedling traits in Alliaria petiolata (Brassicaceae). Am J Bot 87:56–66
Susko DJ, Lovett-Doust L (2000b) Plant-size and fruit-position effects on reproductive allocation in Alliaria petiolata (Brassicaceae). Can J Bot 78:1398–1407
Vaughn SF, Berhow MA (1999) Allelochemicals isolated from tissues of the invasive weed garlic mustard (Alliaria petiolata). J Chem Ecol 25:2495–2504
Winterer J, Walsh MC, Poddar M, et al (2005) Spatial and temporal segregation of juvenile and mature garlic mustard plants (Alliaria petiolata) in a Central Pennsylvania woodland. Am Midl Nat 153:209–216
Acknowledgements
We would like to thank members of the DiTommaso and Blossey lab groups, and Michael Dosmann, Doug Soldat, and Jude Maul for being sounding boards and giving critical reviews of previous incarnations of our state factor concept and reviewing earlier drafts of this manuscript. We also would like to thank two reviewers for helpful suggestions. Special thanks to the late Jack Major for his inspiration.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Rights and permissions
About this article
Cite this article
Barney, J.N., Whitlow, T.H. A unifying framework for biological invasions: the state factor model. Biol Invasions 10, 259–272 (2008). https://doi.org/10.1007/s10530-007-9127-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-007-9127-8