
Abstract
The practice and development of the culture of commercially important penaeid shrimp results in the occurrence of infectious and non-infectious diseases worldwide. Most of the diseases are caused by the opportunistic microorganisms which are part of the microflora and fauna of the penaeid shrimp. Once the pathogens get a favourable condition, it will cause diseases to the host organisms. Most of the organisms are ubiquitous and are found and reported all over the cultured area in the world. The pathogens are the bacteria, viruses, parasites and protozoans. Viruses are the most important disease-causing agents in penaeid shrimps. The penaeid viruses are widely spread in geographical areas of shrimp culture. The shipment of broodstock and post larvae from one geographical region to another region often results in spreading of viral diseases. The penaeid viruses are the Whispovirus, white spot syndrome virus (WSSV), Penaeus monodon baculovirus (monodon baculovirus), infectious hypodermal and haematopoietic necrosis virus (IHHNV), infectious myonecrosis virus (IMNV), Taura syndrome virus (TSV), yellow head virus (YHV), white tail disease (WTD), covert mortality nodavirus (CMNV), Laem–Singh virus (LSNV) and shrimp haemocyte iridescent virus (SHIV). The review examines and highlights the aquaculture penaeid shrimp viral diseases in detail and concludes the need and importance of severe disease surveillance of the global countries through networking and coordination to safeguard the aquaculture practice for our future generation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Aquaculture is defined as the farming of aquatic organisms including fish, molluscs, crustaceans and aquatic plants under controlled conditions (FAO 1997). It involves the culture of animals and plants from aquatic environment including marine, brackish and freshwater environment. According to the findings and research, the widespread assumption that seafood production from capture fisheries is at or near its end. So, aquaculture will become an important source of seafood production and, ultimately, the main source of protein. Aquaculture plays a key role to overcome the protein shortage in the diet of peoples in poverty in developing countries (Subasinghe et al. 2009). World aquaculture production is increasing rapidly year by year compared to capture fisheries and animal husbandry (Ndanga et al. 2013).
The growth of aquaculture production of animals and plants is at a mean rate of 81% over year by year (FAO 2010). In 2004, aquaculture contributes 42% of total global seafood production. When compared to capture fisheries, aquaculture is not limited by the natural environment. Shellfish and finfish are the two major aquaculture families comprising more than 90% in aquaculture sector. Shellfish are aquatic invertebrates with shell. Bivalves (oyster, mussels, clams) and decapods (shrimp, crayfish, crabs) are the major aquaculture groups in shellfish family. The crustacean (marine shrimp) shows an increasing level of production by aquaculture throughout the global country day by day.
For the past three decades, farming of various marine shrimp species has developed widely. Between 1975 and 1985, the global production of farmed shrimp increased by 300%. There are totally 2500 species of shrimp worldwide, but nearly 12 species are farmed to some extent. All the twelve species are belonging to Penaeidae family. Out of the twelve species, two species account for more than 90–95% of global productions: one is black tiger shrimp (Penaeus monodon) and another one is Pacific white shrimp (Litopenaeus vannamei).
In the initial part of marine shrimp farming, the black tiger prawn (Penaeus monodon) was the preferred species. The global production of Penaeus monodon was 623,194 tonnes in 2000 (FAO) and the production of whiteleg shrimp (Litopenaeus vannamei) was only 2130 tonnes in that year. But due to the growth performance, low-level protein requirements and disease resistance, Litopenaeus vannamei overcomes Penaeus monodon in aquaculture production and farming.
The rapid growth of aquaculture in the twentieth and twenty-first centuries results in the occurrence of many emerging diseases. Diseases are the most important limiting factor in aquaculture. This includes bacterial, fungal, viral and parasitic diseases. Aquaculture has more biosecurity challenges now than ever before. The economic evaluations of diseases are relatively rare, but they have been determined and the costs are staggering. The best examples are the US$3 billion loss of cultured marine shrimp by diseases in 1994 (Lundin 1995) and the US$0.5 billion crash of cultured black tiger shrimp in Taiwan (1987–1988). Sustainable and severe management practices will be protecting our shrimp aquaculture field to safeguard the aquaculture industries. Then only, aquaculture will be safe and secure for our future need. This review paper detailed about the different types of viral diseases occurred in penaeid shrimp culture globally.
Yellow head virus
The first serious disease outbreak of yellow head virus (YHV) occurred in the adult Penaeus monodon culture stocks in Central Thailand in the 1990s and was first noted by Limsuwan. The disease was widespread in the Penaeus monodon stock and caused severe mortality in the adult culture of Penaeus monodon in Southeast Asia including China, India, Taiwan, Indonesia, Philippines and Vietnam (OIE 2009a). YHV was considered earlier as baculovirus, but after the purification and characterisation, it is morphologically differed from baculovirus. Presently, the YHV is classified in the genus Okavirus, family Roniviridae (Cowley et al. 2012). The disease caused by yellow head virus genotype 1 (YHV1) is one of the eight known genotypes in the yellow head complex of viruses. Yellow head genotype 2 is commonly called as gill-associated virus (GAV). Other four genotypes (genotypes 3–6) were identified in healthy P. monodon cultures in East Africa, Asia and Australia without any disease symptoms. Recently, YHV7 and YHV8 are the two new genotypes of yellow head virus that have been reported. YHV7 was isolated in diseased P. monodon in Australia (Mohr et al. 2015), and YHV8 was detected in Penaeus chinensis (Liu et al. 2014a). YHV1 has been reported in Chinese Taipei, Indonesia, Malaysia, Philippines, Sri Lanka, Thailand and Vietnam (Walker et al. 2001). GAV and other genotypes in the yellow head complex have been detected in healthy P. monodon from Australia, Chinese Taipei, India, Indonesia, Malaysia, Mozambique, Philippines, Thailand and Vietnam (Wijegoonawardane et al. 2008). YHV1 has also been detected in Penaeus vannamei in Mexico and Thailand (Sanchez-Barajas et al. 2009; Senapin et al. 2010).
YHV1 is a rod-shaped enveloped virion, approximately 40–50 nm × 150–180 nm in size, with prominent spine-like projection approximately 11 nm in size from the surface and with an inner helical nucleocapsid (Chantanachookin et al. 1993; Wongteerasupaya et al. 1995a). Nucleocapsid is 20–30 nm in size and appears as rods. The virion comprises a nucleoprotein (p20), envelope glycoproteins (gp64 and gp116) and a ~ 26-kb positive-sense single-stranded RNA genome. The RNA genome consists of approximately 26,662 nucleotides with a 3′-polyadenylated tail and contains four functional long open reading frames (ORFs) (ORF-1a, ORF-1b, ORF-2 and ORF-3) (Sittidilokratna et al. 2008).
YHV-infected shrimp normally dies within 3 to 5 days, and YHV can cause 100% mortality in 3 to 5 days. Apart from Penaeus monodon, the YHV1 affects Penaeus vannamei, Penaeus stylirostris, Palaemonetes pugio and Metapenaeus affinis. In P. monodon, yellow head virus infections occur at the adult stage of an animal or infect beyond post larval stage 15 (Khongpradit et al. 1995).
YHV1 affects the tissues of ectodermal and endodermal origin including lymphoid organ, haemocytes, haematopoietic tissue, gill lamellae and spongy connective tissue of the subcutis, gut, antennal gland, gonads, nerve tracts and ganglia. The clinical signs of YHV-infected shrimp are characterised by a pale-yellowish cephalothorax due to the discolouration of the hepatopancreas and gill (Chantanachookin et al. 1993; Lightner 1996a). The histological analysis of an YHV1-infected shrimp shows necrosis containing vacuolated cells with hypertrophied nuclei and basophilic viral inclusions in the cytoplasm of infected cells (Fig. 1). Cellular necrosis occurs in connective tissues, haematopoietic and lymphoid organs, abdominal muscle, haemocytes, nerves, hepatopancreas, midgut, heart, head muscle, lymphoid organs and gills.

a Histological section of the gills from a black tiger prawn (Penaeus monodon) with yellow head disease. A generalised, diffuse necrosis of cells in the gill lamellae is shown, and affected cells display pyknotic and karyorrhectic nuclei b, c Histological section of the lymphoid organ (LO) of a juvenile giant black tiger prawn (Penaeus monodon) with severe acute yellow head disease at low and high magnification d Gross signs of yellow-head infection are seen in the shrimps on the right. (Source: T.W. Flegel 2006)
Taura syndrome virus
Taura syndrome virus (TSV) was first reported in the L. vannamei shrimp collected from the shrimp farms located near the mouth of the Taura River in the Gulf of Guayaquil, Ecuador, in 1992, and it was believed that the disease was caused by the pesticide runoff from the nearby banana farms. After the epidemiological characterisation and laboratory analysis, they identified that the disease was caused by the virus (Jimenez 1992; Lightner et al. 1995a). TSV disease caused catastrophic losses to the L. vannamei shrimp farmers with the mortality rate ranging from 60 to 90%. After the first report of TSV and being recognised to cause a distinct disease to L. vannamei culture, it spread rapidly to the other shrimp culture regions of America, Peru and Mexico (Lightner 1996b, 1996c; Lightner and Redman 1998). TSV affects the penaeid shrimps P. vannamei, P. stylirostris, Penaeus schmitti, Penaeus setiferus and Penaeus aztecus in America and P. monodon, Penaeus japonicus and P. chinensis in Asia. In Asia, TSV was first reported in Taiwan in 1999 and the virus spread rapidly to China, Thailand, Malaysia and Indonesia (Cheng et al. 2011; Nielsen et al. 2005). Taura syndrome virus belongs to the genus Aparavirus (family Dicistroviridae) consisting of a non-enveloped, icosahedral virion with a diameter of 31–32 nm and having a buoyant density of 1.338 g ml−1 in CsCl. The genome of TSV consists of a linear, positive-sense single-stranded RNA genome having 10,205 nucleotides in length. The RNA genome has two ORFs: ORF-1 encodes the non-structural proteins including protease, helicase and a RNA-dependent RNA polymerase, and ORF-2 encodes three capsid proteins: VP-1 (40 kDa), VP-2 (55 kDa) and VP-3 (24 kDa). The functions of the capsid are still unknown, even though the genome of the TSV has been characterised and the virion has been identified (Bonami et al. 1997; Mari et al. 1998, 2002; Robles-Sikisaka et al. 2001).
TSV infections were reported in all three stages (post larvae (PL), juveniles and adults) of P. vannamei culture. Clinical signs of TSV-infected shrimp are characterised by lethargy, anorexia, opaque musculature and reddish discolouration in the tail fan and pleopods (Lightner et al. 1995a). The virus affects the hypodermis of the exoskeleton, foregut, hindgut, gills and appendages and often the connective tissues, haematopoietic tissues, lymphoid organ (LO) and antennal gland (Fig. 2). The histological analysis of the enteric organs and smooth, cardiac, striated muscle, and the ventral nerve cord, its branches and its ganglia has not shown any signs of infection by TSV.

Taura syndrome virus. a Tail fan showing reddening of the uropod and multifocal necrotic lesions (I, TSV-infected shrimp; H, normal shrimp). b Multifocal-melanised lesions on the cuticle (I, TSV-infected shrimp; H, normal shrimp). c Histology of H&E-stained ventral cephalothorax of TSV-infected shrimp showing the pyknotic nuclei, cytoplasmic eosinophilia and detachment from the surrounding cell matrix. d In situ hybridisation of TSV-infected cells displaying a probe-positive signal characterised by cytoplasmic blue-black colouration (arrow), while normal cells are golden orange. (Source: Dhar et al. 2010)
White spot syndrome virus
White spot disease is caused by white spot syndrome virus. The first occurrence of white spot syndrome has been reported in Taiwan in 1992 (Chen 1995). After that, it spread rapidly in the shrimp culture regions of Asia and Indo-Pacific. Although isolates from different geographical area with different genotypes have been reported, all are classified as a single species within the genus Whispovirus (family Nimaviridae). In earlier days, five baculoviruses have been reported to cause white spot disease in different geographical areas, such as hypodermal and haematopoietic necrosis baculovirus (HHNB) in China (Huang et al. 1994); systematic ectodermal and mesodermal baculovirus (SEMBV) in Thailand (Wongteerasupaya et al. 1995b) and Bangladesh (Ahmed 1996); rod-shaped nuclear virus in P. japonicus culture stock (RV-PJ) in Japan, China and South Korea (Inouye et al. 1994); white spot baculovirus (WSBV) in India, Malaysia, Vietnam and Indonesia (Wang et al. 1995); and non-occluded baculovirus in Penaeus monodon (PMNOB) in Taiwan (Lo et al. 1995), America (Rosenberry 1996, 2000) and France (Rosenberry 2002). Later, it has been reported that all viruses in the group are very similar in morphology and replicated in the nuclei of the infected cells of mesodermal and ectodermal origin. The characteristic of white spot syndrome virus is more similar to the family Baculoviridae. Volkman et al. (1995) reported that the white spot virus is not a baculovirus and referred to names as white spot syndrome virus (WSSV). WSSV can infect aquatic crustacean including marine, fresh and brackish water shrimps, prawns, crayfish and lobsters.
According to the International Committee on Taxonomy of Viruses (ICTV), WSSV is the only member in the genus Whispovirus (family Nimaviridae). The virions of WSSV are ovoid or ellipsoid to bacilliform in shape with a diameter of 80–120 nm and a length of 250–380 nm. In some isolates, a flagella-like extension has been observed at the one end of the virion (Lo et al. 2012). The nucleic acid of WSSV is composed of a double-stranded DNA (dsDNA) molecule with 100 to 200 kbp in length (Wongteerasupaya et al. 1995a).
WSSV infects and causes mass mortality in P. monodon, Penaeus semisulcatus, P. japonicus, P. chinensis, Penaeus penicillatus (redtail prawn), Penaeus indicus, Penaeus merguiensis, Trachypenaeus curvirostris (southern rough shrimp), Metapenaeus ensis (greasyback shrimp), Macrobrachium rosenbergii (giant river prawn), Procambarus clarkia (red swamp crayfish) and a variety of marine crabs and lobsters (Chang et al. 1998; Lightner 1996b; Wang et al. 1997). WSSV can be susceptible at all stages of the animal from eggs to broodstock (Lightner 1996b; Venegas et al. 1999). It infects the ectodermal and mesodermal origin tissues especially connective tissues and subcuticular connective tissues. Infection of endodermal tissues with WSSV is still not confirmed. The histological analysis showed the intranuclear viral inclusions with hypertrophied nuclei and marginated chromatin (Fig. 3). The similar type of inclusions was shown in the histopathological study of infectious hypodermal and haematopoietic necrosis virus (IHHNV) and other DNA virus–infected tissues.

White spot syndrome virus. a Carapace and columnar epithelial cells showing enlarged nuclei. b Stomach epithelium showing hypertrophied nuclei with eosinophilic staining. c Enlarged infected nuclei in the epithelial cells. d Gill cuticular epithelium with enlarged, infected nuclei (source: Bateman et al. 2012). (1) Clinical signs of white spot virus–infected animal. (2) White spots on the cephalothorax (Lightner 1996b)
Infectious hypodermal and haematopoietic necrosis virus
IHHNV was first reported in P. stylirostris and P. vannamei in the early 1980s in the Americas (Lightner et al. 1983a; Lightner 2011b) and later in P. monodon from Asia. After the initial identification, it spread rapidly along the culture regions in Mexico, Central America, Ecuador, Peru, Brazil, numerous Caribbean Islands, Hawaii, Guam, Tahiti, New Caledonia, Singapore, Philippines, Thailand, Malaysia, Indonesia (Lightner 1996b), India (Rai et al. 2009), Australia (Krabsetsve et al. 2004; Owens et al. 1992; Tang and Lightner 2006) and China (Zhang and Sun 1997). According to the aquatic animal health code, the susceptible host species for IHHNV infection are Penaeus californiensis (yellowleg shrimp), Penaeus monodon (giant tiger prawn), Penaeus setiferus (northern white shrimp), Penaeus stylirostris (blue shrimp) and Penaeus vannamei (white leg shrimp) (Table 1). IHHNV has been reported in all life stages of the animal (eggs to adult). The eggs produced by the IHHNV-infected females were found to be rich in virus load and fail to hatch out (Motte et al. 2003). Natural infections of IHHNV have been detected from Penaeus stylirostris, P. vannamei, Penaeus occidentalis, P. californiensis, P. monodon, P. semisulcatus and P. japonicus. P. setiferus, Penaeus duorarum and P. aztecus have been infected experimentally with IHHNV, and P. indicus and P. merguiensis were found to be refractory to infection (Brock and Lightner 1990a; Lightner 1996b). Some members of P. stylirostris and P. vannamei from different geographical areas are found to have survived with IHHNV infection and carry the virus to the progeny and other populations (Bell and Lightner 1984; Lightner 1996b, 1996c; Morales-Covarrubias and Chavez-Sanchez 1999; Motte et al. 2003). IHHNV can be transmitted through horizontally and vertically. The horizontal transmission is carried out by cannibalism and through contaminated water (Lightner 1996b; Lightner et al. 1983b, 1983c, 1985). The vertical transmission is caused by the infected eggs (Motte et al. 2003). IHHNV is believed to be enzootic in wild reservoir hosts (P. monodon) (Brock and Lightner 1990b), wild stock of P. stylirostris in the lower and upper Gulf of California (Pantoja et al. 1999), wild stock of adult male and female P. stylirostris from the mid-region of the Gulf of California (Morales-Covarrubias and Chavez-Sanchez 1999), wild stock of P. vannamei from the Pacific coast of Panama (Nunan et al. 2000) and wild stock of P. vannamei collected from the Pacific coasts of Ecuador, Colombia and Panama (Motte et al. 2003).
IHHNV infects and replicates in the tissues of ectodermal and mesodermal origin from the embryo. Thus, the IHHNV infects organs including gills, connective tissues, hypodermis, lymphoid organs, antennal gland and nerve cord and its ganglia (Fig. 4). The endodermal-derived tissues such as hepatopancreas, midgut and midgut mucosal epithelial and striated muscles have not showed any histological signs of infection with IHHNV (Lightner 1993, 1996b, 2011a).

Infectious hypodermal and haematopoietic necrosis virus. a, b Haematoxylin and eosin (H&E)-stained sections showing the presence of fixed phagocytes on myocardial fibres (source: Dhar et al. 2005). IHHNV-infected shrimp with deformed rostra; one curved down (c) and the other curved up (d) are marked by blue arrows. (Source: Dhar et al. 2014)
IHHNV is a stable shrimp virus and the smallest of the known penaeid shrimp viruses. The virion is a non-enveloped icosahedron with 20–22 nm diameter in size. It consists of the ssDNA genome with an estimated size of 3.9 to 4.1 kb and has a capsid composed of four polypeptides with different molecular weights of 74, 47, 39, and 37.5 kDa (Bonami et al. 1990; Nunan et al. 2000). Still now, two distinct genotypes of IHHNV have been reported: type 1 genotype from America and East Asia and type 2 genotype from Southeast Asia. These genotypes were reported to be infectious to P. vannamei and P. monodon (Tang et al. 2003). Two sequences homologous to part of the IHHNV genome are found to be present in the genome of penaeid. The homologous sequences were initially described as type 3A from India, Australia and East Africa and as type 3B from the Western Indo-Pacific region including Madagascar, Mauritius and Tanzania (Tang and Lightner 2006; Tang et al. 2007). Tissues containing the IHHNV-homologous sequences in the P. monodon genome are not infectious to the susceptible host species P. vannamei and P. monodon (Lightner et al. 2006; Tang and Lightner 2006; Tang et al. 2007).
Infectious myonecrosis virus
Infectious myonecrosis virus (IMNV) is the emerging viral disease, first reported in the Pacific white leg shrimp in the state of Piauí, northeast Brazil, in 2002 (Lightner et al. 2006). Followed by the first outbreak in Brazil, the disease spread to Southeast Asian countries such as Indonesia (Senapin et al. 2007) and India (Sahul Hameed et al. 2017). Initially, the disease was named as idiopathic myonecrosis (Lightner et al. 2004) and the disease was renamed as infectious myonecrosis. Finally, the aetiological agent was identified as virus and renamed as infectious myonecrosis virus (Poulos et al. 2006; Tang et al. 2008). Apart from L. vannamei, species like Litopenaeus stylirostris, P. monodon and Fenneropenaeus subtilis are shown to be experimentally susceptible (OIE 2009b). The virus infects all three life stages of the shrimp including post larva, juvenile and adult (Nunes et al. 2004). The cumulative mortality to IMNV reaches 40 to 70%. The target tissues for IMNV include the striated muscles (skeletal and less often cardiac), connective tissues, haemocytes and the lymphoid organ parenchymal cells (Tang et al. 2005). The IMNV infection is enzootic, and the signs of the disease in acute phase are characterised by focal to white necrotic areas in the striated muscles, particularly in the distal abdominal segment and tail fan. Infection with IMNV associated with heavy mortality may follow stressful events such as capture by cast-netting, feeding and sudden changes in water salinity or temperature (Poulos et al. 2006). IMNV has been demonstrated to be transmitted horizontally by cannibalism (Lightner 2011b; Poulos et al. 2006). Transmission via water probably occurs. The histological analysis of IMNV in acute phase is characterised by coagulative muscle necrosis, often with marked oedema among affected muscle fibres, fluid accumulation between muscle fibres, haemocytic infiltration, lymphoid organ spheroid formation and fibrosis (Fig. 5). Significant hypertrophy caused by accumulations of lymphoid organ spheroids (LOSs) is a highly consistent lesion in shrimp with acute- or chronic-phase infection with IMNV lesions. Common locations for ectopic LOS include the haemocoelom in the gills, heart, near the antennal gland tubules and ventral nerve cord.

Infectious myonecrosis virus. a IMNV-infected shrimp showing acute coagulative necrosis in the muscle fibres (mesoderm-derived tissue), the progression of myonecrosis giving affected muscle fibre a hyaline appearance, nuclear pyknosis, oedema and haemocytic infiltration. b Lymphoid organ spheroids (LOSs) (Source: Sunarto and Naim 2016). IHHNV-infected shrimp showing reddish opaque muscle at distal abdominal segments c naturally and d experimentally (Source: Sahul Hameed et al., 2017)
IMNV is likely belonging to be a member to Totiviridae family and is the first member of this family to infect a host other than fungus or protozoan (Nibert 2007). Phylogenetic analysis of its RNA-dependent RNA polymerase (RdRp) gene coding sequence indicates that IMNV is most closely related to Giardia lamblia virus, a member of the family Totiviridae (Fauquet et al. 2005; Lightner 2011a, 2011b). IMNV is a non-enveloped virion icosahedral in shape, with a diameter of 40 nm and a buoyant density of 1.366 g ml−1 in caesium chloride. The genome consists of a single, double-stranded RNA (dsRNA) molecule of 8226–8230 bp (Loy et al. 2015; Naim et al. 2015). The genome contains two non-overlapping ORFs. The first ORF (ORF1, nt 470–5596) encodes a putative RNA-binding protein and a capsid protein. The coding region of the RNA-binding protein is located in the first half of ORF1 and contains a dsRNA-binding motif in the first 60 amino acids. The second half of ORF1 encodes a capsid protein, as determined by amino acid sequencing, with a molecular mass of 106 kDa. The second ORF (ORF2, nt 5884–8133) encodes a putative RdRp (Poulos et al. 2006). The complete genomes of IMNV types originating from Brazil and Indonesia have been sequenced and found to be 99.6% identical at the nucleotide level (Poulos et al. 2006; Senapin et al. 2007). The 99.6% full-genome sequence identity indicates that the disease was introduced from Brazil to Indonesia in 2006.
Shrimp haemocyte iridescent virus
Shrimp haemocyte iridescent virus (SHIV) was first isolated and identified by Qiu et al. (2017) from the samples collected from the period 2014 to 2016 in Zhejiang Province in China. The disease caused high mortality in farmed L. vannamei in December 2014 in Zhejiang Province of China. It is confirmed that the farmed Fenneropenaeus chinensis and Macrobrachium rosenbergii were detected positive for SHIV disease from the collected samples from Zhejiang, Guangdong and Hebei provinces, China (Qiu et al. 2017). Based on the sequence of major capsid protein (MCP) (accession number KY681039) and ATPase (accession number KY681040), it has been indicated that SHIV virus belongs to the genus Xiairidovirus from Iridoviridae family. Iridoviridae group is characterised by large icosahedral viruses with diameters ranging from 120 to 300 nm. The virion is composed of the double-strand DNA genome ranging in size from 103 to 220 kbp (Qiu et al. 2017). The SHIV genome consists of a double-stranded DNA molecule composed of 165,809 bp. When compared to other members in the family Iridoviridae, the SHIV genome is larger than the members of the genera Ranavirus and Megalocytivirus and smaller than the members of the genera Iridovirus and Chloriridovirus. The G+C content of the SHIV genome is 34.6%. One hundred seventy ORFs were identified in the SHIV genome. Of this, 102 ORFs were in the forward orientation and 68 ORFs were in the reverse orientation. Eleven overlapping ORFs were found in the 170 ORFs of the SHIV genome (Shi et al. 2010).
SHIV infects tissues of ectodermal and mesodermal origin. Thus, the principal target organs include the gills, cuticular epithelium (or hypodermis), all connective tissues, haematopoietic tissues, lymphoid organ, antennal gland and the ventral nerve cord, its branches and its ganglia. L. vannamei exhibited the symptoms, including empty stomach and guts in all diseased shrimp, a slight loss of colour on the surface and section of hepatopancreas and soft shell in partially infected shrimp. One third of individuals had slightly reddish body. The moribund shrimp lost their swimming ability and sank to the bottom of water. The symptom and mortality were observed in the infected L. vannamei from post larvae to sub-adult shrimp. The histopathology analysis of SHIV-affected animal cephalothoraxes showed that karyopyknosis and basophilic inclusions existed in haematopoietic tissue and haemocytes in hepatopancreas, gills and periopods (Fig. 6). SHIV is a novel pathogen which had the potential to infect a lot of hosts, including shrimp, lobster, crayfish or other aquatic animals (Qiu et al. 2017, 2018).

Histopathology analysis of shrimp haemocyte iridescent virus (SHIV)-affected L. vannamei showing basophilic inclusion (black arrow) and karyopyknotic nuclei (white arrow). Haematoxylin and eosin (H&E) staining of a the haematopoietic tissue, b the gills, c the sinus in the hepatopancreas, and d the periopods (Qiu et al. 2017). Clinical signs of SHIV-affected animal (challenge group) compared with the control group (normal group). (a) External appearance of the shrimp. (b) Section of the hepatopancreas (Qiu et al. 2017)
Viral covert mortality disease
In 2009, serious disease outbreak caused heavy economic loss in China shrimp aquaculture industries (Zhang et al. 2017). The disease was commonly named as covert mortality disease (CMD) due to the moribund shrimp which died at the bottom of the pond instead at the surface of the water as the shrimp suffered from WSD (Zhang 2004; Gu 2012; Huang 2012). The diseased shrimp showed similar histopathology characters of IMNV, Macrobrachium rosenbergii nodavirus (MrNV) and Penaeus monodon nodavirus (PvNV). The disease-affected shrimps tested negative for IMNV, MrNV and PvNV by RT-PCR. Recently, a new emerging virus, covert mortality nodavirus (CMNV), was identified to be the aetiological agent of covert mortality disease. Now, the covert mortality disease was renamed as viral covert mortality disease (VCMD). The cumulative mortality of CMNV was variable, but it reaches 80 to 90% in some L. vannamei culture ponds (Xing 2004; Zhang 2004; Zhang et al. 2014). Apart from L. vannamei, species like Penaeus monodon, Fenneropenaeus chinensis, Marsupenaeus japonicas and Macrobrachium rosenbergii are shown to be CMNV positive by nested RT-PCR assay. Reverse transcription loop-mediated isothermal amplification (RT-LAMP) and RT-PCR assay showed that CMNV-positive samples were identified in brine shrimp (Artemia sinica), barnacle (Balanus sp.), rotifer (Brachionus urceus), amphipod (Corophium sinensis Zhang), Pacific oyster (Crassostrea gigas), hermit crab (Diogenes edwardsii), common clam (Meretrix lusoria), ghost crab (Ocypode cordimundus), hyperiid amphipod (Parathemisto gaudichaudi), fiddler crab (Tubuca arcuata) and an unidentified gammarid amphipod (Liu et al., 2018). After the first appearance in China, CMNV was also reported in Thailand (Thitamadee et al. 2016; Pooljun et al. 2016), Vietnam and Ecuador (Zhang et al. 2017).
CMNV is a non-enveloped, spherical-shaped, single-strand RNA virus, similar to other viruses in the family Nodaviridae. The virus consists of an icosahedral capsid size ranging from 29 to 35 nm in diameter. The capsid is constructed of 32 capsomers. The virion contains single-stranded RNA consisting of 4500 nucleotides. CMNV-infected shrimp shows hepatopancreatic atrophy and necrosis, empty stomach and gut, soft shell, slow growth of infected shrimp, abdominal muscle whitening and necrosis (Zhang 2004; Huang 2012). The histopathology analysis of CMNV-infected shrimp showed coagulative necrosis of striated muscle, eosinophilic inclusions in the epithelium of the tubules in the hepatopancreas and lymphoid organ, mass karyopyknotic nuclei which existed in the muscle and lymphoid organ and the tubular epithelium of the hepatopancreas showing significant atrophy (Fig. 7) (Zhang et al. 2014).

Viral covert mortality disease. a Haematoxylin and eosin (H&E) staining of the muscle showing the presence of necrotic muscle lesion in L. vannamei. b H&E staining of the hepatopancreatic tubular epithelium showing the eosinophilic inclusions. c H&E staining of the transection of abdominal necrotic muscle lesion in Macrobrachium rosenbergii. d H&E staining of the longitudinal section of abdominal necrotic muscle lesion in M. rosenbergii (Source: Zhang et al. 2017). e Clinical signs of CMNV-infected L. vannamei (Source: Huang et al. 2015)
Laem–Singh virus
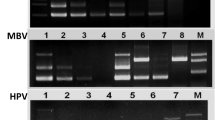
Laem–Singh virus (LSNV) was first identified in 2006 in Penaeus monodon shrimp reared at Laem-Singh District, Chanthaburi Province, Thailand, while investigating the cause of monodon slow growth syndrome (Sritunyalucksana et al. 2006). First, it has been assumed that LSNV is the aetiological agent for the cause of MSGS, but later, it was confirmed that LSNV is not the cause of MSGS because LSNV is detected in normal monodon shrimp and also LSNV was detected in L. vannamei culture stocks by RT-PCR which not retarded the growth of the shrimp. Meanwhile, it was found that LSNV was associated with retinopathy in small-sized shrimps from MSGS-affected ponds but it was not detected in large-sized shrimps from the same MSGS-affected ponds or in shrimps from normal ponds positive for LSNV by RT-PCR (Pratoomthai et al. 2008a). The reason for retinopathy was still unknown. Meanwhile, the presence of a novel integrase-containing element (ICE) along with LSNV was noticed in the retinal lesions of stunted shrimp from MSGS ponds in Thailand (Panphut et al. 2011). LSNV was reported in India in WSSV-, monodon baculovirus (MBV)- and HPV-affected Penaeus monodon shrimps (Prakasha et al. 2007), but it did not retard the growth of the shrimp (Kumar et al. 2011). LSNV was reported with MSGS in both healthy and infected P. monodon from Thailand, India (Poornima et al. 2012; Sathish Kumar et al. 2011), Sri Lanka (NACA), Indonesia, Malaysia and Vietnam. From the distribution of the disease, we came to know that LSNV is geographically restricted to South and Southeast Asia countries (Sittidilokratna et al. 2009). LSNV is a positive-sense single-stranded RNA (ssRNA), and the virion is non-enveloped and icosahedral in shape with a size ranging from 25 to 30 nm, similar to the insect-transmitted plant viruses in the family Luteoviridae.
LSNV was detected in P. monodon, Metapenaeus dobsoni, Fenneropenaeus merguiensis and L. vannamei. Still now, the natural host range of LSNV is unknown (Sathish Kumar et al., 2011). It has been reported that the LSNV can be transmitted vertically and horizontally because of its widespread detection in wild and domesticated broodstock and post-larval shrimp in Thailand and India (Raja et al., 2009; Saksmerprome et al. 2008). The virus affects the lymphoid organ of the shrimp. The in situ hybridisation (ISH) studies reveals the presence of LSNV in the cytoplasm of cells in the LO, heart, hepatopancreas and interstitial cells (Flegel and Withyachumnarnkul 2005; Flegel 2006; Pratoomthai et al. 2008b). The histopathological study of the LSNV with MSGS-positive shrimp shows the unusually large, magenta cytoplasmic inclusions in the LOSs (Fig. 8). Retinopathy was observed in the small shrimps only. Retinopathy is characterised by the abnormally enlarged haemolymphatic vessels, haemocytic infiltration in the fasciculated zone of the small shrimp and rupture of the membrane that separated the fasciculated zone from the overlying row of retinula cells (Poornima et al. 2012; Pratoomthai et al., 2008a, b; Raja et al., 2009).

a Haematoxylin and eosin (H&E) staining of the monodon baculovirus (MBV)-infected hepatopancreas of Penaeus monodon showing eosinophilic multiple inclusions of MBV (black arrow). b Enlarged view of the hepatopancreas of Penaeus monodon infected with monodon baculovirus stained with H&E showing the hypertrophied nuclei with marginated chromatin and inclusions of MBV (Source: Rajendran et al. 2012). c MBV-infected animal showing the pinkish colouration on the body (left) with normal shrimp
Monodon baculovirus
MBV has been first reported in 1977 in Penaeus monodon culture systems from Taiwan (Liao 1977). Later, it has been identified and reported from a variety of penaeid shrimps from different geographical areas throughout the world. Still now, MBV was reported in cultured shrimp in People’s Republic of China, Taiwan, Indonesia, Philippines, Malaysia, Thailand, Sri Lanka, Singapore, Australia, India, Israel, Kuwait, Oman, Italy, Kenya, Gambia, South Africa and USA (Baticados 1988; Baticados et al. 1991; Chen et al. 1989; Colorni 1989; Colorni et al. 1987; Alavandi et al. 1995; Felix and Devaraj 1993; Natividad and Lightner 1992; Ramasamy et al., 1995; Wang et al. 1997). MBV is otherwise called as Penaeus monodon singly enveloped nuclear polyhedrosis virus (PmSNPV). Besides MBV, still now two strains of SNPV were reported: Plebejus baculovirus and bennettae baculovirus. MBV has been reported from cultured or captured or imported shrimp species from different geographical regions. The primary host of the virus appeared to be Penaeus monodon, and it has been reported in Penaeus merguiensis, Penaeus semisulcatus, Penaeus kerathurus, Penaeus vannamei, Penaeus esculentus, Penaeus penicillatus, Penaeus plebejus, Metapenaeus ensis and Macrobrachium rosenbergii (Oanh and Phuong 2005; Vijayan et al., 1995a, b; Hao 1999; Chen et al. 1989; Lightner et al. 1987; Doubrovsky et al. 1988; Lightner 1988, 1996b; Colorni, 1989; Gangnonngiw et al. 2010; Manivannan et al. 2004). MBV has been reported in all stages in P. monodon. Late larval, post larval and young juvenile stages are the most susceptible stages (Brock and Lightner, 1990a). Lethargy, anorexia, dark colour and presence of heavy surface fouling are the morphological symptoms of the disease. Normally, the size of the MBV-infected shrimp is small compared to uninfected shrimps in the same culture pond. Hepatopancreatic tubule and duct epithelium in post larval, juvenile and adult stages and the anterior midgut epithelium of very young post larvae are the target organs for MBV. Acute MBV causes loss of hepatopancreatic tubule and midgut epithelia. The presence of multiple, spherical OBs in the hepatopancreas and anterior midgut epithelial cells is the prominent clinical sign of the disease (Fig. 9). Histopathological study shows the infected cells exhibit necrosis, lysis and sloughing of cells into tubule lumen (Brock and Lightner, 1990b; Baticados et al. 1991; Lightner and Redman 1981; Vijayan et al., 1995b).

Laem–Singh virus-infected animal showing dark discolouration and stunned growth (Source: Poornima et al. 2012)
MBV is a circular, dsDNA virus with a genome size in the range of 80–160 kbp. The genome size is reported to be similar to that of the dsDNA of insect baculoviruses (Mari et al. 1993). Taxonomic position of MBV is still uncertain, and the ICTV lists MBV (spherical baculoviruses) as a tentative species named PemoNPV in the genus Nucleopolyhedrovirus (Theilmann et al. 2005).
White tail disease
White tail disease (WTD) is also known as white muscle disease. The WTD has been first reported in the French West Indies (Arcier et al., 1999a). Later, the disease was reported from China (Qian et al., 2003a), India (Sahul Hameed et al., 2004), Thailand (Yoganandhan et al. 2006), Australia (Anonymous 2008; Owens et al. 2009), Taiwan (Wang et al. 2008) and Malaysia (Saedi Tayebeh et al. 2012). Two viruses (Macrobrachium rosenbergii nodavirus and extra small virus-like particle (XSV)) are found to be the aetiological agents causing white tail disease. Macrobrachium rosenbergii is the only host species for the WTD. But, pathogen-specific positive polymerase chain reaction (PCR) results (but not active infection) have been reported in white leg shrimp (Penaeus vannamei), kuruma prawn (Penaeus japonicus), Indian white prawn (Penaeus indicus), giant tiger prawn (Penaeus monodon), backswimmer (Notonecta sp.), hairy river prawn (Macrobrachium rude), monsoon river prawn (Macrobrachium malcolmsonii), brine shrimps (Artemia sp.) and red claw crayfish (Cherax quadricarinatus). Larval, post larval and early juvenile stages are the susceptible stages to WTD infection, whereas adults are resistant to the disease (Qian et al., 2003b; Sahul Hameed et al., 2004). Transmission of the disease occurs vertically (ovum) and horizontally (waterborne route) (Qian et al., 2003a; Sahul Hameed et al., 2004; Sudhakaran et al., 2007). The target organs for WTD infection include gill tissue, head muscle, heart, abdominal muscle, ovaries, pleopods and tail muscle. But, it did not infect the hepatopancreas and eyestalk (Sahul Hameed et al., 2004; Sri Widada et al. 2003). Pleopods are a convenient source of RNA for the screening of MrNV and XSV (Sahul Hameed et al., 2004). The WTD-infected post larvae show the following clinical signs: lethargy and opaqueness of the abdominal muscle starting at second or third abdominal segment and gradually extending from the centre to the anterior and the posterior sections of the muscle. Degeneration of the telson and uropods is observed in severe cases (Sahul Hameed and Bonami 2012). Mortality reaches a maximum within 5 to 6 days after appearance of whitish colouration. The infected PL show progressive weakening of their feeding and swimming ability (Sahul Hameed et al., 2004).
The histopathology analysis of the affected tissues in the infected PL shows the presence of acute Zenker’s necrosis of striated muscles, characterised by severe hyaline degeneration, necrosis and muscular lysis (Fig. 10). Large oval or irregular basophilic cytoplasmic inclusion bodies are seen in the target tissues (Arcier et al., 1999b; Hsieh et al. 2006; Sri Widada et al. 2003).

a Histological analysis of white tail disease (WTD)-infected M. rosenbergii showing the presence of acute necrosis in the skeletal muscles of shrimp b WTD-infected animal showing the whitish colouration on the body (right) with normal shrimp (left). (Source: Arcier et al. 1999a)
MrNV is a small, icosahedral, non-enveloped particle, 26–27 nm in diameter with two single-stranded RNAs: RNA1 (2.9 kb) and RNA2 (1.26 kb). Its capsid contains a single polypeptide of 43 kDa. XSV is a non-enveloped icosahedral virus, 15 nm in diameter with a linear ssRNA genome of 0.9 kb encoding two overlapping structural proteins of 16 and 17 kDa. XSV is a small size virus and does not have gene-encoding enzymes required for replication, and it depends upon the MrNV virus for their replication. So, it has been suggested that XSV may be a satellite virus and MrNV a helper virus (Sahul Hameed and Bonami 2012).
Conclusions
The decline of capture fisheries worldwide directly increases the importance of aquaculture products in the global market. Finfish and shellfish are the major culture organisms in most of the countries around the world. The demand for the culture fishes and shrimps in the international market gradually increases the intensity of the culture practice worldwide. This results in the failure of Good Manufacturing Practice (GMP) in the culture methods. In the last few decades, the ever increase in shrimp and fish culture activities in the developed and developing countries results in the formation and spreading of viral, bacterial and parasitic diseases and others.
Viral diseases are the major problem to the shrimp aqua farmers, which cause severe economic loss globally. In the 1990s, the disease white spot syndrome virus (WSSV) causes nearly seven billion dollars loss in Asian and American aquaculture industries. Even though IHHNV, YHV and TSV cause severe mortality and economic loss on the particular period, the loss was not much higher when compared to that by WSSV. From 1989 to 1994, viral and bacterial diseases cause a US$3019 million loss to the shrimp aqua farmers (Israngkura and Sae-Hae 2002). Recently, a new viral disease, shrimp haemocyte iridescent virus (SHIV), was identified in China in 2017, which caused severe mortality in the shrimp culture systems. To overcome the problem caused by diseases and to safeguard the shrimp aqua farmers from the severe economic loss, the following recommendations are suggested: (i) introduction of insurance policies to the aqua farmers, (ii) extension activities (experts’ coordination, investment of agencies, etc.), (iii) networking (field investigation, research, exchange of information, preventive measures), (iv) farm management (pond preparation, water quality, feed management, biosecurity etc.), (v) crop diversification (shrimp culture should be stopped, and different species culture were encouraged) and (vi) alternative income generation (explore and engage small-scale shrimp farmers to alternative works).
Disease surveillance is the most important work followed by the most of the developed countries and developing countries in the aquaculture regions. It helps to identify the presence, spreading and information about the disease in a particular region. The Network of Aquaculture Centres in Asia-Pacific (NACA) is an intergovernmental organisation that promotes rural development through sustainable aquaculture and aquatic resource management. The organisation improves the livelihoods of rural people, reduce poverty and increase food security. NACA is the nodal organisation for disease surveillance, which regulates and formulates the disease surveillance in most of the aquaculture countries through networking and coordination. Disease surveillance provides the complete details about the aquaculture status in a particular region, which are helpful to those countries importing culture organisms to clarify the presence of disease in the culture systems in the exporting countries. Disease surveillance is also helpful to the researchers to identify and prevent the spread of particular disease in a particular region.
-
Mandate to use certified (disease-free) broodstock (exotic and native) in all hatcheries
-
To develop origin traceability network systems for aquaculture species and their diseases
-
Even though there are epidemic shrimp diseases, OIE has to edify them and they have to provide auxiliary funds to develop diagnostic kits on those diseases
-
Regular surveillance of culturing ponds which will facilitate to map the viral hotspot areas to be focused by the amendable government bodies
-
Effective regulation and on-spot routine monitoring of infective ponds and their surrounding ponds
-
Molecular screening of imported aquaculture products specific to live feeds
-
We have to develop in-depth educational videos about good farm practices, aetiology and impact of the virus on shrimps and re-habitation complications-cum-expenses to farmers and economy diminution to the sector which has to projected through mobile digital big screens while conducing on-sight awareness camps
References
Ahmed ATA (1996) Disease problems of cultured tiger shrimp (Penaeus monodon) in Bangladesh. 2nd Int. Conference on the Culture of Penaeid Prawns and Shrimps, book of abstracts, SEAFDEC/AQD, Iloilo City
Alavandi SV, Vijayan KK, Rajendran KV (1995) Shrimp diseases, their prevention and control. In: Central Institute of Brackishwater Aquaculture (CIBA) Bulletin, vol 3, pp 1–17
Anonymous (2008) White tail disease in freshwater prawn. wwwTheFishSitecom March 2008
Arcier JM, Herman F, Lightner DV, Redman R, Mari J, Bonami JR (1999a) A viral disease associated with mortalities in hatchery-reared postlarvae of the giant freshwater prawn Macrobrachium rosenbergii. Dis Aquat Org 38:177–181. https://doi.org/10.3354/dao038177
Arcier JM, Herman F, Lightner DV, Redman R, Mari J, Bonami JR (1999b) A viral disease associated with mortalities in hatchery-reared post larvae of the giant freshwater prawn Macrobrachium rosenbergii. Dis Aquat Org 38:177–181. https://doi.org/10.3354/dao038177
Bateman KS, Tew I, French C, Hicks RJ, Martin P, Munro J, Stentiford GD (2012) Susceptibility to infection and pathogenicity of white spot disease (WSD) in non-model crustacean host taxa from temperate regions. J Invertebr Pathol 110(3):340–351
Baticados MCL (1988) Diseases of prawns in Philippines. SEAFDEC Asian Aquaculture 10:1–8
Baticados MCL, Lavilla-Pitogo CR, Paner MG, de la Pena LD, Tendencia EA (1991) Occurrence and pathology of Penaeus monodon baculovirus infection in hatcheries and ponds in Philippines. Bamidegh 43:35–41
Bell TA, Lightner DV (1984) IHHN virus: infectivity and pathogenicity studies in Penaeus stylirostris and Penaeus vannamei. Aquaculture 38:185–194. https://doi.org/10.1016/0044-8486(84)90142-X
Bonami JR, Trumper B, Mari J, Brehelin M, Lightner DV (1990) Purification and characterization of the infectious hypodermal and hematopoietic necrosis virus of penaeid shrimps. J Gen Virol 71:2657–2664. https://doi.org/10.1099/0022-1317-71-11-2657
Bonami JR, Hasson KW, Mari J, Poulos BT, Lightner DV (1997) Taura syndrome of marine penaeid shrimp: characterization of the viral agent. J Gen Virol 78(2):313–319
Brock JA, Lightner DV (1990a) Chapter 3: diseases of Crustacea. In: Kinne O (ed) Diseases of marine animals vol. 3. Biologische Anstalt Helgoland, Hamburg, pp 245–424
Brock JA, Lightner DV (1990b) Diseases of crustacea. Diseases caused by microorganisms. In: Kinne O (ed) Diseases of marine animals, vol 3. Biologische Anstalt Helgoland, Hamburg, pp 245–349
Chang PS, Chen HC, Wang YC (1998) Detection of white spot syndrome associated baculovirus in experimentally infected wild shrimp, crabs and lobsters by in situ hybridization. Aquaculture 164:233–242. https://doi.org/10.1016/S0044-8486(98)00189-6
Chantanachookin C, Boonyaratpalin S, Kasornchandra J, Direkbusarakom S, Aekpanithanpong U, Supamataya K, Sriurairatana S, Flegel TW (1993) Histology and ultrastructure reveal a new granulosis-like virus in Penaeus monodon affected by yellow-head disease. Dis Aquat Org 17:145–157
Chayaburakul K, Nash G, Pratanpipat P, Sriurairatana S, Withyachumnarnkul B (2004) Multiple pathogens found in growth-retarded black tiger shrimp Penaeus monodon cultivated in Thailand. Dis Aquat Org 60:89–96
Chen SN (1995) Current status of shrimp aquaculture in Taiwan. In: Browdy CL, Hopkins JS (eds) Swimming through troubled water. Proceedings of the special session on shrimp farming. Aquaculture ’95. World Aquaculture Society, Baton Rouge, pp 29–34
Chen SN, Chang PS, Kou GH, Lightner DV (1989) Studies on virogenesis and cytopathology of Penaeus monodon baculovirus (MBV) in the giant tiger prawn (Penaeus monodon) and the red tail prawn (Penaeus penecillatus). Fish Pathol 24:89–100. https://doi.org/10.3147/jsfp.24.89
Cheng LT, Lin WH, Wang PC, Tsai MA, Ho PY, Hsu JP, Chern RS, Chen SC (2011) Epidemiology and phylogenetic analysis of Taura syndrome virus in cultured Pacific white shrimp Litopenaeus vannamei B. in Taiwan. Dis Aquat Org 97:17–23. https://doi.org/10.3354/dao02407
Colorni A (1989) Penaeid pathology in Israel: problems and research. In: Advances in tropical aquaculture, Tahiti. Actes de Colloque 9 AQUACOP IFREMER. pp. 89–96
Colorni A, Samocha T, Colorni B (1987) Pathogenic viruses introduced into mariculture systems by imported penaeid shrimp. Bamidgeh 39:21–28
Cowley JA, Walker PJ, Flegel TW, Lightner DV, Bonami JR, Snijder EJ, De Groot RJ (2012) Family Roniviridae. In: King A, Adams M, Carstens E, Lefkowitz EJ (eds) Virus taxonomy, IXth report of the International Committee on Taxonomy of Viruses. Elsevier, Academic, London, pp 797–801
Dhar AK, Lakshman DK, Amundsen K, Robles-Sikisaka R, Kaizer KN, Roy S, Hasson KW, Thomas Allnutt FC (2010) Characterization of a Taura syndrome virus isolate originating from the 2004 Texas epizootic in cultured shrimp. Arch Virol 155(3):315–327
Dhar AK, Robles-Sikisaka R, Saksmerprome V, Lakshman DK (2014) Biology, genome organization, and evolution of parvoviruses in marine shrimp. Adv Virus Res 89:85–139. https://doi.org/10.1016/b978-0-12-800172-1.00003-3
Doubrovsky A, Paynter JL, Sambhi SK, Atherton JG, Lester RJG (1988) Observations on the ultrastructure of baculovirus in Australian Penaeus monodon and Penaeus merguiensis. Aust J Mar Freshw Res 39:743–749. https://doi.org/10.1071/MF9880743
FAO (Food and Agriculture Organization) (1997) Aquaculture development technical guidelines for responsible fisheries no. 5. FAO, Rome, 40
FAO (Food and Agriculture Organization) (2010) The State of World Fisheries and Aquaculture Food and Agriculture Organization of the United Nations. Rome, FAO, 244
Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA (2005) Totiviridae. In: Virus taxonomy: classification and nomenclature of viruses. Eighth report of the International Committee on the Taxonomy of Viruses. Elsevier, San Francisco, pp 571–580
Felix S, Devaraj M (1993) Incidence of destructive MBV and IHHNV in a commercial shrimp hatchery. Seaf Export J 25:13–18
Flegel TW (2006) Detection of major penaeid shrimp viruses in Asia, a historical perspective with emphasis on Thailand. Aquaculture 258:1–33. https://doi.org/10.1016/j.aquaculture.2006.05.013
Flegel TW and Withyachumnarnkul B (2005) Research progress on monodon slow growth syndrome (MSGS), World Aquaculture Meeting in Thailand
Gangnonngiw W, Laisutisan K, Sriurairatana S, Senapin S, Chuchird N, Limsuwan C, Chaivisuthangkura P, Flegel TW (2010) Monodon baculovirus (MBV) infects the freshwater prawn Macrobrachium rosenbergii cultivated in Thailand. Virus Res 148:24–30. https://doi.org/10.1016/j.virusres.2009.12.001
Gu SJ (2012) Analysis of causes of the covert mortality disease of Pacific white shrimp and its control strategies. Sci Fish Farming 8:62–63 (in Chinese)
Hao NV (1999) Study on diseases in Penaeus monodon cultured in Tra Vinh Province, Vietnam. Report of UNDP project (VIE/96/025)
Hsieh CY, Chuang PC, Chen LC, Tu C, Chien MS, Huang KC, Kao HF, Tung MC, Tsai SS (2006) Infectious hypodermal and haematopoietic necrosis virus (IHHNV) infections in giant freshwater prawn, Macrobrachium rosenbergii. Aquaculture 258:73–79. https://doi.org/10.1016/j.aquaculture.2006.04.007
Huang J (2012) Experience in EMS/AHPNS from China. In: NACA Bangkok: Network of Aquaculture Centres in Asia-Pacific (ed) Report of the Asia Pacific emergency regional consultation on the emerging shrimp disease: early mortality syndrome (EMS)/acute hepatopancreatic necrosis syndrome (AHPNS), pp 21–23
Huang J, Song X-L, Yu J, Yang CH (1994) Baculoviral hypodermal and hematopoietic necrosis - pathology of the shrimp explosive epidemic disease. Abstract, Yellow Sea Fishery Research Institute, Qingdao Cited by Lightner, 1996
Huang S-P, Shen C-N, Chen I-S (2015) The complete mitochondrial genome of the Abe’s mangrove goby (Teleostei, Gobiidae). Mitochondrial DNA 26(1):143–144
Inouye K, Miwa S, Oseko N, Nakano H, Kimura T, Momoyama K, Hiraoka M (1994) Mass mortalities of cultured Kuruma shrimp Penaeus japonicus in Japan in 1993: electron microscope evidence of the causative virus. Fish Pathol 29:149–158
Israngkura A, Sae-Hae S (2002) A review of the economic impacts of aquatic animal disease. In: Arthur JR, Phillips MJ, Subasinghe RP, Reantaso MB, MacRae IH (eds) Primary aquatic animal health care in rural, small-scale, aquaculture development, pp 253–286 FAO Fish Tech Pap No. 406
Jimenez R (1992) Sindrome de Taura (Resumen). In: Acuacultura del Ecuador. Camara Nacional de Acuacultura, Guayaquil, pp 1–16
Khongpradit R, Kasornchandra J, Boonyaratpalin S (1995) Susceptibility of the postlarval stages of black tiger shrimp (Penaeus monodon) to yellow-head baculovirus (YBV). In: Shariff M, Subasinghe RP, Arthur JR (eds) Diseases in Asian aquaculture II. Asian Fisheries Society, Manila, p 6
Krabsetsve K, Cullen BR, Owens L (2004) Rediscovery of the Australian strain of infectious hypodermal and haematopoietic necrosis virus. Dis Aquat Org 61:153–158. https://doi.org/10.3354/dao061153
Kumar TS, Krishnan P, Makeshn M, Chaudhari A, Purushothaman CS, Rajendran KV (2011) Natural host-range and experimental transmission of Laem-Singh virus (LSNV). Dis Aquat Org 96:21–27. https://doi.org/10.3354/dao02374
Liao I (1977) A culture study on grass prawn, Penaeus monodon, in Taiwan- the patterns, the problems and the prospects. J Fish Soc Taiwan 5:11–29
Lightner DV (1988) Diseases of cultured penaeid shrimp and prawns. In: Sinderman CJ, Lightner DV (eds) Disease diagnosis and control in North American marine aquaculture. Elsevier, Amsterdam, pp 8–127
Lightner DV (1993) Diseases of penaeid shrimp. In: McVey JP (ed) CRC handbook of mariculture: crustacean aquaculture. CRC, Boca Raton
Lightner DV (1996a) Handbook of pathology and diagnostic procedures for diseases of penaeid shrimp. World Aquaculture Society, Baton Rouge
Lightner DV (1996b) A handbook of shrimp pathology and diagnostic procedures for diseases of cultured penaeid shrimp. World Aquaculture Society, Baton Rouge
Lightner DV (1996c) The penaeid shrimp viruses IHHNV and TSV: epizootiology, production impacts and role of international trade in their distribution in the Americas. Revue Scientifique et Technique Office International des Epizooties 15:579–601
Lightner DV (2011a) Status of shrimp diseases and advances in shrimp health management. In: Bondad-Reantaso MG, Jones JB, Corsin F, Aoki T (eds) Diseases in Asian aquaculture VII. Fish Health Section, Asian Fisheries Society, Selangor, pp 121–134
Lightner DV (2011b) Virus diseases of farmed shrimp in the Western Hemisphere (the Americas): a review. J Invertebr Pathol 106:110–130. https://doi.org/10.1016/j.jip.2010.09.012
Lightner DV, Redman RM (1981) A baculovirus-caused disease of the penaeid shrimp, Penaeus monodon. J Invertebr Pathol 38:299–302. https://doi.org/10.1016/0022-2011(81)90137-3
Lightner DV, Redman RM (1998) Shrimp diseases and current diagnostic methods. Aquaculture 164:201–220. https://doi.org/10.1016/S0044-8486(98)00187-2
Lightner DV, Redman RM, Bell TA, Brock JA (1983a) Detection of IHHN virus in Penaeus stylirostris and Penaeus vannamei imported into Hawaii. J World Maricult Soc 14:212–225. https://doi.org/10.1111/j.1749-7345.1983.tb00077.x
Lightner DV, Redman RM, Bell TA (1983b) Infectious hypodermal and hematopoietic necrosis a newly recognised virus disease in penaeid shrimp. J Invertebr Pathol 42:62–70. https://doi.org/10.1016/0022-2011(83)90202-1
Lightner DV, Redman RM, Williams RR, Mohney LL, Clerx JPM, Bell TA, Brock JA (1985) Recent advances in penaeid virus disease investigations. J World Maricult Soc 16:267–274. https://doi.org/10.1111/j.1749-7345.1985.tb00209.x
Lightner DV, Hedrick RP, Fryer JL, Chen SN, Lia IC, Kou GH (1987) A survey of cultured penaeid shrimps in Taiwan for viral and other important diseases. Fish Pathol 22:127–140. https://doi.org/10.3147/jsfp.22.127
Lightner DV, Redman RM, Hasson KW, Pantoja CR (1995a) Taura syndrome in Penaeus vannamei: histopathology and ultrastructure. Dis Aquat Org 21:53–59. https://doi.org/10.3354/dao021053
Lightner DV, Redman RM, Hasson KW, Pantoja CR (1995b) Taura syndrome in Penaeus vannamei (Crustacea: Decapoda): gross signs, histopathology and ultrastructure. Dis Aquat Org 21:53–59. https://doi.org/10.3354/dao021053
Lightner DV, Pantoja CR, Poulos BT, Tang KFJ, Redman RM, Andreas T, Bonami JR (2004) Infectious myonecrosis (IMN): a new virus disease of Litopenaeus vannamei. In: Aquaculture 2004 book of abstracts, Baton Rouge
Lightner DV, Poulos BT, Tang-Nelson KF, Pantoja CR, Nunan LM, Navarro SA, Redman RM, Mohney LL (2006) Application of molecular diagnostic methods to penaeid shrimp diseases: advances of the past 10 years for control of viral diseases in farmed shrimp. Dev Biol (Basel). 126: 117–122
Limsuwan C (1993) Diseases of black tiger shrimp, Penaeus monodon Fabricius in Thailand. In: Akiyama DM (ed) Technical bulletin of the American Soybean Association. American Soybean Association, Singapore, pp 1–22
Liu Q, Huang J, Yang HL, Yang B, Want HL, Want QT, Liu F, Zhang QL (2014) Detection of a new genotype of yellow-head virus in farmed shrimp suspicious of EMS/AHPNS infection. Oceanol Limnol Sin 45:703–709. https://doi.org/10.11693/hyhz20130500045
Liu S, Wang X, Xu T, Li X, Du L, Zhang Q (2018) Vectors and reservoir hosts of covert mortality nodavirus (CMNV) in shrimp ponds. J Invertebr Pathol 154:29–36. https://doi.org/10.1016/j.jip.2018.03.011
Lo CF, Wang CH, Kou GH (1995) Purification and genomic analysis of white spot syndrome associated non-occluded baculovirus (PmNOB) isolated from Penaeus monodon. Abstract, European Association of Fish Pathologists, 7th International Conference on Diseases of Fish and Shellfish, Spain. p. 77
Lo CF, Aoki T, Bonami JR, Flegel TW, Leu JH, Lightner DV, Stentiford G, Soderhall K, Walker PW, Wang HC, Xun X, Yang F, Vlak JM (2012) Nimaviridae. In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (eds) Virus taxonomy: classification and nomenclature of viruses: ninth report of the International Committee on Taxonomy of Viruses. Elsevier Academic, San Diego, pp 229–234
Loy DS, Liu S, Mogler MA, Loy DJ, Blitvich BJ, Bartholomay LC (2015) Characterization of newly revealed sequences in the infectious myonecrosis virus genome in Litopenaeus vannamei. J Gen Virol 96:1821–1829. https://doi.org/10.1099/vir.0.000137
Lundin CG (1995) Global attempts to address shrimp disease. Report of the land, water and natural habitats division, environment department. The World Bank, Washington
Maeda M, Kasornchandra J, Itami T, Suzuki N, Hennig O, Kondo M, Albaladejo JD, Takahashi Y (1998) Effect of various treatments on white spot syndrome virus (WSSV) from Penaeus japonicus (Japan) and P. monodon (Thailand). Fish Pathol 33:381–387
Manivannan S, Kennedy B, Karunasagar I, Karunasagar I (2004) Prevalence of monodon baculovirus in wild Metapenaeus species along the southwest coast of India. Aquaculture 232:63–67. https://doi.org/10.1016/S0044-8486(03)00461-7
Mari J, Bonami JR, Poulos B, Lightner DV (1993) Preliminary characterization and partial cloning of the genome of a baculovirus from Penaeus monodon (PmSNPV = MBV). Dis Aquat Org 16:207–215. https://doi.org/10.3354/dao016207
Mari J, Bonami JR, Lightner DV (1998) Taura syndrome of penaeid shrimp: cloning of viral genome fragments and development of specific gene probes. Dis Aquat Org 33:11–17. https://doi.org/10.3354/dao033011
Mari J, Poulos BT, Lightner DV, Bonami JR (2002) Shrimp Taura syndrome virus: genomic characterization and similarity with members of the genus Cricket paralysis-like viruses. J Gen Virol 83:915–926. https://doi.org/10.1099/0022-1317-83-4-915
Mohr PG, Moody NJG, Hoad J, Williams LM, Bowater RO, Cummins DM, Cowley JA, Crane MSTJ (2015) New yellow head virus genotype (YHV7) in giant tiger shrimp Penaeus monodon indigenous to northern Australia. Dis Aquat Org 115:263–268. https://doi.org/10.3354/dao02894
Morales-Covarrubias M, Chavez-Sanchez MC (1999) Histopathological studies on wild broodstock of white shrimp Penaeus vannamei in the Platanitos area, adjacent to San Blas, Nayarit, Mexico. J World Aquacult Soc 30:192–200. https://doi.org/10.1111/j.1749-7345.1999.tb00866.x
Motte E, Yugcha E, Luzardo J, Castro F, Leclercq G, Rodríguez J, Miranda P, Borja O, Serrano J, Terreros M, Montalvo K, Narvaez A, Tenorio N, Cedeno V, Mialhe E, Boulo V (2003) Prevention of IHHNV vertical transmission in the white shrimp Litopenaeus vannamei. Aquaculture 219:57–70. https://doi.org/10.1016/S0044-8486(02)00631-2
Naim S, Tang KFJ, Yang M, Lightner DV, Nibert ML (2015) Extended genome sequences of penaeid shrimp infectious myonecrosis virus strains from Brazil and Indonesia. Arch Virol 160:1579–1158. https://doi.org/10.1007/s00705-015-2406-2
Natividad JM, Lightner DV (1992) Susceptibility of the different larval and postlarval stages of the black tiger prawn, Penaeus monodon Fabricius, to monodon baculovirus (MBV). In: Shariff M, Subasinghe RP, Arthur JR (eds) Diseases in Asian aquaculture 1, Fish Health Section, Asian Fisheries Society, Manila, pp 111–125
Ndanga LZB, Quagrainie KK, Dennis JH (2013) Economically feasible options for increased women participation in Kenyan aquaculture value chain. Aquaculture 414–415:183–190. https://doi.org/10.1016/j.aquaculture.2013.08.012
Nibert ML (2007) ‘2A-like’ and ‘shifty heptamer’ motifs in penaeid shrimp infectious myonecrosis virus, a monosegmented double-stranded RNA virus. J Gen Virol 88:1315–1318. https://doi.org/10.1099/vir.0.82681-0
Nielsen L, Sang-Oum W, Cheevadhanarak S, Flegel TW (2005) Taura syndrome virus (TSV) in Thailand and its relationship to TSV in China and the Americas. Dis Aquatic Org 63:101–106. https://doi.org/10.3354/dao063101
Nunan LM, Poulos BT, Lightner DV (2000) Use of polymerase chain reaction for the detection of infectious hypodermal and hematopoietic necrosis virus in penaeid shrimp. Mar Biotechnol 2:319–328. https://doi.org/10.1007/s101260000003
Nunes AJP, Cunha-Martins P, Vasconselos-Gesteira TC (2004) Carcinicultura ameaçada. Rev Panoram Aquic 83:37–51 (in Portuguese)
Oanh DTH, Phuong NT (2005) Prevalence of white spot syndrome virus (WSSV) and monodon baculovirus (MBV) infection in Penaeus monodon postlarvae in Vietnam. In: Walker P, Lester R, Bondad-Reantaso MG (eds) Diseases in Asian aquaculture V. Fish Health Section, Asian Fisheries Society, Manila, pp 395–404
OIE - World Organisation for Animal Health (2009a) Manual of diagnostic tests for aquatic animals. OIE, Paris
OIE - World Organisation for Animal Health (2009b) Manual of diagnostic tests for aquatic animals 2009. OIE, Paris
Owens L, Anderson IG, Kenway M, Trott L, Benzie JAH (1992) Infectious hypodermal and hematopoietic necrosis virus (IHHNV) in a hybrid penaeid prawn from tropical Australia. Dis Aquat Org 14:219–228
Owens L, La Fauce K, Juntunen K, Hayakijkosol O, Zeng C (2009) Macrobrachium rosenbergii nodavirus disease (white tail disease) in Australia. Dis Aquat Org 85:175–180. https://doi.org/10.3354/dao02086
Panphut W, Senapin S, Sriurairatana S, Withyachumnarnkul B, Flegel TW (2011) A novel integrase-containing element may interact with Laem-Singh virus (LSNV) to cause slow growth in giant tiger shrimp. BMC Vet Res 7:18. https://doi.org/10.1186/1746-6148-7-18
Pantoja CR, Lightner DV, Holtschmit KH (1999) Prevalence and geographic distribution of Infectious Hypodermal and Hematopoietic Necrosis Virus (IHHNV) in wild blue shrimp from the gulf of California, Mexico. J Aquat Anim Health 11(1):23–34
Pooljun C, Direkbusarakom S, Chotipuntu P, Hirono I, Wuthisuthimethavee S (2016) Development of a TaqMan real-time RT-PCR assay for detection of Covert Mortality Nodavirus (CMNV) in penaeid shrimp. Aquaculture 464:445–450
Poornima M, Alavandi SV, Kalaimani N (2012) Laem-Singh virus (LSNV) and monodon slow growth syndrome in farmed black tiger shrimp (Penaeus monodon): an update. In: Pillai SM et al., editors. National conference on new vistas in indian aquaculture—book of abstracts. Chennai: Coastal Aquaculture Society of India, Chennai and Central Institute of Brackishwater Aquaculture, pp. 49–50
Poulos BT, Tang KFJ, Pantoja CR, Bonami JB, Lightner DV (2006) Purification and characterization of infectious myonecrosis virus of penaeid shrimp. J GenVirol 87:987–996. https://doi.org/10.1099/vir.0.81127-0
Prakasha BK, Ramakrishna RP, Karunasagar I, Karunasagar I (2007) Detection of Laem-Singh virus (LSNV) in cultured Penaeus monodon from India. Dis Aquat Org 77:83–86. https://doi.org/10.3354/dao01835
Pratoomthai B, Sakaew W, Sriurairatana S, Wongprasert K, Withyachumnarnkul B (2008a) Retinopathy in stunted black tiger shrimp Penaeus monodon and possible association with Laem-Singh virus (LSNV). Aquaculture 284:53–58. https://doi.org/10.1016/j.aquaculture.2008.07.040
Pratoomthai B, Wongprasert K, Flegel WT, Withyachumnarnkul B (2008b) Infection by Laem-Singh virus in the fasciculated zone and organ of Bellonci of the eyes of small Penaeus monodon from monodon slow-growth syndrome pond. In: 7th symposium on diseases in Asian aquaculture at Taipei, Taiwan
Qian D, Shi Z, Zhang S, Cao Z, Liu W, Li L, Xie Y, Cambournac I, Bonami JR et al (2003a) Extra small virus-like particles (XSV) and nodavirus associated with whitish muscle disease in the giant freshwater prawn, Macrobrachium rosenbergii. J Fish Dis 26:521–527. https://doi.org/10.1046/j.1365-2761.2003.00486.x
Qian D, Shi Z, Zhang S, Cao Z, Liu W, Li L, Xie Y, Cambournac I, Bonami JR (2003b) Extra small virus-like particles (XSV) and nodavirus associated with whitish muscle disease in the giant fresh water prawn, Macrobrachium rosenbergii. J Fish Dis 26:521–527. https://doi.org/10.1046/j.1365-2761.2003.00486.x
Qiu L, Chen MM, Wan XY, Li C, Zhang QL, Wang RY, Cheng DY, Dong X, Yang B, Wang XH, Xiang JH, Huang J (2017) Characterization of a new member of Iridoviridae, shrimp hemocyte iridescent virus (SHIV), found in white leg shrimp (Litopenaeus vannamei). Sci Rep 7:11834. https://doi.org/10.1038/s41598-017-10738-8
Qiu L, Chen MM, Wang RY, Wan XY, Li C, Zhang QL, Dong X, Yang B, Xiang JH, Huang J (2018) Complete genome sequence of shrimp hemocyte iridescent virus (SHIV) isolated from white leg shrimp, Litopenaeus vannamei. Arch Virol 163:781–785. https://doi.org/10.1007/s00705-017-3642-4
Rai P, Pradeep B, Karunasagar I, Karunasagar I (2009) Detection of viruses in Penaeus monodon from India showing signs of slow growth syndrome. Aquaculture 289:231–235. https://doi.org/10.1016/j.aquaculture.2008.12.035
Raja JJS, Sivakumar S, Singaravel R, Poornima M, Alavandi SV, Kalaimani N, Santiago TC (2009) Detection of slow growth syndrome disease in farmed shrimp in India. In: Indian Youth Science Congress (IYSC 2009) at Rajiv Gandhi National Institute of Youth Development, Sriperumbudur
Rajendran KV, Makesh M, Karunasagar I (2012) Monodon baculovirus of shrimp. Indian J Virol 23(2):149–160
Ramasamy P, Brennan GP, Jayakumar R (1995) A record and prevalence of monodon baculovirus from postlarval Penaeus monodon in Madras. India Aquac 130:129–135
Robles-Sikisaka R, Garcia DK, Klimpel KR, Dhar AK (2001) Nucleotide sequence of 3′-end of the genome of Taura syndrome virus of shrimp suggests that it is related to insect picornaviruses. Arch Virol 146:941–952. https://doi.org/10.1007/s007050170126
Rosenberry B (1996) World shrimp farming. Shrimp News International, San Diego
Rosenberry B (2000) World shrimp farming. Shrimp News International, San Diego
Rosenberry B (2002) World shrimp farming. Shrimp News International, San Diego
Saedi Tayebeh A, Moeini H, Wen Tan S, Yusoff K, Hassan Daud M, Kua Chu B, Soon Tan G, Bhassu S (2012) Detection and phylogenetic profiling of nodavirus associated with white tail disease in Malaysian Macrobrachium rosenbergii (de Man). Mol Biol Rep 39:5785–5790. https://doi.org/10.1007/s11033-011-1389-7
Sahul Hameed S, Bonami JR (2012) White tail disease of freshwater prawn, Macrobrachium rosenbergii. Indian J Virol 23:134–140. https://doi.org/10.1007/s13337-012-0087-y
Sahul Hameed AS, Yoganandhan K, Sri Widada J, Bonami JR (2004) Studies on the occurrence and RT-PCR detection of Macrobrachium rosenbergii nodavirus and extra small virus-like particles associated with white tail disease of Macrobrachium rosenbergii in India. Aquaculture 238:127–133. https://doi.org/10.1016/j.aquaculture.2004.06.009
Sahul Hameed AS, Abdul Majeed S, Vimal S, Nithiyanandham M, Rajkumar T, Santhoshkumar S, Sivakumar S (2017) Studies on the occurrence of infectious myonecrosis virus in pond-reared Litopenaeus vannamei (Boone, 1931) in India. J Fish Dis 40:1823–1830
Saksmerprome V, Wongtripop S, Noonin C, Withyachumnarnkul B (2008) Double-stranded RNA induces interference with Lame-Singh virus (LSNV) replication in penaeid shrimp. In: 7th symposium on diseases in Asian aquaculture at Taipei, Taiwan
Sanchez-Barajas M, Linan-Cabello MA, Mena-Herrera A (2009) Detection of yellow-head disease in intensive freshwater production systems of Litopenaeus vannamei. Aquacult Int 17:101–112. https://doi.org/10.1007/s10499-008-9183-9
Sathish Kumar T, Krishnan P, Makesh M, Chaudhari A, Purushothaman CS, Rajendran KV (2011) Natural host-range and experimental transmission of Laem-Singh virus (LSNV). Dis Aquatic Org 96:21–27
Senapin S, Phewsaiya K, Briggs M, Flegel TW (2007) Outbreaks of infectious myonecrosis virus (IMNV) in Indonesia confirmed by genome sequencing and use of an alternative RT-PCR detection method. Aquaculture 266(1–4):32–38
Senapin S, Thaowbut Y, Gangnonngiw W, Chuchird N, Sriurairatana S, Flegel TW (2010) Impact of yellow head virus outbreaks in the whiteleg shrimp, Penaeus vannamei (Boone), in Thailand. J Fish Dis 33:421–430. https://doi.org/10.1111/j.1365-2761.2009.01135.x
Shi CY, Jia KT, Yang B, Huang J (2010) Complete genome sequence of a Megalocytivirus (family Iridoviridae) associated with turbot mortality in China. Virol J 7:1–9. https://doi.org/10.1186/1743-422X-7-159
Sittidilokratna N, Dangtip S, Cowley JA, Walker PJ (2008) RNA transcription analysis and completion of the genome sequence of yellow head nidovirus. Virus Res 136:157–165. https://doi.org/10.1016/j.virusres.2008.05.008
Sittidilokratna N, Dangtip S, Sritunyalucksana K, Babu R, Pradeep B, Mohan CV, Gudkovs N, Walker PJ (2009) Detection of Laem-Singh virus in cultured Penaeus monodon shrimp from several sites in the Indo-Pacific region. Dis Aquat Org 84:195–200
Spann KM, Vickers JE, Lester RJG (1995) Lymphoid organ virus of Penaeus monodon from Australia. Dis Aquat Org 23:127–134
Sri Widada J, Durand S, Cambournac I, Qian D, Shi Z, Dejonghe E, Richard V, Bonami JR (2003) Genome-based detection methods of Macrobrachium rosenbergii nodavirus, a pathogen of the giant fresh water prawn, Macrobrachium rosenbergii: dot-blot, in situ hybridization and RT-PCR. J Fish Dis 26:583–590. https://doi.org/10.1046/j.1365-2761.2003.00493.x
Sritunyalucksana K, Apisawetakan S, Boonnat A, Withyachumnarnkul B, Flegel TW (2006) A new RNA virus found in black tiger shrimp Penaeus monodon from Thailand. Vir Res 118:31–38. https://doi.org/10.1016/j.virusres.2005.11.005
Subasinghe R, Soto D, Jia J (2009) Global aquaculture and its role in sustainable development. Rev Aquacult 1:2–9. https://doi.org/10.1111/j.1753-5131.2008.01002.x
Sudhakaran R, Ishaq Ahmed VP, Haribabu P, Mukherjee SC, Sri Widada J, Bonami JR, Sahul Hameed AS (2007) Experimental vertical transmission of Macrobrachium rosenbergii nodavirus (MrNV) and extra small virus (XSV) from brooders to progeny in Macrobrachium rosenbergii and Artemia. J Fish Dis 30:27–35. https://doi.org/10.1111/j.1365-2761.2007.00774.x
Sunarto A, Naim S (2016) Totiviruses of crustaceans. In: Kibenge FSB, Godoy MG (eds) Aquaculture virology. Academic Press/Elsevier Inc, pp 425–439. https://doi.org/10.1016/B978-0-12-801573-5.00030-9
Tang KFJ, Lightner DV (2006) Infectious hypodermal and hematopoietic necrosis virus (IHHNV)-related sequences in the genome of the black tiger prawn Penaeus monodon from Africa and Australia. Virus Res 118:185–191. https://doi.org/10.1016/j.virusres.2006.01.003
Tang KFJ, Poulos BT, Wang J, Redman RM, Shih HH, Lightner DV (2003) Geographic variations among infectious hypodermal and hematopoietic necrosis virus (IHHNV) isolates and characteristics of their infection. Dis Aquat Org 53:91–99. https://doi.org/10.3354/dao053091
Tang KFJ, Pantoja CR, Poulos BT, Redman RM, Lightner DV (2005) In situhybridization demonstrates that Litopenaeus vannamei, L. stylirostris and Penaeus monodon are susceptible to experimental infection with infectious myonecrosis virus (IMNV). Dis Aquat Org 63:261–265. https://doi.org/10.3354/dao063261
Tang KFJ, Navarro SA, Lightner DV (2007) A PCR assay for discriminating between infectious hypodermal and hematopoietic necrosis virus (IHHNV) and virus-related sequences in the genome of Penaeus monodon. Dis Aquat Org 74:165–170. https://doi.org/10.3354/dao074165
Tang J, Ochoa WF, Sinkovits RS, Poulos BT, Ghabrial SA, Lightner DV, Baker TS, Nibert ML (2008) Infectious myonecrosis virus has a totivirus-like, 120-subunit capsid, but with fiber complexes at the fivefold axes. Proc Natl Acad Sci U S A 105:17526–17531. https://doi.org/10.1073/pnas.0806724105
Theilmann DA, Blissard GW, Bonning B, Jehle J, O’Reilly DR, Rohrmann GF, Thiem S, Vlak JM (2005) Family Baculoviridae. In: Fauquet CM, Mayo MA, Maniloff J, Desselberger U, Ball LA (eds) Virus taxonomy, eighth report of the International Committee on Taxonomy of Viruses. Elsevier, Amsterdam, pp 177–185
Thitamadee S, Prachumwat A, Srisala J, Jaroenlak P, Salachan PV, Sritunyalucksana K, Flegel TW, Itsathitphaisarn O (2016) Review of current disease threats for cultivated penaeid shrimp in Asia. Aquaculture 452:69–87
Venegas CA, Nonaka L, Mushiake K, Shimizu K, Nishizawa T, Muroga K (1999) Pathogenicity of penaeid rod-shaped DNA virus (PRDV) to kuruma prawn in different developmental stages. Fish Pathol 34:19–23
Vijayan KK, Alavandi SV, Rajendran KV, Alagarswami K (1995a) Prevalence and histopathology of monodon baculovirus (MBV) infection in Penaeus monodon and P. indicus in shrimp farms in the south-east coast of India. Asian Fish Sci 8:267–272 http://eprints.cmfri.org.in/id/eprint/6834
Vijayan KK, Alavandi SV, Rajendran KV, Alagarswami K (1995b) Prevalence and histopathology of monodon baculovirus (MBV) infection in Penaeus monodon and P. indicus in shrimp farms in the south-east coast of India. Asian Fish Sci 8:267–272
Volkman LE, Blissard GW, Friesen P, Keddie BA, Possee R, Theilmann DA (1995) Baculoviridae. In: Murphy FA, Fauquet CM, Bishop DHL, Ghabrial SA, Jarvis AW, Martelli GP, Mayo MA, Summers MD (eds) Virus taxonomy, sixth report of the International Committee on Taxonomy of Viruses. Springer, New York, pp 104–113
Walker PJ, Cowley JA, Spann KM, Hodgson RAJ, Hall MR, Withyachumnarnkul B (2001) Yellow head complex viruses: transmission cycles and topographical distribution in the Asia-Pacific region. In: Browdy CL, Jory DE (eds) The new wave, proceedings of the special session on sustainable shrimp culture, aquaculture 2001. World Aquaculture Society, Baton Rouge, pp 292–302
Wang CH, Lo CF, Leu JH, Chou CM, Yeh PY, Chuo HY, Tung MC, Chang CF, Su MS, Kou GH (1995) Purification and genomic analysis of baculovirus associated with white spot syndrome (WSBV) of Penaeus monodon. Dis Aquat Org 23:239–242. https://doi.org/10.3354/dao023239
Wang CS, Tang KFJ, Kou GH, Chen SN (1997) Light and electron microscopic evidence of white spot disease in the giant tiger shrimp, Penaeus monodon(Fabricius), and the kuruma shrimp, Penaeus japonicus (Bate), cultured in Taiwan. J Fish Dis 20:323–331. https://doi.org/10.1046/j.1365-2761.1997.00301.x
Wang CS, Chang JS, Wen CM, Shih HH, Chen SN (2008) Macrobrachium rosenbergii noda virus infection in M. rosenbergii (de Man) with white tail disease cultured in Taiwan. J Fish Dis 31:415–422. https://doi.org/10.1111/j.1365-2761.2008.00921.x
Wijegoonawardane PKM, Cowley JA, Phan T, Hodgson RAJ, Nielsen L, Kiatpathomchai W, Walker PJ (2008) Genetic diversity in the yellow head nidovirus complex. Virology 380:213–225. https://doi.org/10.1016/j.virol.2008.07.005
Wongteerasupaya C, Sriurairatana S, Vickers JE, Akrajamorn A, Boonsaeng V, Panyim S, Tassanakajon A, Withyachumnarnkul B, Flegel TW (1995a) Yellow-head virus of Penaeus monodon is an RNA virus. Dis Aquat Org 22:45–50. https://doi.org/10.3354/dao022045
Wongteerasupaya C, Vickers JE, Sriurairatana S, Nash GL, Akarajamorn A, Boonsaeng V, Panyim S, Tassanakajon A, Withyachumnarnkul B, Flegel TW (1995b) A non-occluded, systemic baculovirus that occurs in the cells of ectodermal and mesodermal origin and causes high mortality in the black tiger prawn, Penaeus monodon. Dis Aquat Org 21:69–77. https://doi.org/10.3354/dao021069
Xing H (2004) Discussion of the control measures for the “bottom death” (covert mortality disease) of Pacific white shrimp. China Fish 4:88–89 (In Chinese)
Yoganandhan K, Leartvibhas M, Sriwongpuk S, Limsuwan C (2006) White tail disease of the giant freshwater prawn Macrobrachium rosenbergii in Thailand. Dis Aquat Org 69:255–258. https://doi.org/10.3354/dao069255
Zhang QH (2004) To be cautious of bottom death in the intensive farming of Pacific white shrimp. Chin J Sci Fish Farming 10:48–49 (In Chinese)
Zhang J, Sun X (1997) A study on pathogens of Chinese prawn (Penaeus chinensis) virus diseases in the Fourth Asian Fisheries Forum ed. Zhou Yingqi, Zhou Hongqi, Yao Chaoqi, Lu Yi, Hu Fuyuan, Cui He and Din Fuhui Ocean, Bejing
Zhang QL, Liu Q, Liu S, Yang H, Liu S, Zhu L, Yang B, Jin J, Ding L, Wang X, Liang Y, Wang Q, Huang J (2014) A new nodavirus is associated with covert mortality disease of shrimp. J Gen Virol 95:2700–2709. https://doi.org/10.1099/vir.0.070078-0
Zhang QL, Xu TT, Wan XY, Liu S, Wang XH, Li XP, Dong X, Yang B, Huang J (2017) Prevalence and distribution of covert mortality nodavirus (CMNV) in cultured crustacean. Virus Res 233:113–119. https://doi.org/10.1016/j.virusres.2017.03.013
Acknowledgements
The authors are thankful to the National Surveillance Programme for Aquatic Animal Diseases, National Bureau of Fish Genetic Resources (NBFGR) and the authorities of Rajiv Gandhi Centre for Aquaculture for their support and encouragement.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Arulmoorthy, M.P., Anandajothi, E., Vasudevan, S. et al. Major viral diseases in culturable penaeid shrimps: a review. Aquacult Int 28, 1939–1967 (2020). https://doi.org/10.1007/s10499-020-00568-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-020-00568-3