
Abstract
Agrilus planipennis Fairmaire (Col: Buprestidae) (EAB) has been spreading rapidly throughout North America, killing millions of ash trees, Fraxinus spp. L. (Oleaceae). Eradication is not viable, so biological control using natural enemies is now the leading management strategy. Little information is available on whether native parasitoids and predators can be manipulated to increase EAB mortality. We moved freshly cut ash logs infested with EAB parasitoids to field sites where there was no record of EAB mortality by native North American natural enemies. Changes in EAB parasitism, EAB density, and woodpecker predation were monitored over the following three years. Higher parasitism of EAB by Phasgonophora sulcata Westwood (Hym: Chalcididae) (16.67 ± 16.67%) and Atanycolus spp. Foerster (Hym: Braconidae) (48.18 ± 4.29 %) was observed in all plots over three years after native parasitoids were released. However, no reduction was seen in EAB density between the treatment and control plots or over time. There was no significant relationship found between predation by woodpeckers and year or between control and parasitoid-release plots. However, woodpecker predation increased significantly with EAB density. Movement of ash logs containing native parasitoids to sites newly infested by EAB, but with low native natural enemy populations, can increase long-term EAB mortality as an added component in management strategies to slow its spread.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The emerald ash borer (EAB), Agrilus planipennis Fairmaire (Col: Buprestidae), is an invasive jewel beetle introduced into North America from Asia during the 1990s (Haack et al. 2002; Siegert et al. 2014). Since then, it has shown a continued pattern of spread in all directions from intial invasion and is now present in five Canadian provinces and 35 US states. EAB is considered one of the most damaging and costly insects to invade North American forests because of its ability to kill healthy ash trees, Fraxinus spp. L. (Oleaceae), with up to 99% tree mortality observed within a few years of arrival (Knight et al. 2013). Early management efforts involved quarantine strategies to isolate and eradicate known EAB populations, thereby limiting the spread of infested ash material (Cappaert et al. 2005). However, these were discontinued due to the beetle’s rapid spread, lack of funding, and the cryptic nature of the wood-boring larvae (Liu et al. 2003; Herms and McCullough 2014). Now, short-term management tools (e.g., quarantine of EAB-infested material, chemical treatment, tree removal, girdled “trap trees”) and long-term approaches such as biological control are being used to combat EAB in North America (Herms and McCullough 2014).
Although many abiotic and biotic mortality factors affect EAB populations, including natural host tree resistance (Anulewicz et al. 2007; Tannis and McCullough 2012), extreme cold (Crosthwaite et al. 2011), cold-warm-cold fluctuations (Sobek-Swant et al. 2012), predators (Rutledge et al. 2013; Jennings et al. 2015), parasitoids (Duan et al. 2009, 2012), and pathogens (Bauer et al. 2004; Kyei-Poku and Johny 2013), none have yet provided sufficient mortality to naturally suppress EAB. Thus, classical biological control has been proposed involving the introduction of non-native parasitoids from Asia where EAB is native, namely, Tetrastichus planipennisi Yang (Hym: Eulophidae), Oobius agrili Zhang and Huang (Hym: Encyrtidae), Spathius agrili Yang (Hym: Braconidae), and Spathius galinae Belokobylskij and Strazanac (Hym: Braconidae). Recent studies show that the introduction of several parasitoids, especially T. planipennisi, can cause significant EAB mortality (Duan et al. 2017). However, these co-evolved parasitoids alone have not yet suppressed EAB populations below damaging thresholds. There remains a need to explore additional biocontrol options for managing EAB spread throughout its introduced range in North America.
Augmenting native natural enemy populations where they are absent or in low numbers to combat introduced species such as EAB have been used against at least 32 species in Canada (MacQuarrie et al. 2013, 2016). North America is home to a rich complex of native natural enemies of Agrilus beetles, all with the potential to switch from native Agrilus Curtis (Col: Buprestidae) hosts to EAB, a closely-related, introduced species. Native avian predators, especially woodpeckers (Picidae) such as the hairy woodpecker [Picoides villosus (L.)], downy woodpecker [Picoides pubescens (L.)], and red-bellied woodpecker [Melanerpes carolinus (L.)], are known to increase in abundance after outbreaks of saproxylic beetles [i.e., mountain pine beetle, Dendroctonus ponderosae Hopkins (Col: Curculionidae)] and Asian longhorned beetle, Anoplophora glabripennis Motschulsky (Col: Cerambycidae) (Jiao et al. 2008; Edworthy et al. 2011). These three species have also been shown to attack EAB in North America with up to 95% mortality (Cappaert et al. 2005; Lindell et al. 2008; Jennings et al. 2013). Similarly, high rates of EAB parasitism by native wasps [i.e., up to 71% mortality by Atanycolus cappaerti Marsh and Strazanac (Hym: Braconidae) (Cappaert and McCullough 2009) and 40% mortality by Phasgonophora sulcata Westwood (Hym: Chalcididae) (Lyons 2010)] suggest that they could be augmented for biological control. This latter approach for managing EAB has received little attention in North America to date.
Successful augmentative biological control requires key information on the most effective method to distribute large quantities of natural enemies, optimal timing, and location for release, as well as factors affecting subsequent dispersal and monitoring. In forestry, augmentation of natural enemies has been explored only in a few forest systems. Notable examples include (1) mass release of the native egg parasitoid, Trichogramma minutum Riley (Hym: Trichogrammatidae), against outbreak populations of native spruce budworm, Choristoneura fumiferana (Clemens) (Lep: Tortricidae) in Canada (Smith 1996), and (2) mass release of native parasitoids against non-native Sirex woodwasps (Hym: Siricidae) in Australia (Haugen and Underdown 1990). The life-history traits of native parasitoids attacking EAB in North America are generally unfavourable for mass rearing and release, i.e., P. sulcata has a 1:1 parasitoid:host ratio (i.e., solitary) and is univoltine with a long generation time (Roscoe 2014). Thus, despite clearly showing promise, the only feasible way for native parasitoids to be augmented against EAB would be to move plant material containing parasitoids from the field to target sites with high EAB populations, similar to that used against Sirex (Hauden and Underdown 1990).
To date, proof of concept and criteria for long-term introduction, establishment, and spread of native parasitoids on EAB remains to be shown. Here, we assess the relative rate of increase in parasitism and its relationship to EAB density following augmentative biological control of native North American natural enemies. Specifically, we ask: (1) does EAB parasitism increase over time following augmentation?; (2) do all native parasitoids respond similarly?; (3) does EAB parasitism reduce EAB density?; and (4) how does predation by native woodpeckers impact EAB density or parasitism over the same time?
Materials and methods
Experimental design
In 2013, woodlots near the epicentre of the initial EAB invasion in southwestern Ontario, Canada were surveyed for a source of native parasitoids attacking EAB and suitable collection sites were identified using data on EAB ash-infested logs obtained by the Canadian Forest Service in 2012. Three collection sites were identified, two in Middlesex County, Ontario and another in Elgin County, Ontario, all which had ≥ 50% combined EAB parasitism by two native parasitoids, P. sulcata and Atanycolus spp. Foerster (Hym: Braconidae). Parasitoid release and non-release control plots were established in three mixed hardwood forests within Toronto, Ontario, Canada that had ≥ 25% ash trees with a diameter at breast height of 9.5 ± 1.0 cm and no record of EAB parasitism. Over the next three years (2014, 2015, and 2016), ash logs cut into 60 cm lengths from trees infested with native parasitoids were collected annually in early spring from the collection sites and transported to the release plots in Toronto. The release entailed placing the logs adjacent to living ash trees in each release plot, evenly divided so that each release plot received a similar surface area and quantity of EAB-parasitoid-infested material from each of the three collection sites. The control plots (without parasitoid-infested ash logs) were located at least 500 m away from the release plots based on known poor dispersal capacity of P. sulcata (Gaudon et al. 2018). Changes in EAB parasitism, density, and woodpecker predation were monitored across all plots.
Ash trees were sampled in 2013 before parasitoid-infested logs were transported to the sites, as well as one and three years after the initial transport. In 2013, we removed one to three logs per sample tree from two or three trees in each plot to quantify the number of native EAB parasitoids, number of EAB, and number of woodpecker holes in the log linked directly to EAB. In the following year (2014), we sampled fewer trees (i.e., one or two) per plot, removing more logs (i.e., two to four logs per tree) to conserve the number of living trees remaining in each plot. Due to the destructive nature of sampling and rapid decline of ash trees in these EAB-infested plots, a complete sample of trees in the parasitoid release and non-release control plots was not done in 2015, but rather all plots were examined to ensure that sufficient living ash and viable phloem was available for EAB and its parasitoids to reinfest. In the final sampling year (2016) two to four logs per tree were sampled from two to seven trees per plot. Upon removal from the field, logs were fully enclosed in emergence netting and hung on hooks in a rearing chamber at a mean temperature of 22.8 ± 0.5 °C and RH of 51.4 ± 2.9%. The length and diameter of both ends of all logs were measured, and a mean surface area calculated for each log.
The numbers of adult EAB and parasitoids emerging from each ash log were counted to determine the change in EAB density and parasitism between the parasitoid-release and control plots. Logs were then dissected after four months to include all EAB and parasitoids that had failed to emerge. EAB parasitism in the release plots was calculated for solitary parasitoids by dividing the total number of wasps by the sum of the total number of EAB plus wasps collected from the logs (Lyons 2010; Roscoe et al. 2016), and then compared similarly to the control plots.
Woodpecker predation was determined by visually inspecting each log for feeding damage on the bark and/or sapwood that led to an EAB larval gallery or pupal chamber in 2014 and 2016. EAB density was measured as the sum of the number of D-shaped exit holes and woodpecker predation holes on each ash log. The predation rate was then calculated as feeding damage divided by the sum of feeding damage plus number of D-shaped exit holes.
Data analysis
We used a generalized linear mixed model (GLMM) and assumed a binomial distribution and a logit link function to explore the effects of treatment and year on EAB parasitism. We also used a GLMM and assumed a binomial distribution with a logit link function to explore the effects of treatment, year, and EAB density on woodpecker predation. The relationship between EAB density and treatment or year were analyzed using a linear mixed-effects model (LMM). Treatment, year, and EAB density were used as fixed effects in the GLMMs and LMM, and random effects were used as a block effect in each model to account for release and control plots being nested within each of our three forested sites. Tukey’s range test was used to explore any differences between means of significant effects.
R was used to analyze all data (R Development Core Team 2018) and interaction terms were considered for all models. If there were no significant interactions, then they were removed and the simplest model was used to consider main effects. Statistical significance was set at P < 0.05 and all errors reported were set at ± SE.
Results
Data from southern Ontario sites collected by the Canadian Forest Service in the year before our first release (2012) found EAB parasitism to range from 0.6% EAB for Atanycolus spp. to 49.4% for P. sulcata (n = 2063 EAB). In the first year of our releases (2013), we subsampled a small number of ash logs being transported to the parasitoid-release plots in Toronto (n = 5 logs) and found EAB parasitism to be 46.15% for P. sulcata, 30.77% for Atanycolus spp., and 7.69% for Balcha indica (Mani and Kaul) (Hym: Eupelmidae). Of the 13 EAB larvae collected in these logs, 11 were parasitized by native parasitoids suggesting that only a small fraction of EAB were transported into our parasitoid-release sites.
Despite the relatively low release numbers, Phasgonophora sulcata and Metapelma spectabile Westwood (Hym: Eupelmidae) were found emerging from ash trees after only one year in our release plots where no previous parasitism had been recorded. Spathius floridanus Ashmead (Hym: Braconidae) was also found emerging from trees in our release plots. However, percentage parasitism could not be determined in this case as more than one wasp usually emerges from each EAB host (i.e., gregarious). Atanycolus spp. were found emerging from ash trees in both our control and release plots. Balcha indica emerged from the sample logs being transported to the release plots. However, it was not recovered in either the release or control plots over any of the sampling years (Table 1). All of these species are parasitoids attacking other native woodborers in North America.
Overall EAB parasitism by native parasitoids was influenced significantly by treatment (χ2 = 5.61; df = 1; P = 0.018) and year (χ2 = 52.59; df = 2; P < 0.001) with parasitism significantly higher in the release plots than in the control plots. Parasitism levels were significantly higher in 2016 than in earlier years (2013 or 2014) with on average 64.8 ± 18.1% of EAB parasitized in all three release plots.
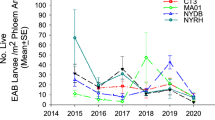
EAB density ranged from 0 to 7 EAB per m2 of ash surface area across all release and control plots with highest levels (i.e., 7 EAB per m2 of ash surface area) at one release plot in 2014. There were no differences in EAB density between treatment plots (χ2 = 0.45; df = 1; P = 0.500) or treatment years (χ2 = 1.69; df = 2; P = 0.431) (Table 2).
Woodpecker predation also varied across the release and control plots, with a low of 0% to a high of 70.3% EAB predation observed on the ash logs collected, with highest levels in control plots in 2016 (49.32 ± 20.98%) (Table 1). Woodpecker predation did not differ significantly between the release and control plots (χ2 = 1.70; df = 1; P = 0.192) nor between years (χ2 = 1.08; df = 1; P = 0.299). However, woodpecker predation did increase with EAB density (χ2 = 6.89; df = 1; P = 0.009).
Discussion
We show that EAB parasitism by North American parasitoids can be significantly increased by augmenting their populations through transport of parasitoid-infested ash logs. Studies on native parasitoids attacking EAB usually report low parasitism (i.e., < 1%) in newly-invaded regions (Liu et al. 2003; Duan et al. 2010, 2012). However, high parasitism by P. sulcata and Atanycolus spp., notably A. cappaerti, have been observed in some American and Canadian sites including ours (Cappaert and McCullough 2009; Lyons 2010). Evidence suggests that these parasitoids may not have strong dispersal capacities, especially P. sulcata (Gaudon et al. 2018), and that they are influenced by vegetation and habitat characteristics found within the sites, notably tree biomass, tree condition, canopy closure, and floral resource availability (Gaudon 2019). Thus, their populations would not be expected to move quickly following the spread of EAB and they are in need of augmentation in target areas to combat the large variation in population size and impact on EAB across the landscape.
Native natural enemies, such as P. sulcata in Ontario, Canada and A. cappaerti in Michigan, USA, are considered important due to high rates of observed parasitism on EAB (e.g., Cappaert and McCullough 2009; Lyons 2010). Other native species of Atanycolus that have been reared from EAB in Ontario include A. hicoriae Shenefelt, A. tranquebaricae Shenefelt, A. disputabilis Cresson, and A. longicauda Shenefelt (Roscoe 2014). Unfortunately, other wasp species, such as M. spectabile, have a relatively rare association with EAB (Lyons 2010) and thus have not been considered for biological control. Interestingly, B. indica has been observed attacking EAB in North America, although it is a non-native parasitoid established in North America prior to the arrival of EAB (Gibson 2005; Duan et al. 2012). Although not yet reported attacking EAB in Ontario, Canada, S. floridanus has also been shown to have potential for biological control of EAB in North America as it can be reared from EAB larval galleries in ash trees from Michigan, USA (Marsh and Strazanac 2009). Here, we report the first observation of S. floridanus attacking EAB in Canada. Until now, S. floridanus has only been recorded from native hosts in eastern North America (Lyons 2010) and has only been found on EAB in the USA (Duan et al. 2012). We suspect this is an artefact of our biocontrol efforts as it was undetected during our baseline sampling for native EAB parasitoids in the parasitoid release and non-release control plots, and remained absent in the control plots over the entire study period.
The EAB parasitism rates achieved after augmentative releases here were similar to those parasitism rates seen in existing classical biological control programs with non-native T. planipennisi and O. agrili. Duan et al. (2018) found that EAB parasitism by T. planipennisi was ~ 1 to 6% between 2008 and 2011, increasing to ~ 30% parasitism by 2014 in both parasitoid-release and control plots, while O. agrili averaged 1 to 4% EAB parasitism between 2008 and 2011, increasing to ~ 28% by 2014 in release plots. We found a comparable steady increase in overall EAB parasitism by Atanycolus spp. in parasitoid-release plots. However, we observed decreases in EAB parasitism by both M. spectabile and P. sulcata in the release plots by the end of the three-year study. For M. spectabile, it is possible that too few individuals were transported, and mate limitation negatively affected their population growth and establishment.
In the case of P. sulcata, the decrease in parasitism might be due to an increase in host refuge for EAB as parasitism would become increasingly limited by the thicker bark found at the lower levels in the tree [i.e., ~ 12.0 mm in trunks and large structural branches (J.M. Gaudon, unpublished data)]. EAB infestations usually progress from the upper canopy and branches downwards as populations build up in a given tree, and the relatively short ovipositor of P. sulcata [i.e., ~ 6.4 mm (Roscoe 2014)] means that these wasps would be unable to parasitize many EAB larvae within the plots, especially those at the lower tree levels. Abell et al. (2012) observed similar refuges for EAB from T. planipennisi, where this introduced parasitoid was unable to successfully parasitize EAB in trees having > 3.2 mm bark thickness. It is possible then that P. sulcata will be most effective for biocontrol on ash saplings in young stands and become an important mortality factor in regenerating ‘aftermath’ forests, similar to T. planipennisi (Duan et al. 2017). In contrast to T. planipennisi and P. sulcata, Atanycolus spp. have much longer ovipositors and can successfully parasitize EAB in ash trees with bark thickness up to 8.8 mm (Abell et al. 2012). In our study, we saw similar EAB mortality by Atanycolus spp. as Duan et al. (2019) with introduced Spathius spp.
Although EAB parasitism was significantly higher in the parasitoid-release plots than the control plots, there was no difference in EAB density between the treatment plots or between years. Duan et al. (2015) also found no difference in EAB densities between parasitoid-release and control plots over a seven-year study with T. planipennisi, although they did see significant differences in EAB density by year. In their study, increased parasitism by native parasitoids decreased EAB densities and this reduction was furthered with introductions of non-native T. planipennisi suggesting that both are important mortality factors on EAB (Duan et al. 2015). It is possible that the shorter duration of our study (three years vs. their seven years) obscured any immediate effect on EAB density. However, it is likely that a long-term increase may eventually appear since we found native parasitism differed between the release and control plots and others have correlated parasitism by both native and introduced EAB parasitoids with declines in EAB density (Duan et al. 2015). Thus, it would be beneficial to continue to establish populations of both native and introduced parasitoids in biological control programs against EAB as it spreads across North America in order to augment and accelerate population mortality.
We found no difference in woodpecker predation between parasitoid release and non-release plots and treatment years. However, not surprisingly, woodpecker predation did increase with EAB density. Past research examining the response of woodpeckers to EAB has been mixed. Lindell et al. (2008) and Jennings et al. (2013) observed linear increases in predation with increasing larval density while Duan et al. (2010) reported no relationship. It is known that woodpeckers are less likely to forage on parasitized than unparasitized EAB larvae, although this does not appear to impact overall EAB parasitism (Murphy et al. 2018). Murphy et al. (2018) hypothesized that a reduction in food quality from parasitism would make it less rewarding for woodpeckers to forage on parasitized EAB, and thus make them more likely to leave low-quality food patches of parasitized larvae to locate one of higher quality (and fewer parasitized EAB), thereby dampening the effect. Another factor that may explain the poor relationship between woodpecker predation and EAB density is that not all EAB larvae are equal, i.e., some EAB may be more active than others and this would allow better detection by woodpeckers. For example, idiobiont parasitoids such as Atanycolus spp. attack and paralyze their host prior to oviposition, making their host larvae less likely to be discovered by woodpeckers. Such a mechanism would change predation rates and may be particularly important in our system given that we found Atanycolus spp. to be the primary driver of increasing EAB parasitism and woodpecker predation decreased, albeit not significantly, as EAB parasitism by Atanycolus spp. increased in our release plots. Overall, Duan et al. (2018) predicted that EAB populations would decrease (growth rate r < 1) when ~ 95% EAB mortality occurred (comprised of approximately 60% woodpecker predation and 35% parasitism). Our research also suggests that such high mortality could be achieved through the action of native natural enemies (parasitoids and woodpeckers) alone.
It appears that the transport of recently cut ash logs is an effective means of augmenting EAB parasitoid populations, and this may be an easier and more efficient approach compared to mass rearing in the laboratory or a production facility. Research on another invasive forest insect, Sirex noctilio F. (Hym: Siricidae), has shown that redistributing infested plant material is both effective in disseminating natural enemies and cost-effective compared to rearing cultures. They recommended this approach for long-term management of Sirex in southern Australia (Haugen and Underdown 1990). Their approach allowed populations of Megarhyssa nortoni (Cresson) (Hym: Ichneumonidae), an important parasitoid of S. noctilio (Haugen and Underdown 1990), to be increased for release in numbers larger than previous rearing efforts in the laboratory. Although we observed no difference in EAB densities between treatment plots, the transport of parasitoid-infested ash logs can still be recommended as part of a broader strategy to slow the spread of EAB since it will result in extremely abundant populations of natural enemies and increase parasitism under field conditions. In our study, we moved infested logs over short distances (i.e., < 250 km) and only within the EAB quarantine/infested range. While there is potential to introduce a limited number of EAB to new areas, if used with caution, this approach will increase EAB mortality and this, combined with natural mortality from woodpecker predation and other natural enemies, has potential to limit its spread.
References
Abell KJ, Duan JJ, Bauer L, Lelito JP, van Driesche RG (2012) The effect of bark thickness on host partitioning between Tetrastichus planipennisi (Hymen: Eulophidae) and Atanycolus spp. (Hymen: Braconidae), two parasitoids of emerald ash borer (Coleop: Buprestidae). Biol Control 63:320–325
Anulewicz AC, McCullough DG, Cappaert DL (2007) Emerald ash borer (Agrilus planipennis) density and canopy dieback in three North American ash species. Arbor Urban Forestry 33:338–349
Bauer LS, Liu H, Haack RA, Petrice TR, Miller DL (2004) Natural enemies of emerald ash borer in southeastern Michigan. In: Mastro V, Reardon R (eds) Emerald ash borer research and technology development meeting, 30 September-1 October 2004, Port Huron, Michigan, United States of America. United States Department of Agriculture, Morgantown, West Virginia, United States of America, pp 33–34
Cappaert D, McCullough DG (2009) Occurrence and seasonal abundance of Atanycolus cappaerti (Hymenoptera: Braconidae) a native parasitoid of emerald ash borer, Agrilus planipennisi (Coleoptera: Buprestidae). Great Lakes Entomol 51:152–165
Cappaert D, McCullough DG, Poland TM, Siegert NW (2005) Emerald ash borer in North America: a research and regulatory challenge. Am Entomol 51:152–165
Crosthwaite JC, Sobek S, Lyons DB, Bernards MA, Sinclair BJ (2011) The overwintering physiology of the emerald ash borer, Agrilus planipennis Fairmaire (Coleoptera: Buprestidae). J Insect Physiol 57:166–173
Duan JJ, Fuester RW, Wildonger J, Taylor PB, Barth S, Spichiger SE (2009) Parasitoids attacking the emerald ash borer (Coleoptera: Buprestidae) in western Pennsylvania. Fla Entomol 92:588–592
Duan JJ, Ulyshen MD, Bauer LS, Gould J, van Driesche R (2010) Measuring the impact of biotic factors on populations of immature emerald ash borers (Coleoptera: Buprestidae). Environ Entomol 39:1513–1522
Duan JJ, Bauer LS, Abell KJ, van Driesche R (2012) Population responses of hymenopteran parasitoids to the emerald ash borer (Coleoptera: Buprestidae) in recently invaded areas in north central United States. BioControl 57:199–209
Duan JJ, Bauer LS, Abell KJ, Ulyshen MD, van Driesche RG (2015) Population dynamics of an invasive forest insect and associated natural enemies in the aftermath of invasion: implication for biological control. J Appl Ecol 52:1246–1254
Duan JJ, Bauer LS, van Driesche RG (2017) Emerald ash borer biocontrol in ash saplings: the potential for early stage recovery of North American ash trees. For Ecol Manag 394:64–72
Duan JJ, Bauer LS, van Driesche RG, Gould JR (2018) Progress and challenges of protecting North American ash trees from the emerald ash borer using biological control. Forests 9:142
Duan JJ, van Driesche RG, Crandall RS, Schmude JM, Rutledge CE, Slager BH, Gould JR, Elkinton JS (2019) Establishment and early impact of Spathius galinae (Hymenoptera: Braconidae) on Emerald Ash Borer (Coleoptera: Buprestidae) in the Northeastern United States. J Econ Entomol 112:2121–2130
Edworthy AB, Drever MC, Martin K (2011) Woodpeckers increase in abundance but maintain fecundity in response to an outbreak of mountain pine bark beetles. For Ecol Manag 261:203–210
Gaudon JM (2019) Natural enemies of wood-boring beetles in northeastern temperate forests and implications for biological control of the emerald ash borer (Coleoptera: Buprestidae) in North America. PhD thesis, University of Toronto, Canada.
Gaudon JM, Allison JD, Smith SM (2018) Factors affecting the dispersal of a native parasitoid, Phasgonophora sulcata, attacking the emerald ash borer: implications for biological control. BioControl 63:751–761
Gibson GAP (2005) The world species of Balcha Walker (Hymenoptera: Chalcidoidea: Eupelmidae), parasitoids of wood-boring beetles. Zootaxa 1033:1–62
Haack RA, Jendek E, Liu H, Marchant KR, Petrice TR, Poland TM, Ye H (2002) The emerald ash borer: a new exotic pest in North America. Newsl Michigan Entomol Soc 47:1–5
Haugen DA, Underdown MG (1990) Release of parasitoids for Sirex noctilio by transporting infested logs. Aust Forestry 53:266–270
Herms DA, McCullough DG (2014) Emerald ash borer invasion of North America: history, biology, ecology, impacts, and management. Annu Rev Entomol 59:13–30
Jennings DE, Gould JR, Vandenberg JD, Duan JJ, Shrewsbury PM (2013) Quantifying the impact of woodpecker predation on population dyanmics of the emerald ash borer (Agrilus planipennis). PLoS ONE 8(12):e83491
Jennings DE, Duan JJ, Bauer LS, Schmude JM, Wetherington MT, Shrewsbury PM (2015) Temporal dynamics of woodpecker predation on emerald ash borer (Agrilus planipennis) in northeastern USA. Agric For Entomol 18:174–181
Jiao Z-B, Wan T, Wen J-B, Hu J-F, Luo Y-Q, Zhang L-S, Fu L-J (2008) Functional response and numerical response of great spotted woodpecker Picoides major on Asian longhorned beetle Anoplophora glabripennis larvae. Acta Zool Sin 54:1106–1111
Knight KS, Brown JP, Long RP (2013) Factors affecting the survival of ash (Fraxinus spp.) trees infested by emerald ash borer (Agrilus planipennis). Biol Invasions 15:371–383
Kyei-Poku G, Johny S (2013) Evaluation of indigenous Beauveria isolates as potential agents for emerald ash borer management and the development of a diagnostic marker to monitor a post-release isolate. Int Org Biol Int Cont-West Palaear Reg Sec Bull 90:119–124
Lindell CA, McCullough DG, Cappaert D, Apostolou NM, Roth MB (2008) Factors influencing woodpecker predation on emerald ash borer. Am Midl Nat 159:434–444
Liu H, Bauer LS, Gao R, Zhao T, Petrice TR, Haack RE (2003) Exploratory survey for the emerald ash borer, Agrilus planipennis (Coleoptera: Buprestidae), and its natural enemies in China. Great Lakes Entomol 36:191–204
Lyons DB (2010) Biological control of emerald ash borer. In: Lyons DB, Scarr TA (eds) Guiding principles for managing the emerald ash borer in urban environments. Natural Resources Canada and Ontario Ministry of Natural Resources, Burlington, pp 29–34
MacQuarrie CJK, Williams DJ, Langor DW (2013) Update on the establishment of birch leafminer parasitoids in western Canada. J Entomol Soc BC 110:35–37
MacQuarrie CJK, Lyons DB, Seehausen ML, Smith SM (2016) A history of biological control in Canadian forests, 1882–2014. Can Entomol 148(S1):S239–S269
Marsh PM, Strazanac JS (2009) A taxonomic review of the genus Spathius Nees (Hymenoptera: Braconidae) in North America and comments on the biological control of the emerald ash borer (Coleoptera: Buprestidae). J Hymenopt Res 18:80–112
Murphy TC, Gould JR, van Driesche RG, Elkinton JS (2018) Interactions between woodpecker attack and parasitism by introduced parasitoids of the emerald ash borer. Biol Control 122:109–117
R Development Core Team (2018) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. URL https://www.r-project.org
Roscoe LE (2014) Phasgonophora sulcata Westwood (Hymenoptera: Chalcididae): a potential augmentative biological control agent for the invasive Agrilus planipennis (Fairmaire) (Coleoptera: Buprestidae) in Canada. PhD thesis, University of Toronto, Canada.
Roscoe LE, Lyons DB, Smith SM (2016) Observations on the life-history traits of the North American parasitoid Phasgonophora sulcata Westwood (Hymenoptera: Chalcididae) attacking Agrilus planipennis (Coleoptera: Buprestidae) in Ontario, Canada. Can Entomol 148:294–306
Rutledge CE, Fierke MK, Careless PD, Worthley T (2013) First detection of Agrilus planipennis in Connecticut made by monitoring Cerceris fumipennis (Crabonidae) colonies. J Hymenopt Res 32:75–81
Siegert NW, McCullough DG, Liebhold AM, Telewski FW (2014) Dendrochronological reconstructions of the epicentre and early spread of emerald ash borer in North America. Divers Distrib 20:847–858
Smith SM (1996) Biological control with Trichogramma: advances, successes, and potential of their use. Annu Rev Entomol 41:375–406
Sobek-Swant S, Crosthwaite JC, Lyons DB, Sinclair BJ (2012) Could phenotypic plasticity limit an invasive species? Incomplete reversibility of mid-winter deacclimation in emerald ash borer. Biol Invasions 14:115–125
Tannis SR, McCullough DG (2012) Differential persistence of blue ash and white ash following emerald ash borer invasion. Can J Forest Res 42:1541–1550
Acknowledgements
We thank J. Trott, D. Little, J. DeZwart, and J. Lilley for access to woodlots; L. Roscoe, C. de Keyzer, G. Jones, and D. B. Lyons for help with parasitoid collection; K. Vitols and J. Ric for site selection and sampling; and M. Orr for rearing space. Funding was provided by an Ontario Graduate Scholarship, Queen Elizabeth II Graduate Scholarship in Science and Technology, University of Toronto Fellowship, the Ontario Ministry of Natural Resources and Forestry, and the Invasive Species Centre.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Handling Editor: Dirk Babendreier
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Gaudon, J.M., Smith, S.M. Augmentation of native North American natural enemies for the biological control of the introduced emerald ash borer in central Canada. BioControl 65, 71–79 (2020). https://doi.org/10.1007/s10526-019-09986-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-019-09986-6