
Abstract
In this study, Astronotus ocellatus were fed with food supplemented with Dunaliella salina (a natural β-carotene source) or astaxanthin (synthetic pigment), and their effects on fish skin carotenoids, growth indices and immune responses were evaluated. 135 A. ocellatus, weighing 25.6 ± 0.6 g, were randomly divided into three groups with three replicates (15 fish in each replicate). Group 1 and 2 were fed with a diet supplemented with 200 mg/kg astaxanthin or D. salina, respectively. Control group received the same diet without supplemented carotenoids. After 50 days, the growth indices were compared with the groups. Blood, mucus and skin samples were taken from each group. The immunological parameters of dietary D. salina and astaxanthin were studied in terms of serum and mucus lysozyme and bactericidal activity, as well as resistance against Aeromonas hydrophila infection. Carotenoid content of skin was assayed as well. Results showed that growth indices increased significantly in fish fed diet supplemented either with astaxanthin or D. salina (P < 0.05). Serum and mucus lysozyme and bactericidal activity increased in D. salina and astaxanthin groups (P < 0.05). Skin carotenoid content was statistically higher in astaxanthin group (6.48 ± 0.84 mg g−1) and D. salina group (4.89 ± 0.83 mg g−1) compared with the control group (3.05 ± 0.32 mg g−1) (P < 0.05). Rate of mortality following the challenge with A. hydrophila was significantly lower in D. salina group (50 ± 10) and astaxanthin (56.7 ± 5.8) group compared with control group (76.7 ± 5.8) (P < 0.05). Conclusively, D. salina and astaxanthin as a food additive can affect positively the growth and immunological parameters as well as skin carotenoid of A. ocellatus.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
As fish cannot synthesize their own pigments de novo, the pigments, which are synthesized by plants, algae and microorganisms, need to be incorporated in their diet. One of the greatest challenges in the ornamental fish industry is to replicate the accurate natural colour of the fish in the captive environment. In addition to affecting the skin colour of fish, these pigments also have other positive influence on strengthening the immune system and stimulating the growth indices of fish (Tanaka et al. 1976; Tacon 1981). There are four main pigment groups that give colour to the skin and tissues of animals and plants, namely melanines, purines, pteridiums and carotenoids. Carotenoids, which dissolve in fat, give the skin the yellow and red colours. They also give the orange and green colours to the egg, skin and flesh of many fish (Fuji 1969). Carotenoids, which are produced primarily by phytoplankton and plants, are divided into two groups as carotenes and xanthophylls. Although more than 600 carotenoids in nature have been defined, only a few of them are used in animal feeding, pharmaceuticals, cosmetics and food colouring (Bricaud et al. 1998, 1998). Carotenoids, particularly those that are vitamin A precursors, have received increasing attention due to their reported health benefits in mammals (Ong and Tee 1992; Krinski 1991). Microalgae, which are important in the production of larval fish because of their nutritive ingredients, can also be used as a natural pigment source in fish foods. The use of microalgal biomass has been recently investigated with regard to its potential as a colouring agent (Gourveia et al. 1997; Raymundo et al. 2005), but the use of synthetic pigment sources is more common because they are easy to obtain (Sales and Janssens 2003).
Astaxanthin (3,3′-dihydroxy-β,β-carotene-4,4′-dione) is a carotenoid from dietary origin that fish cannot synthesize. The wild fish derive astaxanthin through their prey organisms, while in aquaculture (aquariums and fish farms) astaxanthin is added to the feed in the form of nature identical synthetic beadlets (Bjerkeng et al. 1999). Although astaxanthin is an effective carotenoid, it is too expensive. The cost of astaxanthin supplementation represents about 10–20 % of the ingredient cost of fish feeds (Baker et al. 2002).
In some fish species such as ornamental fish and salmonids, skin and fillet colour constitutes an important quality parameter, and improving the fish colouration increases directly economical value of fish (Kop and Durmaz 2008). Since fish, like other animals, is unable to synthesize carotenoids de novo (Tacon 1981), skin colour is highly dependent on the carotenoids present in the diet.
Dunaliella salina is a type of halophile green microalgae especially found in hypersaline waters. It is well known for its antioxidant activity and large amount of carotenoids (Borowitzka and Borowitzka 1988). Dunaliella spp. are grown as a food source in aquaculture, and D. salina is the richest algal source of β-carotene and glycerol (Ben-Amotz and Avron 1983; Raja et al. 2007). D. salina can produce β-carotene up to 14 % of its dry weight under conditions of high salinity, light and temperature, even under nutrient limitation. D. salina as a natural source of β-carotene can improve skin and fillet pigmentation in fish.
In last decades, ornamental fish industry has developed and become more popular in Iran society. Astronorus ocellatus is a market-friendly and preferred ornamental fish in different countries especially in Iran. One of the most attractive features of this species is wide spectrum of body colour. Unfortunately, bacterial infection (especially motile Aeromonas infection) is very common health problem among this species in Iran, which leads the aquarists to use large amount of antibiotics and as a result, they are encountering bacterial resistance to antibiotics plus increase of production cost, so substances that either induce skin pigmentation or stimulate immune response and resistance against bacterial infections of this species can be the best choice as food additive.
Price and efficiency are indicators to choose the sources of pigment that can be used in fish feed. Although carotenoids are known substances to enhance immune function and disease resistance in higher animal (Bendich and Shapiro 1986; Jyounchi et al. 1993), little information is available for fish and no data about A. ocellatus. Therefore, in this study, effects of dietary astaxanthin and D. salina on skin carotenoids, growth performance, immune responses and disease resistance of A. ocellatus were evaluated.
Materials and methods
Fish
Albino Oscar fish (A. ocellatus) (weighing 25.6 ± 0.6 g), having similar skin colour, were purchase from an ornamental fish hatchery in suburb Ahvaz, Iran. Fish were transferred in oxygenated plastic bags to the wet laboratory of Shahid Chamran University, Ahvaz, Iran. Fish were fed with commercial granulated food (Biomar company, France), without any other supplementation for 1 month. After that, fish with similar skin colour were selected for the experimental study.
To acclimate and ascertain the constant colour of fish, impure fish were omitted, after 1-month period, in initial study.
Dunaliella salina preparation
Dunaliella sallina coded as Dunaliella M1 an isolate from Uremia Lake, Iran, was cultivated within a 14-l photobioreactor using the modified Junson medium described by Hejazi et al. (2010). NaCl concentration of the medium was 1.5 M. The photobioreactor was illuminated by Osram lamps (SPOT R80 NATURA E27/ESW) with the average light intensity of 400 μmol m−2 s−1. The temperature and pH were 26 ± 0.5 °C and 7.5 ± 0.3, respectively. After 7 days, stress condition was applied for over-production of β-carotene. During the stress stage, NaCl concentration of the medium and the average light were increased to 3 M and 2,000 μmol m−2 s−1, respectively. The cells were harvested after 8 days when the colour of the algae in photobioreactor turned to orange(Hejazi et al. 2010). D. sallina was harvested, lyophilized and stored in refrigerator until use. Then 200 mg/kg of alga mixed in a small amount of water and sprayed on commercial granulated fish food, and in order to prevent wasting of the alga in the water during feeding time, the surface of feed granules was coated with 5 % gelatin (Merk, Germany) then spread on a tray and air-dried in room temperature.
Astaxanthin (CAROPHYLL® 135 pink, DSM, Basel, Switzerland) at concentration of 200 mg/kg food was dissolved in distilled water and sprayed on the thin layer of basal food then air-dried in room temperature.
Sampling procedure
A total of 135 apparently healthy Oscar fish, which adapted to wet laboratory situation randomly distributed in three equal groups, in triplicate, following a completely randomized design (CRD) Group 1 and 2, were fed with a diet supplemented with 200 mg/kg astaxanthin (CAROPHYLL® pink, DSM, Basel, Switzerland) and D. sallina, respectively. Control group received the same food without supplementation. Fish sex was not taken into consideration because sex affects Oscar fish colour just after sexual maturation, and experimental fish were immature. Feeding trial was conducted for 50 days. The fish of each replicate weighed in day 0 and day 50, at the end of experiment. Fish were anaesthetized with 100 ppm MS222, and blood samples were taken randomly from six fish of each replicate of experimental groups. Sera samples were separated via centrifugation at 3,000g for 10 min and stored at −20 °C until used. Skin sample (around 0.5 g) of lateral flank near the dorsal fin was taken from each fish after taking the blood and euthanasia of fish with 1,000 ppm MS222 (Choubert and Storebakken, 1989).
Mucus sample preparation
After anaesthesia with 100 ppm MS222, the fish were placed with the ventral side of the body facing downward and cutaneous mucus from the dorsal side of the fish was collected by a cell scraper and transferred to a tube. Samples were kept on ice during transportation to the laboratory and kept frozen to avoid bacterial growth and degradation at −80 °C until used. The mucus was centrifuged (3,000 rpm for 10 min), and supernatant was used to analyse the lysozyme activity and antibacterial activity of mucus.
Lysozyme and bactericidal activity
Serum and mucus lysozyme activities were measured as described by Ellis (1990). Briefly, 10 μl of serum/mucus was mixed with 200 μl of a lyophilized Micrococcus lisodeichticus (Sigma) suspension at 0.2 mg ml−1 in 0.05 M sodium phosphate buffer (pH 6.2). The mixture was incubated at 27 °C, and its optical density (OD) was detected after 1 and 6 min at 530 nm using an enzyme-linked immunosorbent assay (ELISA) plate reader (Dynatech, Netherland). One unit of lysozyme activity was defined as the amount of enzyme that produced a decrease in absorbance of 0.001 min ml−1 serum.
The method used for serum bactericidal activity was followed a modified version of that adopted by Kajita et al. (1990). Briefly, the serum samples were diluted three times with 0.1 % gelatin-Veronal buffer, and mucus samples were used without dilution. Aeromonas hydrophila (live washed cells) were suspended in the same buffer, and bacteria were mixed at a ratio of 1:1 and incubated for 60 min at 25 °C and continuously agitated. The number of viable bacteria was then calculated by counting the resultant colonies from the incubated mixture on tryptic soy agar after incubation for 24 h in duplicate.
Growth performance indices
The growth indices including weight gain percentage (WGP), specific growth rate (SGR), feed conversion ratio (FCR) and protein efficiency ratio (FER) were calculated according to the following equations:
Total carotenoids
Skin total carotenoids of five fish in each replicate were determined at the end of the experiment by using spectrophotometer (Choubert and Storebakken 1989). After that, 10 mg of dry skin sample was homogenized with the addition of 5 ml acetone (98 %, Merck Germany) and centrifuged for 10 min at 3,500 rpm.Footnote 1 Supernatant of the samples was read at 475 nm wavelength on the spectrophotometer (JENWAY 6305).
Challenge study with A. hydrophila
At the end of experiment (day 50) of feeding, 0.1 ml of pathogenic isolates of A. hydrophila in sterile PBS (2 × 107 CFU ml−1) were intraperitoneally injected to ten fish per replicate. Mortality was recorded for 10 days. Tissues were taken from the dead fish for bacteriological culture to confirm A. hydrophila as the cause of death (Schaperclaus et al. 1992).
Statistical analysis
Statistical analysis consisted of one-way ANOVA, using the probability level of 0.05 for rejection of the null hypothesis. After ANOVA, significant differences between means were determined by Dancan’s multiple range test. All statistical analysis was performed using SPSS version 16.0.
Results
Growth performance
Significant differences were seen between astaxanthin- and D. salina-treated groups and control in all evaluated growth indices (P = 0.015). These data indicate a deep influence of carotenoids on growth performance. Although PER and FCR did not vary markedly between astaxanthin and D. salina groups (P = 0.0735), WGP and SGR showed significant difference between these two groups (P < 0.01) (Fig. 1).

Effect of astaxanthin and Dunaliella salina on growth performance of Astronorus ocellatus: Protein efficacy rate (PER), food conversion ratio (FCR), weight gain percentage (WGP) and specific growth rate (SGR) of A. ocellatus. N = 45 and values are mean ± SD
Lysozyme and bactericidal activity
The serum and mucus lysozyme activity was significantly increased with astaxanthin and D. salina groups when compared with the control group (Fig. 2). Serum and mucus Lysozyme activity between astaxanthin and Dunaliella did not show significant difference (P = 0.115).

Effect astaxanthin and Dunaliella sallina on Lysozyme activity of serum and mucus of Astronorus ocellatus: M1 and S1: mucus and serum samples of fish fed with free supplementation food. M2 and S2: mucus and serum samples of fish fed with 200 mg/kg D. salina supplemented food. M3 and S3: mucus and serum samples of fish fed with 200 mg/kg astaxanthin supplemented food. Different letters show significant difference (P < 0.05) between groups. N = 15 and values are mean ± SD
The number of the counted bacterial colonies was lower with D. salina group followed by astaxanthin group when compared with the control group (P < 0.05) as shown in Fig. 3. Serum bactericidal activity did not show markedly difference between astaxanthin and D. salina group (P = 0.0675), but Mucus bactericidal activity in D. salina group was significantly higher than astaxanthin group (P < 0.01).

Effect of astaxanthin and Dunaliella sallina on bactericidal activity of serum and mucus of Astronorus ocellatus: Legends are the same as on Fig. 2
Skin carotenoid content
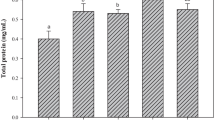
Supplementation of the food with D. salina and astaxanthin significantly affected the skin carotenoid rate of A. ocellatus (Fig. 4) (P = 0.023). Skin carotenoids were higher with astaxanthin group followed by D. salina group when compared with the control group.

Skin carotenoid content of A. ocellatus fed with basal diet (control) and 200 mg/kg D. salina or astaxanthin supplemented food for 7 weeks. Legends are the same as on Fig. 2
Disease resistance
The mortality was observed up to 10 days. The clinical signs observed in fish were swollen abdomen, reddish vent, exophthalmia and loss of equilibrium. Percentage cumulative mortality of A. ocellatus after challenging with A. hydrophila in different experimental groups showed that the highest mortality was recorded in control group which was significantly (P < 0.01) higher than D. salina- and astaxanthin-treated groups. Lowest mortality was observed in astaxanthin group (Fig. 5).

Cumulative mortality (%) of A. ocellatus following an experimental Aeromonas hydrophila infection, different letters show significant difference (P < 0.01) between groups. Values are mean ± SD
Discussion
Although no adverse effects of astaxanthin have been reported and that is permitted to be used in many countries, there is a great tendency for finding alternative similar materials, because astaxanthin derivates are expensive and add about an extra 10–15 % to the cost of feed. Microalgae (especially Dunaliella) are of the most favourite of these alternative materials both because of its nutritive quality and being a good source of carotenoid (Becker 1994). The two dietary supplements (astaxanthin and D. salina) affected growth indices in A. ocellatus. SGR, PER, FCR and WGP were promoted in fish fed with astaxanthin and D. salina supplemented diet compared with control. Nevejena et al. reported an increase in growth following administration of D. salina as a natural source of β- carotene in Argopecten purpuratus that is in agreement with our results. But Amar et al. (2004) and Choubert et al. (2006) reported no change in growth indices following administration of supplemented food with not only natural but synthetic carotenoid sources in rainbow trout.
Alishahi et al. (2014) reported similar growth stimulation effects of D. salina in Heros severus.
In fish, the effectiveness of carotenoid source in terms of deposition and physiological function is species-specific. In addition, all fish species do not possess the same pathways for the metabolism of carotenoids; therefore, there is no universal transformation of carotenoids in fish tissues (Kop and Durmaz 2008).
Natural β-carotene sources have been reported to act as a potent immunostimulants in fish (Raymundo et al. 2005; Supamattaya et al. 2005). In this study, some non-specific immune responses (i.e. serum and mucus Lysozyme activity, serum and mucus bactericidal activity and resistance against bacterial infection) were significantly increased following administration of astaxanthin and D. salina supplemented food. The lysozyme activity is an important indicator of the immune defence in both invertebrates and vertebrates (Ellis 1990). The lysozyme causes hydrolysis of the N-acetylmuramic acid and N-acetylglucosamine that are constituents of the peptidoglycan layer of bacterial cell wall and activation of the complement system and phagocytes by acting as an opsonin (Magnado’ttir 2006). Lysozyme is secreted by leucocytes and is an indicator of leucocyte activity, increasing concomitantly with phagocytic activity (Keshav et al. 1991). In this study, food supplemented with 200 mg/kg astaxanthin/D. salina significantly increased the serum and mucus lysozyme activity and stimulated the immune response in A. ocellatus. The increased lysozyme activity has been reported after supplementing the fish feed, with astaxanthin and D. salina in H. severus (Alishahi et al. 2014) rainbow trout (Amar et al.2004), Aloe vera extract (Alishahi et al. 2010) and injection of vaccines (Alishahi and Buchmann 2006).
β-carotene is a vitamin A precursor and a quencher of singlet oxygen; thus, it could affect the immune system through both the antioxidant and retinoid pathways (Krinski, 1991). D. salina also contains large amounts of 9-cis β-carotene, which has been shown to be more potent than all-trans β-carotene in enhancing cell–cell interaction through gap junctions (Zhang et al. 1992). Thus, the augmentation of lysozyme activity by these carotenoid sources could be via its stimulation of phagocytic cells as is the case with most immunostimulants (Sakai 1999). Astaxanthin is also a precursor of vitamin A in fish (Matsuno 1991) and possesses free radical scavenging ability.
Our results also showed that supplemented diet with astaxanthin and D. salina significantly increased the serum and mucus bactericidal activity against A. Hydrophila (P < 0.05). Alishahi et al. (2014) reported increased lysozyme activity following oral administration of 100 and 200 mg/kg D. salina in H. severus. Divyagnaneswari et al. (2007) in tilapia and Kajita et al. (1990) in rainbow trout reported an increase in serum bactericidal activity after administration of herbal extracts as immunostimulants.
It is important to estimate bacterial resistance of treated fish to determine the efficiency of an immunostimulant. In the present study, the lowest mortality after challenge was registered in D. salina group followed by astaxanthin group, which was significantly lower than the control group. The higher survival rate of D. salina group may be because of presence of other beneficial substances in algae structure that are not available in pure astaxanthin source.
The study showed that supplementation of astaxanthin and D. salina had a positive influence on the survival of A. ocellatus by resisting the A. hydrophila infection. Based on the results of the present study, it can be concluded that growth indices are affected by dietary supplementation of D. salina and astaxanthin. It appears that D. salina and astaxanthin are effective in enhancing some of the non-specific immune responses in A. ocellatus. Moreover, mostly lower humoral responses observed with natural sources in some parts of this study compared with synthetic sources may be due to the isomers and esterified forms of carotenoids found in the former which may have a lower bioavailability. Nevertheless, natural carotenoids from D. salina are promising and advantageous sources of dietary supplements for fish, not only for pigmentation purposes but also for the health benefits they provide.
Notes
Rounds per minute.
References
Alishahi M, Buchmann K (2006) Temperature-dependent protection against Ichthyophthirius multifiliis following immunisation of rainbow trout using live theronts. Dis Aquat Org 72:269–273
Alishahi M, Ranjbar M, Ghorbanpor M, Peyghan R, Mesbah M, Razijalali M (2010) Effects of dietary Aloe vera on specific and nonspecific immunity in the common carp (Cyprinus carpi). Int J Vet Res 4:189–195
Alishahi M, Karamifar M, Mesbah M, Zarei M (2014) Hemato-immunological responses of Heros severus fed diets supplemented with different levels of Dunaliella salina. Fish Physiol Biochem 40:57–65
Amar EC, Kirona V, Satoha S, Watanabea T (2004) Enhancement of innate immunity in rainbow trout (Oncorhynchus mykiss Walbaum) associated with dietary intake of carotenoids from natural products. Fish Shellfish Immunol 16:527–537
Baker RTM, Pfeiffer AM, Schöner FJ, Smith-Lemmon L (2002) Pigmenting efficacy of astaxanthin and canthaxanthin in fresh-water reared Atlantic salmon (Salmo salar). Anim Feed Sci Technol 99:97–106
Becker EW (1994) Microalgae biotechnology and microbiology. Cambridge University Press, Cambridge
Ben-Amotz A, Avron M (1983) On the factors which determine massive β-carotene accumulation in the halotolerant alga Dunaliella bardawil. Plant Physiol 72:593–597
Bendich, Shapiro SS (1986) Effect of β-carotene and canthaxanthin on the immune responses of the rat. J Nutr 116:2254–2262
Bjerkeng B, Hatlen B, Wathne E (1999) Deposition of Astaxanthin in fillets of Atlantic salmon (Salmo salar) fed diets with herring, capelin, sandeel, or Peruvian high PUFA oils. Aquaculture 180:307–319
Borowitzka MA, Borowitzka LJ (1988) Dunaliella. In: Borowitzka MA, Borowitzka LJ (eds) Microalgal Biotechnology. Cambridge University Press, New York
Bricaud A, Morel A, Babin M, Allali K, Claustre H (1998) Variations of light absorption by suspended particles with chlorophyll a concentration in oceanic (case 1) waters: analysis and implications for biooptical models. J Geophys Res 103:31033–31044
Choubert G, Storebakken T (1989) Dose response to astaxanthin and canthaxanthin pigmentation of rainbow trout fed various dietary carotenoids concentrations. Aquaculture 81:69–77
Choubert G, Mendes-Pinto MM, Morais R (2006) Pigmenting efficacy of astaxanthin fed to rainbow trout Oncorhynchus mykiss: effect of dietary astaxanthin and lipid sources. Aquaculture 257:429–436
Divyagnaneswari MD, Christybapita R, Dinakaran M (2007) Enhancement of nonspecific immunity and disease resistance in Oreochromis mossambicus by Solanum trilobatum leaf fractions. Fish Shellfish Immunol 23:249–259
Ellis AE (1990) Lysozyme assay. In: Stolen JS, Fletcher DP, Anderson BS, Robertson BS (eds) Techniques in fish immunology. SOS Publication, Fair Haven, New Jersey, pp 101–103
Fuji R (1969) Chromatophores and pigments. In: Hoar WS, Randall DJ (eds) Fish physiology. Reproduction and growth. Bio Luminescence, Pigments and Poisons. Academic Press, New York, pp 301–353
Gourveia L, Gomes E, Empis J (1997) Use of Chlorella vulgaris in diets for rainbow trout to enhance pigmentation of muscle. J Appl Aquat Cult 7:61–70
Hejazi M, Barzegari A, Hosseinzadeh A, Hejazi MS (2010) Introduction of a novel 18S rDNA gene arrangement along with distinct ITS region in the saline water microalga Dunaliella Saline Systems 6(4):11–17
Jyounchi H, Zhang L, Tomita Y, Gross M (1993) Immunomodulating actions of carotenoids-enhancement of Ab production against T-dependent Ag (TD-Ag) in vivo and in vitro. FASEB J 7:519–521
Kajita Y, Sakai M, Atsuta S, Kobayash M (1990) The immunomodulatory effects of levamisole on rainbow trout, Oncorhynchus mykiss. Fish Pathol 25:93–98
Keshav S, Chung P, Milton G, Gordon S (1991) Lysozyme is an inducible marker of macrophage activation in murine tissues as demonstrated by in situ hybridization. J Exp Med 174:1049–1058
Kop A, Durmaz Y (2008) The effect of synthetic and natural pigments on the colour of the cichlids (Cichlasoma severum). Aquaculture 16:117–122
Krinski N (1991) Effects of carotenoids in cellular and animal systems. Am J Clin Nutr 53:238–246
Magnado’ttir B (2006) Innate immunity of fish (overview). Fish Shellfish Immunol 20:137–151
Matsuno T (1991) Xanthophylls as precursors of retinoids. Pure Appl Chem 63:81–88
Ong ASH, Tee ES (1992) Natural sources of carotenoid from plants and oils. Meth Enzymol 213:142–167
Raja R, Hemaiswarya S, Rengasamy R (2007) Exploitation of Dunaliella for bcarotene production. Appl Microbiol Biotechnol 74:517–523
Raymundo A, Gouveida L, Batisa AP, Empis J, Sousa L (2005) Fat mimetic Capacity of chlorella vulgaris biomass in oil-in-water food emulsions stabilized by pea protein. Food Res lnt 38:961–965
Sakai M (1999) Current status of fish immunostimulants. Aquaculture 172:63–92
Sales J, Janssens PX (2003) Nutrient requirements of ornamental fish. Aquat Living Resour 16:533–540
Schaperclaus W, Kulow H, Schreckenbach K (1992) Fish Disease. A.A. Balkema, Rotterdam
Supamattaya K, Kiriratnikom S, Boonyaratpalin M (2005) Effect of a Dunaliella salina extract on growth performance, health condition, immune response and disease resistance in black tiger shrimp (Penaeus monodon). Aquaculture 248:207–216
Tacon AGJ (1981) Speculative review of possible carotenoid function in fish. Progess Fish Cult 43:205–208
Tanaka Y, Matsuguchi H, Katayama T, Simpson KL, Chichester CO (1976) The biosynthesis of astaxanthin XVIII. The metabolism of the carotenoids in the prawn, Penaeus japonicus Bates. Bull Jpn Soc Sci Fish 42:197–202
Zhang LX, Cooney RV, Bertram JS (1992) Carotenoids up-regulate Connexin 43 gene expression independent of their provitamin A or antioxidant properties. Cancer Res 52:5707–5712
Acknowledgments
This work was financially supported by the Research Council of Shahid Chamran University, Ahvaz, Iran.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Alishahi, M., Karamifar, M. & Mesbah, M. Effects of astaxanthin and Dunaliella salina on skin carotenoids, growth performance and immune response of Astronotus ocellatus . Aquacult Int 23, 1239–1248 (2015). https://doi.org/10.1007/s10499-015-9880-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-015-9880-0