
Abstract
Hymenopterous parasitoids of herbivorous insects can be useful biocontrol agents in integrated pest management strategies. However, the potential effects on these beneficials of new components in such strategies are often neglected. Essential oil of lavender, Lavendula angustifolia (Miller) (Lamiaceae), has recently been identified as a potential repellent in new control strategies being developed for the pollen beetle Meligethes aeneus (Fabricius) (Coleoptera: Nitidulidae), a major pest of oilseed rape, Brassica napus L. (Brassicaceae). We tested the electrophysiological and behavioural responses of two common parasitoids of M. aeneus: Phradis interstitialis (Thomson) and P. morionellus (Holmgren) (both Ichneumonidae) using coupled gas chromatography-electroantennodetection (GC-EAD) and olfactometry techniques. Both species elicited electrophysiological responses to lavender oil volatiles, including two compounds known to be repellent to M. aeneus. However, the parasitoids gave no significant responses to the odours of lavender oil in behavioural bioassays and there was no evidence to suggest that lavender-treated oilseed rape plants would reduce host habitat location by parasitoids of the target pest.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The use of botanical extracts as insecticides, deterrents and repellents in insect pest management is receiving increasing attention (reviewed by Isman 2006; Cook et al. 2007). Essential oils from plant species of the Laminaceae have a broad spectrum of biological activities and essential oil of lavender Lavendula angustifolia (Miller) (Lamiaceae) has been reported as interfering with orientation to, and selection of, host plants by a range of pest insects (Landolt et al. 1999; Koschier et al. 2002; Koschier and Sedy 2003; Mauchline et al. 2005). It has therefore been proposed as a useful component of alternative pest management strategies, but effects on the natural enemies of the pests concerned have never been investigated.
Both insect herbivores and their hymenopterous parasitoids often use olfactory cues from plants during host location. Plant volatiles provide reliable cues enabling herbivores to distinguish host from non-host plants (Bernays and Chapman 1994). Parasitoids also often depend on plant cues, because those from the insect host can be difficult to detect (Vet et al. 1995). These behaviours potentially confound the benefits to pest management strategies of using repellents and non-host volatiles that interfere with pest location and selection of crops, since their natural enemies may be similarly affected, thereby reducing biocontrol efficacy.
The pollen beetle, Meligethes aeneus (Fabricius) (Coleoptera: Nitidulidae) is a Brassica specialist and a major pest of oilseed rape (Brassica napus L.) (Brassicaceae). Insecticide-resistant populations are reported across mainland Eurpoe (Ekbom and Kuusk 2001; Hansen 2003; Wegorek 2006), and new control strategies are sought. Meligethes aeneus is known to be attracted to odours from its host plants (Evans and Allen-Williams 1994; Ruther and Thiemann 1997; Cook et al. 2002; Mauchline et al. 2005; Cook et al. 2006). The use of non-host volatiles to mask this attraction or to repel M. aeneus from the crop has been investigated and essential oil of lavender was found to be an effective repellent (Mauchline et al. 2005).
Parasitoids can act as very efficient biocontrol agents of M. aeneus, with parasitism rates often exceeding 50% (Nilsson 2003). Phradis interstitialis (Thomson) and P. morionellus (Holmgren) (Ichneumonidae) are the most abundant specialist parasitoids of M. aeneus across Europe (Nilsson 2003). They are univoltine and oviposit in the first and second larval instars of their hosts, respectively, during the bud and flowering growth stages of the host plant (Nilsson 2003). Both species are attracted to oilseed rape odours (Jönsson et al. 2005). Therefore, the use of non-host volatiles to disrupt colonisation of oilseed rape by M. aeneus may potentially also adversely affect host habitat location by their parasitoids.
We investigated the electrophysiological and behavioural responses of P. interstitialis and P. morionellus to lavender oil in order to determine the potential effect of the inclusion of this repellent in pest management strategies, prior to more detailed field studies.
Materials and methods
Insects and plant material
Parasitoids were collected from oilseed rape crops using a sweep net. They were stored at 10°C with a 16L:8D photoperiod in ventilated plastic boxes, with water and honey provided. They were used on the day of collection in electrophysiology studies and within four days in behavioural experiments. Only females were used; provisional species-level identifications of live parasitoids were made using ovipositor morphology and after experiments had been conducted, identification was confirmed by taxonomic examination (Horstmann 1971). Oilseed rape (cv. Aries) plants were grown individually in 3 l pots in a glasshouse. Lavender essential oil solutions were made using the same L. angustifolia variety (cv. Mailette, Botanix Ltd., Paddock Wood, Kent, UK), and at the same concentrations (1% and 10% v:v) as used in previous studies (Mauchline 2003; Mauchline et al. 2005), with focus on the 1% concentration, which was selected for further work and is being developed as a repellent for M. aeneus (Mauchline 2003; Mauchline et al. unpublished data).
Gas chromatography-electroantennodetection (GC-EAD)
In the GC-EAD recordings the effluent from the GC column was split (1:1) between the antennal preparation and the FID-detector of the GC. The posterior of the parasitoid head was placed in the ground-glass electrode containing Beadle-Ephrussi ringer solution (Bjostad 1998). The tips of the antennae were introduced into a recording electrode connected to an amplifier (Syntech, The Netherlands) and monitored on a PC. A SolGel-Wax column (SGE, 30 m × 0.25 mm i.d., 0.25 μm stationary phase) was installed in the GC (HP-6890). The oven temperature was maintained at 40°C for 1 min, then increased by 5°C/min to 200°C. The carrier gas was hydrogen and an injection volume of 1 μl lavender oil (0.1 % v:v hexane) was used. Six GC-EAD experiments with each species were conducted. Compounds eliciting responses in three or more of these were regarded as active. These were identified by GC-mass spectrometry using an HP-5890 GC connected to an HP-5970 mass detector (column and temperature conditions as described above). The carrier gas was helium and ionisation potential 70 eV. Obtained mass spectra were compared with the Wiley/NIST mass spectral database (1990) and with spectra obtained from standards. Identities were confirmed by comparison of retention times with standards on the described systems and also on an HP-5890 with an apolar column (DB-5; J&W Scientifics, 30 m × 0.32 mm I.d. 0.25 μm stationary phase). Stereochemistry of chiral compounds was not determined.
Behavioural responses
Experiments were conducted using a glass Y-tube olfactometer (Soham Scientific, Fordham, Cambs., UK; internal diameter 1.1 cm, 6.5 cm arm length, 8 cm stem length). Filtered air, pushed from a diaphragm pump, was divided into two airflows, each passing via a flow meter into a glass vessel (5 l) containing the test odour sources and then into the arms of the olfactometer (0.5 l/min). All visual stimuli were excluded behind screens surrounding the apparatus. Parasitoids were introduced individually into the stem of the Y-tube. Those that crossed a line 3.5 cm from the end of an arm and remained there for more than 10 s were considered to have made a choice. Parasitoids making no choice after five minutes were excluded from the analysis. Experiments were conducted at 20°C ± 2°C in a controlled environment room. The apparatus was lit by an overhead fluorescent light that emited daylight wavelengths and was fitted with a plane-polarizing prismatic refractive cover to provide even illumination. The olfactometer was turned through 180° between each run and the Y-tube and glass vessels were washed and heated to 200°C after every 5 runs.
The behavioural responses of P. interstitalis and P. morionellus to olfactory stimuli were tested in three choice tests: (1) oilseed rape vs. moist filter paper control (2) 10 μl 1% lavender oil vs. 10 μl acetone (3) oilseed rape with 10 μl 1% lavender oil vs. oilseed rape with 10 μl acetone. Phradis morionellus, which emerges later than P. interstitalis and is available for a longer period, was tested in two further experiments: (4) 10 μl 10% lavender oil vs. 10 μl acetone, and (5) oilseed rape sprayed with a formulation of lavender oil in gum Arabic suitable for field application vs. oilseed rape sprayed with gum Arabic formulation (spray rate was 2.5 kg ha−1 at 200 l ha−1 vol.). For experiments 2–4, acetone was used as the solvent for the lavender oil. The lavender or acetone treatments were applied to filter paper (1.8 cm diameter) and introduced into the glass vessels; the disc was replaced every 5 runs. For P. interstitialis, oilseed rape odour was provided by 10 g racemes with some flowers (BBCH growth stage 63: Lancashire et al. 1991). For P. morionellus, 20 g racemes in green bud were used (BBCH 55–57), because in preliminary tests, they did not respond to flowering racemes. The racemes were placed in 10 ml water; corresponding controls were 10 ml water. With both parasitoid species, replicates of each experiment were performed each day. Oilseed rape racemes were changed with each experiment (within four hours of excision from the plant) and parasitoids were used only once. Between 69–101 individual parasitoids were used for each test. A log-linear analysis was used to assess whether there was a significant difference in preference between the treatments in each experiment. Terms were fitted for day, orientation of the equipment and choice of odour. The interaction terms in the model were used to test the effect of day and orientation of the equipment on the choice of odour (GenStat, 9th Edition, VSN International).
Results and discussion
Both parasitoid species detected lavender essential oil volatiles in concentrations known to be electrophysiologically active for their host, M. aeneus (Mauchline 2003) (Fig. 1). Phradis interstitialis responded to seven compounds, and five elicited responses in P. morionellus (Fig. 1). All of the active compounds are common floral volatiles and none are specific to lavender. In a previous study, two of the active compounds, linalool and linalyl acetate, also elicited electrophysiological responses in M. aeneus and were repellent to the beetle in behavioural tests (Mauchline 2003). If behavioural responses of parasitoids had been observed to the lavender essential oil, then further bioassays would have been performed to determine the role of the active components in the observed attraction or repulsion. However, no such responses were observed (Table 1). Furthermore, the potential use of lavender as a repellent involves deployment of the essential oil itself, and not deployment of individual compounds.

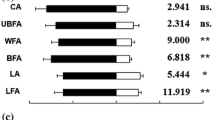
Electrophysiological responses of Phradis interstitialis parasitoids to volatiles of lavender essential oil. The upper trace represents chemical detector (FID) responses and the lower trace represents antennal responses (EAD). Numbered peaks elicited responses in at least 3 of 6 coupled runs and correspond to the following volatiles; numbers in square brackets following each compound represent the number of responses shown by P. interstitialis and P. morionellus, respectively: (1) 3-octanone [5,1]; (2) octen-1-ol, acetate [3,5]; (3) linalool [6,5]; (4) linalyl acetate [4,3]; (5) β-caryophyllene [2,4]; (6) (E)-β-farnesene [4,1]; (7) borneol [4,0]; (8) geranyl acetate [4,3]. Note: *EAD response to this compound not shown on this example trace
In olfactometer tests, no evidence was found in any of the experiments of an effect of day or orientation of the equipment on the parasitoids choice of odour (P >0.05 in all cases). Both species of parasitoid chose the arm into which oilseed rape odours were introduced significantly more often than the control arm (Table 1). This suggests these parasitoids are attracted to odours from the host plant of their host and importantly, shows that the experimental set up is working, in that a positive response was shown to a positive control; this is necessary when evaluating potential repellent responses. In a previous study, Jönsson et al. (2005) showed that P. morionellus was attracted to the odour of buds as we found, but that P. interstitialis was repelled by the odour of flowering racemes alone; a yellow coloured stimulus was necessary to evoke a preference in this species. The reason for the apparent discrepancy between current and previous observations needs further investigation, but could be explained by differing growth stages of the flowering racemes used (several buds were also present in our samples), and differing experimental conditions (experiments were performed under controlled environmental conditions in the present study, and in outside conditions in the Jönsson et al. (2005) study).
Neither parasitoid species gave any significant responses to the odours of 1% lavender oil when this was presented against the control (Table 1). Phradis morionellus exhibited a similar lack of response to lavender odours when presented at a higher concentration (10%) (Table 1), also found previously to repel M. aeneus (Mauchline et al. 2005). Neither parasitoid species significantly discriminated against odours of oilseed rape presented with lavender when tested against oilseed rape odours alone (Table 1). Furthermore, P. morionellus did not respond differently to the odours of oilseed rape sprayed with lavender oil formulation compared with control plants (Table 1).
Our results indicate that although parasitoids can detect the lavender essential oil at the concentrations intended for use as a repellent against their host M. aeneus, they do not respond to them. Therefore, treating oilseed rape with lavender essential oil would not have a significant negative impact on host habitat location by these parasitoids. This is important, as biological control plays a key role in pest management strategies in which pesticide use is minimised.
The lavender formulation used in the behavioural study is now planned for use in field trials to test the effectiveness of lavender in reducing M. aeneus damage to oilseed rape crops. During these trials we aim to monitor the effects of the lavender odours on attraction of other parasitoids of M. aeneus and determine its effect on realized parasitism, which is also influenced by behaviours such as mate finding and host location. Olfactometer tests are relatively cheap and quick to perform and if validated by field trails, could be useful in screening potential repellents for possible negative effects on beneficials.
References
Bernays EA, Chapman RF (1994) Host-plant selection by phytophagous insects. Chapman & Hall, New York
Bjostad LB (1998) Electrophysiological methods. In: Millar JG, Haynes KF (eds), Methods in chemical ecology. Kluwer Academic Publishing, Dordrecht, The Netherlands. pp 339–375
Cook SM, Bartlet E, Murray DA, Williams IH (2002) The role of pollen odour in the attraction of pollen beetles to oilseed rape flowers. Entomol Exp Appl 104:43–50
Cook SM, Khan ZR, Pickett JA (2007) The use of ‘push-pull’ strategies in integrated pest management. Annu Rev Entomol 52:357–400
Cook SM, Smart LE, Martin JL, Murray DA, Watts NP, Williams IH (2006) Exploitation of host plant preferences in pest management strategies for oilseed rape (Brassica napus). Entomol Exp Appl 119:221–229
Ekbom B, Kuusk AK, (2001) Pollen beetles (Meligethes aeneus) and resistance against pyrethroids. Vaxtskyddsnotiser 65:39–42
Evans KA, Allen-Williams LJ (1994) Laboratory and field response of the pollen beetle, Meligethes aeneus, to the odour of oilseed rape. Physiol Entomol 19:285–290
Hansen LM (2003) Insecticide-resistant pollen beetles (Meligethes aeneus F) found in Danish oilseed rape (Brassica napus L) fields. Pest Manage Sci 59:1057–1059
Horstmann K (1971) Revision der europäichen Tersilochinen I (Hym., Ichneumonidae). Veröffentlichungen der Zoologischen Staatssammlung (München) 15:13–45
Isman MB (2006) Botanical insecticides, deterrents, and repellents in modern agriculture and an increasingly regulated world. Annu Rev Entomol 51:45–66
Jönsson M, Lindkvist A, Anderson P (2005) Behavioural responses in three ichneumonid pollen beetle parasitoids to volatiles emitted from different phenological stages of oilseed rape. Entomol Exp Appl 115:363–369
Koschier EH, Sedy KA (2003) Labiate essential oils affecting host selection and acceptance of Thrips tabaci lindeman. Crop Prot 22:929–934
Koschier EH, Sedy KA, Novak J (2002) Influence of plant volatiles on feeding damage caused by the onion thrips Thrips tabaci. Crop Prot 21:419–425
Lancashire PD, Bleiholder H, Vandenboom T, Langeluddeke P, Stauss R, Weber E, Witzenberger A (1991) A uniform decimal code for growth stages of crops and weeds. Ann Appl Biol 119:561–601
Landolt PJ, Hofstetter RW, Biddick LL (1999) Plant essential oils as arrestants and repellents for neonate larvae of the codling moth (Lepidoptera : Tortricidae). Environ Entomol 28:954–960
Mauchline AL (2003) Behavioural and chemical ecology of Meligethes aeneus: effects of non-host plant volatiles. Ph.D. Thesis, The Open University. pp 197
Mauchline AL, Osborne JL, Martin AP, Poppy GM, Powell W (2005) The effects of non-host plant essential oil volatiles on the behaviour of the pollen beetle Meligethes aeneus. Entomol Exp Appl 114:181–188
Nilsson C (2003) Parasitoids of pollen beetles. In: DV Alford (eds) Biocontrol of oilseed rape pests. Blackwell Publishing, Oxford, UK. pp 73–86
Ruther J, Thiemann K (1997) Response of the pollen beetle Meligethes aeneus to volatiles emitted by intact plants and conspecifics. Entomol Exp Appl 84:183–188
Vet LEM, Lewis WJ, Carde RT (1995) Parasitoids foraging and learning. In: RT Carde WJ Bell (eds) Chemical Ecology of Insects. Chapman & Hall, New York. pp 65–104
Wegorek P (2006) Resistance of pollen beetle (Meligethes aeneus F.) to pyrethroids, chloronicotinyls and organophosphorous insecticides in Poland. IOBC/wprs Bull 9(7):137–142
Acknowledgements
We thank Barry Pye for spraying the plants and Ingrid Williams for her support throughout this study. This work was facilitated by John Pickett and funded by the UK Department for Environment, Food and Rural Affairs. Rothamsted Research receives grant-aided support from the UK Biotechnology and Biological Sciences Research Council.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cook, S.M., Jönsson, M., Skellern, M.P. et al. Responses of Phradis parasitoids to volatiles of lavender, Lavendula angustifolia—a possible repellent for their host, Meligethes aeneus . BioControl 52, 591–598 (2007). https://doi.org/10.1007/s10526-006-9057-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-006-9057-x