
Abstract
Rainbow trout were fed a diet containing phytase-sprayed and phytase-pretreated soybean meal with different phytase levels. The single factor random block design was used to analyze the effects on rainbow trout of dietary phytase supplementation on growth performance, nutritional ingredient digestibility and nutrient excretion. After 90 days, the results showed that feed conversion ratio (FCR) and protein efficiency ratio (PER) were significantly improved and specific growth rate (SGR) was not affected by spraying phytase, but SGR, FCR and PER were not significantly improved by phytase pretreatment. A digestibility trial conducted after the feeding trial showed that apparent digestibility coefficient (ADC) of diet protein and minerals was increased with phytase supplementation. However, there was a negative effect of phytase on the ADC of lipid. The excretion experiment showed that the supplementation of phytase resulted in decreased nutrient excretion in feces, but lipid excretion was slightly increased with phytase supplementation. In addition, the results of P excretion and ADC of P analyzed by t-test showed that phytase pre-treatment method should be a more rational method than the spraying method. The results of SGR, ADC of P and P excretion analyzed by quadratic regression indicated that 2,000–3,000 U/kg levels by the spraying method could be a rational range of phytase supplementation, and about 1,000 U/kg should be an optimal level by the pretreatment method. Thus, use of phytase in rainbow trout feeds can have economic and environmental benefits.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Fish meal has been a large part of dietary protein source in feed industry. However, with the expansion of aquaculture increasing demand and high price has made it necessary to search for alternative protein sources. At present, replacing fish meal by plant protein sources in fish nutrition is of increasing interest (Fontainhas-Femandes et al. 1999; Mabahinzireki et al. 2001). In plant protein sources, soybean meal is used in animal feeds due to its well-balanced amino acid profile and relatively high crude protein level (Cheng and Hardy 2002). However, the major limitation in the use of alternative protein is related to anti-nutritional factors in soybean meal, such as protease inhibitors, non-starch polysaccharides and phytate (Francis et al. 2001). Phytate is relatively heat-stable and cannot be effectively removed without enzymatic reactions. Therefore, several mineral elements like Ca, Mg, Zn, Mn, Cu and Fe (Papatryphon et al. 1999) as well as feed protein (Liu et al. 1998; Sugiura et al. 2001) can be reduced in their bioavailability by phytate. Phytate is also a major P component in soybean meal, and is a typical example of a component affecting the availability of dietary elements. So phytate-P is not available to monogastric animals including fish without enzymatic reactions (National Research Council 1993). Sugiura et al. (1998) reported that 22% of the P in soybean meal was available for rainbow trout. Riche and Brown (1996) found that P in soybean meal was completely unavailable to rainbow trout. So P in aquaculture effluents is considered a point source of pollution by many regulatory agencies (Lall 1991). Uneaten food and unavailable dietary P in feces are the two primary contributors in fish farm P effluents (Bergheim et al. 1991). In intensive fish farming systems, a poor degradation of phytate leads to detrimental effects on the aquatic environment such as eutrophication effects on fresh water.
Microbial phytase is an enzyme used to specifically hydrolyze phytate and release the mineral elements chelated in plant protein source. It has been demonstrated that the addition of phytase in the diet has the potential to improve the digestibility of nutrients and minerals (Cheng and Hardy 2003; Yoo et al. 2005), and the feed conversion efficiency and growth (Sajjadi and Carter 2004; Schafer et al. 1995; Rodehutscord and Pfeffer 1995; Vielma et al. 1998), and to reduce P discharge into water (Jackson et al. 1996; Lanari et al. 1998; Papatryphon et al. 1999; Forster et al. 1999; Vielma et al. 2002; Sajjadi and Carter 2004). But some studies showed no effect on weight gain in fish (Lanari et al. 1998; Vielma et al. 2000; Yan et al. 2002). Several factors influence the efficacy of phytase, including the type and amount of phytase, supplementation method of phytase, concentrations of nutrients (especially protein and minerals), as well as the fish species (Sajjadi and Carter 2004). Results of these studies have shown that dietary nutrient retention is increased in fish fed phytase-supplemented diets, but effects of addition of phytase by different methods on nutrient excretion and the aquatic environment have not been well documented. This study was conducted to quantify the effects of dietary phytase at continuous levels with different methods on growth performance, apparent digestibility, nutrient excretion and the aquatic environment, and to quantify which phytase supplementation method (spraying and pretreatment) and what dose of phytase in each phytase supplementation method were better.
Materials and methods
Diets
Fish meal, soybean meal and corn gluten meal were used as protein sources in the basal diet. Menhaden fish oil and soybean oil were used as the lipid sources, and the basal diet was formulated to contain approximately 43.19% crude protein and 10.17% crude fat. The formulation and chemical composition of the basal diet is shown in Table 1. The data of phytate-P and phytase activity in test diets are shown in Table 2.
All the ingredients were ground into fine powder through a 320-μm mesh, and were thoroughly mixed with fish oil and soybean oil, then pelleted with an experimental feed, and dried for about 12 h in a ventilated oven at 40°C. 0.5% chromic oxide (Cr2O3) was added to each diet as an inert marker. The particle size of diet was Φ 1.5 × 3.0 mm. After drying, the diet with 8–10% moisture was stored at 4°C until feeding.
Microbial phytase was supplied by BASF Corporation. The actual enzymatic activity of phytase (5,280 U/g) was measured by the method of Slominski et al (2007). In spraying groups, phytase was added to the basal diet at the levels of 0, 500, 1,000, 1,500, 2,000, 2,500, and 3,000 U/kg diets, respectively. Phytase coating of basal diet followed recommendations of Jackson et al. (1996). The detail was as follows:
-
(1)
50 ml of water containing citrate buffer at pH 5.0 was prepared at room temperature.
-
(2)
The proper amount of phytase was dissolved in the buffer.
-
(3)
The buffer was sprayed on 1 kg of the finished diet as needed.
In pretreatment group, phytase was added to the basal diet at the levels of 0, 500, 1,000, 1,500 U/kg diets, respectively. Phytase pretreatment of soybean meal followed recommendations of Cain and Garling (1995). The detail was as follows:
-
(1)
One liter of water containing citrate buffer at pH 5.0 was prepared at room temperature.
-
(2)
The proper amount of phytase was dissolved in the buffer.
-
(3)
One kilogram of soybean meal was added to the buffer and the mixture was rapidly heated, with constant stirring, to 50–55°C. The mixture was held at this temperature for 5–6 h. Dry matter content was 40–50% after the heating period.
-
(4)
Once the heating process was complete, treated soybean meal was cooled and dried at room temperature in a forced-air drying oven for 24 h to reduce the moisture content to approximately 10%.
-
(5)
Dried, phytase-pretreated soybean meal was added to the experimental feeds.
Experimental units and feeding
The experiment was carried out at the Fisheries Research Institute; Harbin Academy of Agricultural Sciences (Heilongjiang, China). The experiment started in July 2006 and lasted for 90 days. Each diet was fed to rainbow trout in three 150-l tanks. Each tank was stocked with 30 fish with an initial average weight of 1.70 ± 0.03 g. The photoperiod was set at 12-h light and 12-h dark. Freshwater temperature was 14–18°C. The O2-content of the outlet water remained above 8 mg/l. Water flow was similar in all tanks (0.5 l/min).
The rainbow trout were hand-fed twice (0800 and 1400 hours) per day and the amount of feed distributed was adjusted weekly according to the increased body weight. Fish were fed about 2.5% of body weight. Each tank was equipped to collect rejected feed from the water outlet, as described by Helland et al. (1996). The feces were collected at 2–4 h after feeding. Then the feces were freeze dried and ground with a pulverizer to analyze for subsequent determinations of protein, fat, P, Ca and other nutrient contents.
Chemical analyses
The dry matter content of diet and feces was determined by drying at 105°C until stable weight, and ash was determined after combustion at 550°C until stable weight. Crude protein was determined as Kjeldahl-N × 6.25 on a KJELTE 2300 auto-analyzer (FOSS-2300, USA). Crude fat was determined by ether extraction using Soxhlet. Total-P and phytate-P in samples were determined in duplicate using the vanadium-molybdate method (total-P) and ammonium-molybdate method (phytate-P), respectively (AOAC 1990). Ca, Mg, Fe, Zn, Mn and Cu were determined by flame atomic absorption spectrophotometer (TAS-990, China) after wet ashing and acid digestion. Cr2O3 concentrations were determined by flame atomic absorption spectrophotometer following combustion of the sample in a muffle furnace, before and after digestion in nitric acid (AOAC 1995).
Calculations and statistical analysis
The following variables were calculated:
-
Specific growth rate (SGR) = (LnW t − LnW 0) × 100/t
-
Feed conversion ratio (FCR) = feed consumed (g, DW)/weight gain (g)
-
Protein efficiency ratio (PER) = weight gain (g)/protein intake (g)
-
Nutrient excretion per gain (g/kg weight gain) = FCR × nutrient contents in diet × (1 − ADC of nutrient) × 1,000
-
Nutrient apparent digestibility coefficient (ADC) = 100 × \( \left[ {1 - \left( {\frac{{F\,\times\,D_{Cr} }}{{D\,\times\,F_{Cr} }}} \right)} \right] \)
where W t and W 0 are final and initial body mass, respectively, t is the experimental duration days. F and D represent nutrient contents in faeces and diet, respectively. F Cr and D Cr represent Cr2O3 contents in feces and diet, respectively. All data were subjected to one-way ANOVA after homogeneity in variance was tested using SAS for Windows. When a significant treatment effect was observed, Duncan’s multiple range test was used to compare means. A quadratic regression analysis method (Snedecor and Concbran 1978) was conducted to analyze SGR, P excretion and the ADC of P of the rainbow trout (Figs. 1–3). And these data in spraying and pretreatment groups (500–1,500 U/kg) were compared using a two-tailed t-test (Table 3). The results were presented as means ± SEM. A significant level of P < 0.05 was employed in all cases.

The relationship between SGR and phytase level in rainbow trout
Results
Growth
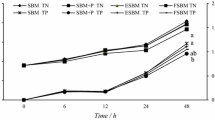
The growth data are shown in Table 4. In spraying groups, fish fed the diet with supplementation of phytase showed slightly higher SGR than fish fed the basal diet, but SGR did not differ significantly among treatment groups (P > 0.05). Quadratic regression analysis indicated that a SGR optimum occurred at the level of 2,145 U/kg diet phytase (Fig. 1). The FCR and PER were improved significantly by phytase supplementation (≥1,000 U/kg), but FCR and PER did not differ among fish fed ≥500 U/kg. In pretreatment groups, SGR, FCR and PER were not affected significantly by phytase (P > 0.05). Quadratic regression analysis indicated that a SGR optimum occurred at the level of 1,194 U/kg diet phytase (Fig. 1).
Apparent digestibility
The ADC of nutrient is shown in Table 5. In spraying groups, the ADC of P was significantly influenced by supplementation phytase (P < 0.05). Quadratic regression analysis indicated that ADC of P optimum occurred at the level of 3,236 U/kg diet phytase (Fig. 2). Fish fed phytase supplements had higher ADC of protein and Ca than fish fed unsupplemented diet (P < 0.05). The ADC of protein was 94.61–95.73% for diets containing ≥500 U/kg and no significant difference was observed (P > 0.05). The Ca digestibility was 20–40% higher for fish fed the diet with supplementation phytase than for fish fed the basal diet. There was also no significant difference among the 500–3,000 U/kg groups (P > 0.05). However, there was a negative effect of phytase on the ADC of lipid. The ADC of lipid was slightly reduced from 89.58% to 84.92% of the diet with supplementation phytase (P > 0.5). In pretreatment groups, the ADC of P was improved significantly by supplementation phytase (P < 0.05), but there was no significant difference among the 500–1,500 U/kg groups (P > 0.05).The highest ADC of P was observed in fish fed diet with 1,000 U/kg phytase, and quadratic regression analysis indicated that a ADC of P optimum occurred at the 950 U/kg level (Fig. 2). Similarly, the ADC of protein had a similar trend with ADC of P, but ADC of lipid and Ca was not affected significantly by phytase (P > 0.05).

The relationship between the ADC of P and phytase level in rainbow trout
In the spraying groups, the diet with phytase supplementation significantly increased the bioavailability P in feed ingredients and increased utilization of naturally occurring minerals in feed ingredients. The ADC of Cu, Fe, Zn, Mn and Mg were improved significantly for the diet with supplementation phytase compared with the basal diet (P < 0.05). The highest ADC of Cu, Fe, Zn, Mn and Mg were observed in the 1,500, 1,000, 2,500, 2,500, 1,500 U/kg groups, respectively. In pretreatment groups, the ADC of Cu, Zn, Mn, Mg were improved significantly for the diet with 1,000 U/kg phytase compared with the basal diet (P < 0.05), but the ADC of these minerals had no significant difference among fish fed ≥500 U/kg (P > 0.05).
Nutrient excretion
The nutrient excretion data were shown in Table 6. In spraying groups, P excretion was significantly decreased for fish fed diet with supplementation phytase (P < 0.05), and quadratic regression analysis indicated that 2,795 U/kg diet phytase was an optimum level (Fig. 3). Fish fed diet with supplementation phytase significantly decreased the excretion of Ca, protein and other minerals (P < 0.05). However, lipid excretion was slightly increased and it was not affected by phytase (P > 0.05). In pretreatment groups, there was the similar trend within spraying groups. Quadratic regression analysis indicated that P excretion optimum occurring at the level of 1,065 U/kg (Fig. 3). The excretion of Ca and Fe were not affected significantly by phytase (P > 0.05). Furthermore, the other nutrients excretion did not differ significantly among fish fed ≥500 U/kg (P > 0.05).

The relationship between P excretion and phytase level in rainbow trout
Discussion
Growth
Previous study reported that phytate binds trypsin in vitro thus reducing protein digestibility (Singh and Krikorian 1982). Spinelli et al. (1983) also found that phytate decrease protein digestibility, growth rate and feed efficiency in rainbow trout. The effect of phytic acid on growth depends primarily on the amount in the diet (Sajjadi and Carter 2004). In the present study, SGR was slightly improved for fish fed with phytase. FCR was lower than 1.0 and PER was improved for fish fed with phytase. This means that the phytase used in degrading the phytate resulted in the release of more minerals and increased minerals utilization in growth performance of the fish. Growth improvement demonstrates that soybean can replace most of the fish meal in the diet by supplementation phytase for rainbow trout. Similarly, growth improvements were also observed in some studies on dietary supplementation phytase (Rodehutscord and Pfeffer 1995; Schafer et al. 1995; Papatryphon et al. 1999; Cain and Garling 1995; Vielma et al. 2002). Conversely, the inclusion of phytase did not improve growth in other studies (Lanari et al. 1998; Forster et al. 1999; Vielma et al. 2000). Obviously, further research is needed to clarify the influence of supplemental phytase on protein availability, growth performance and feed utilization of fish.
Apparent digestibility coefficient
Singh and Krikorian (1982) reported that phytate binds trypsin in vitro thus reducing the bioavailability of various dietary components and fish performance. Phytate can also chelate with protein to form phytate–protein or phytate–mineral–protein complexes that are resistant to proteolytic digestion (Cheryan 1980). Additionally, phytate may bind to amino acids in the fish stomach and decrease amino acid availability.
Riche and Brown (1996) and Sugiura et al. (1998) reported that the apparent availability of P in soybean meals was 0% and 22% for rainbow trout, respectively. And about 60% P in soybean meal is in the form of phytate (Sajjadi and Carter 2004) which fish cannot digest. In the present study, the phytate-P level of soybeans diets was 57.72% of the total-P, and this was the reason why the bioavailability of P was lower in the control group. Addition of phytase can improve ADC of P in diet and then it is possible to improve P retention and reduce P discharge into water. Phytase supplementation in diet increased P digestibility by rainbow trout, similar to other studies with Atlantic salmon (Storebakken et al. 1998) and other salmon (Brown 1993; Cain and Garling 1995; Rodehutscord and Pfeffer 1995; Teskeredzic et al. 1995; Riche and Brown 1996; Lanari et al. 1998; Vielma et al. 1998; Forster et al. 1999; Sugiura et al. 2001). However, in this study, the high levels of phytase slightly reduced ADC of P. This is because dietary P absorption is regulated by blood P level (Lall 1991), and hence, when the blood P level is saturated or dietary P concentration increases above the requirements, the utilization efficacy of retention decreases (Rodehutscord 1996; Jahan et al. 2003).
In the present study, the ADC of protein, Ca, P, and minerals were improved for diets with supplementation phytase. The effect on protein digestibility for fish fed dietary phytase agreed with earlier observations, e.g., with Vielma et al. (2004). But conversely, some studies on rainbow trout (Vielma et al. 2000; Cheng and Hardy 2002) and Atlantic salmon (Sajjadi and Carter 2004; Lanari et al. 1998) reported that a lack of effect on protein digestibility was observed for fish fed a diet with supplementation phytase. The discrepancy reported for protein digestibility in several studies might be related to differences in some other factors, such as protein feedstuff quality, drying procedures and gastrointestinal pH. In the present study, ADC of lipid was slightly reduced by dietary phytase. Adding phytase may inhibit the activity of lipase and decrease the efficiency of lipase hydrolysis lipid, thus ADC of lipid was reduced with supplementation phytase. However, why phytase inhibited the activity of lipase is not clear, and further research is needed to clarify it.
The ability of phytate to bind divalent minerals is well known. Wise (1983) pointed out that phytate in plant meals could form a phytate–mineral complex and bind to other divalent and trivalent canons such as Ca, Cu, Fe, Mg, Mn and Zn. Lall (2002) reported that growth reduction may be related to deficiency of several minerals. In the present study, the ADC of minerals in the control group was the lowest along with the lowest weight gain. When ADC of P was increased, the ADC of Mg, Mn, Fe, Cu and Zn were also increased. The function of dietary phytase to release P and other minerals for rainbow trout and other fish has been reported by other studies (Cain and Garling 1995; Rodehutscord and Pfeffer 1995; Schaefer et al. 1995; Jackson et al. 1996; Lanari et al. 1998; Oliva-Teles et al. 1998; Powers Hughes and Soaves 1998; Storebakken et al. 1998; Vielma et al. 1998, 2000; Forster et al. 1999; Papatryphon et al. 1999; Masumoto et al. 2001; Papariyphon and Soaves 2001; Sugiura et al. 2001).
Nutrient excretion
P is one of the most important pollution sources in the freshwater environment. Therefore, reducing dietary P levels and fecal P discharges are critical approaches to reducing environmental pollution. Because phytate-bound P is not available, unavailable dietary P in feces is the primary contributor in fish farm effluents. In the present study, P excretion of 3.81 g/kg rainbow trout was produced by the fish fed the basal diet. The P excretion decreased from 3.81 to 1.46 g/kg by spraying phytase and from 2.78 to 0.62 g/kg by phytase pretreatment. This indicates that more degradation of phytate occurred as other components of diet were digested and absorbed. Therefore, P excretion was reduced with dietary phytase. Furthermore, protein excretion of fish fed dietary phytase was decreased significantly. Meanwhile, mineral excretion of fish fed un-supplemented diet was significantly higher than of fish fed with dietary phytase. The reduced nitrogen and mineral excretion with phytase supplementation in our study validated the fact that the protein and mineral utilization were improved by phytase supplementation. According to Storebakken et al. (1998), fish fed diet containing phytase pre-treated soybean released significantly less amount of fecal P into the environment than those fed either fish meal or soy concentrate diets. Sajjadi and Carter (2004) also reported that phytase incorporation in diets containing high levels of plant protein substantially increased P digestibility and reduced P leaching from feces. Thus, the application of phytase to fish diets is significant.
Phytase supplementation method
As we know, many factors interactively influence the efficacy of phytase, which include pH, temperature, ratios of calcium to P and the supplementation phytase method. In our study, we chose two supplementation phytase methods (phytase spraying and phytase pretreatment). In spraying groups, phytate-P concentration of diet with supplementation phytase was slightly lower than that of the basal diet with no supplementation phytase, but that was nearly identical among the 500–3,000 U/kg groups (Table 2). It indicated that phytase spraying on feed pellets cannot hydrolyze phytate efficiently in vitro. In the pretreatment groups, the phytate-P concentration of diet with supplementation phytase was decreased greatly compared with that of the basal diet, which was reduced 68.3%, 92.7% and 90.2% in diets with supplementation phytase 500–1,500 U/kg, respectively. Phytate-P was nearly completely hydrolyzed by phytase in vitro when diet was pretreated ≥1,000 U/kg phytase.
The effects of phytase spraying onto diet pellets have been studied in many aquaculture species (Vielma et al. 1998; Schaefer and Koppe 1995; Jackson and Robinson 1996), as a current method to use liquid enzyme coating pellets in feed processing. However, phytase pretreatment feed ingredients has been studied in some species, such as Atlantic salmon (Storebakken et al. 1998; Teskeredzic et al. 1995) and rainbow trout (Cain and Garling 1995; Vielma et al. 2002). In the present study, the results of SGR, P excretion and ADC of P analyzed by t-test showed that effects of phytase with different addition methods on P excretion (P = 0.0030) and ADC of P (P = 0.0008) differed significantly, but the effects of phytase with different methods on SGR (P = 0.5347) showed no significant difference (Table 3). Although the pre-treatment method could not improve growth performance significantly compared with spraying, pre-treatment can increase bioavailability of P and reduce P excretion efficiently at the same phytase level. So phytase pre-treatment of soybean meal might be a more rational method than conventional phytase addition to the formula diet of rainbow trout.
In addition, the results of SGR, ADC of P and P excretion analyzed by quadratic regression indicated that 2,000–3,000 U/kg levels by the spraying method and about 1,000 U/kg level by the pretreatment method could be a rational range of phytase supplementation. In this study, the optimum level of spraying addition phytase was higher than in several other studies (Schaefer et al. 1995; Cheng and Hardy 2002). The discrepancy might be related to differences in phytase source, diet formulation, fish species and water environment.
Conclusion
Although, this fecal collection method may have some minor influence on the ADC of nutrients, the experimental results still indicated that phytase addition could improve mineral utilization in rainbow trout. In this study, phytase pretreatment might be a better method than phytase spraying to increase mineral digestibility and to decrease mineral excretion of rainbow trout at the same phytase level. For better protection of the environment, when adding phytase the approximate level should be 3,000 U/kg by the spraying method, but for the expenditure of feed production, about 2,000 U/kg was sufficient. However, about 1,000 U/kg diet phytase should be an optimal level by the pretreatment method.
Abbreviations
- ANOVA:
-
Analysis of variance
- ADC:
-
Apparent digestibility coefficient
- SGR:
-
Specific growth rate
- FCR:
-
Feed conversion ratio
- PER:
-
Protein efficiency ratio
References
AOAC (1990) Official methods of analysis of Official Analytical Chemists International, 15th edn. Association of Official Analytical Chemists, Arlington
AOAC (1995) Official methods of analysis of Official Analytical Chemists International, 16th edn. Association of Official Analytical Chemists, Arlington
Bergheim A, Aabel JP, Seymour EA (1991) Past and present approaches to aquaculture waste management in Norwegian net pen culture operations. In: Cowey CB, Cho CY (eds) Nutritional strategies and aquaculture waste. Proceedings of the first international symposium on nutritional strategies in management of aquaculture waste, 2–6 June 1990, Guelph, Canada. University of Guelph, Ontario, Canada, pp 117–136
Brown PB (1993) Comparison of fecal collection methods for determining P absorption in rainbow trout. In: Kaushik SJ, Luquet P (eds) Fish nutrition in practice, Biarritz (France), June 24–27, 1991. Ed. INRA, Paris 1993 (Les Colloques, no. 61), pp 443–447
Cain KD, Garling DL (1995) Pretreatment of soybean meal with phytase for salmonid diets to reduce P concentrations in hatchery effluents. Prog Fish-Cult 57:114–119
Cheng ZJ, Hardy RW (2002) Effect of microbial phytase on apparent nutrient digestibility of barley, canola meal, wheat and wheat middlings, measured in vivo using rainbow trout (Oncorhynchus mykiss). Aquac Nutr 8:271–277
Cheng ZJ, Hardy RW (2003) Effects of extrusion and expelling processing, and microbial phytase supplementation on apparent digestibility coefficients of nutrients in full-fat soybeans for rainbow trout (Oncorhynchus mykiss). Aquaculture 218:501–514
Cheryan M (1980) Phytic acid interactions in food systems. Crit Rev Food Sci Nutr 13:297–335
Fontainhas-Femandes A, Gomes E, Reis-Henriques MA, Coimbra J (1999) Replacement of fish meal by plant protein in the diet of Nile tilapia: digestibility and growth performance. Aquac Int 7:57–67
Forster I, Higgs DA, Dosanjh BS, Rowshandeli M, Parr J (1999) Potential for dietary phytase to improve the nutritive value of canola protein concentrate and decrease P output in rainbow trout (Oncorhynchus mykiss) held in 11°C fresh water. Aquaculture 179:109–125
Francis G, Makkar HPS, Becker K (2001) Antinutritional factors present in plant-derived alternate ash feed ingredients and their effects in ash. Aquaculture 99:197–227
Helland SJ, Grisdale-Helland B, Nerland S (1996) A simple method for the measurement of daily feed intake of groups of fish in tanks. Aquaculture 139:157–163
Jackson LS, Li MH, Robinson EH (1996) Use of microbial phytase in channel catfish (Ictalurus punctatus) diets to improve utilization of phytate P. J World Aquac Soc 27:309–313
Jahan P, Watanabe T, Kiron V, Satoh S (2003) Improved carp diets based on plant protein sources reduce environmental P loading. Fish Sci 69:219–225
Lall SP (1991) Digestibility, metabolism and excretion of dietary P in fish. In: Cowey CB, Cho CY (eds) Nutritional strategies and aquaculture waste. Proceedings of the first international symposium on nutritional strategies in management of aquaculture waste. University of Guelph, Ontario, Canada, pp 21–36
Lall SP (2002) The minerals. In: Halver JE, Hardy RW (eds) Fish nutrition. Academic Press, San Diego, pp 259–308
Lanari D, D’Agaro E, Turri C (1998) Use of nonlinear regression to evaluate the effects of phytase enzyme treatment of plant protein diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 161:345–356
Liu BL, Rafing A, Tzeng YM, Rob A (1998) The induction and characterization of phytase and beyond. Enzyme Microb Technol 22:415–424
Mabahinzireki GB, Dabrowski K, Lee KJ, EI-Saidy D, Wisner ER (2001) Growth, fed utilization and body composition of tilapia (Oreochromis sp.) fed with cottonseed meal-based diets in a recirculating system. Aquac Nutr 7:189–200
Masumoto T, Tamura B, Shimeno S (2001) Effects of phytase on bioavailability of P in soybean meal-based diets for Japanese flounder (Paralichthys olivaceus). Fish Sci 67:1075–1080
National Research Council (1993) Nutrient requirements of fish. National Academic Press, Washington, DC
Oliva-Teles A, Pereira JP, Gouveia A, Gomes E (1998) Utilization of diets supplemented with microbial phytase by seabass (Dicentrarchus labrax) juveniles. Aquat Living Resour 11:255–259
Papatryphon E, Howell RA, Soares JH Jr (1999) Growth and mineral absorption by striped bass (Morone saxatilis) fed a plant feedstuff based diet supplemented with phytase. J World Aquac Soc 30:161–173
Papatryphon E, Soares JH Jr (2001) The effect of phytase on apparent digestibility of four practical plant feedstuffs fed to striped bass (Morone saxatilis). Aquac Nutr 7:161–167
Powers Hughes K, Soares JH Jr (1998) Efficacy of phytase on P utilization in practical diets fed to striped bass (Morone saxatilis). Aquac Nutr 4:133–140
Riche M, Brown PB (1996) Availability of P from feedstuffs fed to rainbow trout (Oncorhynchus mykiss). Aquaculture 142:269–282
Rodehutscord M (1996) Response of rainbow trout (Oncorhynchus mykiss) growing from 50 to 200 g to supplements of dibasic sodium phosphate in a purified diet. J Nutr 126:324–331
Rodehutscord M, Pfeffer E (1995) Effects of supplemental microbial phytase on P digestibility and utilization in rainbow trout (Oncorhynchus mykiss). Water Sci Technol 31:143–147
Sajjadi M, Carter CG (2004) Dietary phytase supplementation and the utilization of P by Atlantic salmon (Salmo salar L.) fed a canola-meal-based diet. Aquaculture 240:417–431
Schaefer A, Koppe WM, Meyer-Burgdorff KH, Gunther KD (1995) Effects of microbial phytase on utilization of native P by carp in a diet based on soybean meal. Water Sci Technol 31:149–155
Singh M, Krikorian AD (1982) Inhibition of trypsin activity in vitro by phytate. J Agric Food Chem 30:799–800
Slominski BA, Davie T, Nyachoti MC, Jones O (2007) Heat stability of endogenous and microbial phytase during feed pelleting. Livest Sci 109:244–246
Snedecor GW, Concbran WG (1978) Statistical methods, 6th edn. Iowa State University Press, Ames
Spinelli J, Houle CR, Wekell JC (1983) The effect of phytates on the growth of rainbow trout (Salmo gairdneri) fed purified diets containing varying quantities of Ca and Mg. Aquaculture 30:71–83
Storebakken T, Shearer KD, Roem AJ (1998) Availability of protein, P and other elements in fish meal, soy-protein concentrate and phytase-treated soy-protein-concentrate-based diets to Atlantic salmon (Salmo salar). Aquaculture 161:365–379
Sugiura SH, Dong FM, Hardy RW (1998) Effects of dietary supplements on the availability of minerals in fish meal; preliminary observations. Aquaculture 160:283–303
Sugiura SH, Gabaudan J, Dong FM, Hardy RW (2001) Dietary microbial phytase supplementation and the utilization of P, trace minerals and protein by rainbow trout [Oncorhynchus mykiss (Walbaum)] fed soybean meal-based diets. Aquac Res 32:583–592
Teskeredzic Z, Higgs DA, Dosanjh BS, McBride JR, Hardy RW, Beames RM, Simell M, Vaara T, Bridges RB (1995) Assessment of unphytinized and dephytinized rapeseed protein concentrate as sources of dietary protein for juvenile rainbow trout (Oncorhynchus mykiss). Aquaculture 131:261–277
Vielma J, Lall SP, Koskela J, Schoner FJ, Mattila P (1998) Effects of dietary phytase and cholecalciferol on P bioavailability in rainbow trout (Oncorhynchus mykiss). Aquaculture 163:309–323
Vielma J, Mäkinen T, Ekholm P, Koskela J (2000) Influence of dietary soy and phytase levels on performance and body composition of large rainbow trout (Oncorhynchus mykiss) and algal availability of P load. Aquaculture 183:349–362
Vielma J, Ruohonen K, Peisker M (2002) Dephytinization of two soy proteins increases P and protein utilization by rainbow trout (Oncorhynchus mykiss). Aquaculture 204:145–156
Vielma J, Ruohonen K, Gabaudan J, Vogel K (2004) Top-spraying soybean meal-based diets with phytase improves protein and mineral digestibilities but not lysine utilization in rainbow trout [Oncorhynchus mykiss (Walbaum)]. Aquac Res 35:955–964
Wise A (1983) Dietary factors determining the biological activities of phytate. Nutr Abs Rev 53:791–807
Yan W, Reigh R, Xu Z (2002) Effects of fungal phytase on utilization of dietary protein and minerals, and dephosphorylation of phytic acid in the alimentary tract of channel catfish (Ictalurus punctatus) fed an all-plant-protein diet. J World Aquac Soc 33:10–22
Yoo GY, Wang X, Choi S, Han K, Kang JC, Bai SC (2005) Dietary microbial phytase increased the P digestibility in juvenile Korean rockfish (Sebastes schlegeli) fed diets containing soybean meal. Aquaculture 243:315–322
Acknowledgements
The place of study was supported by Harbin Academy of Agricultural Sciences Fisheries Research Institute and the experiment was financially supported by grant # 2005AA6CN183 from Harbin Science and Technology Project.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wang, F., Yang, YH., Han, ZZ. et al. Effects of phytase pretreatment of soybean meal and phytase-sprayed in diets on growth, apparent digestibility coefficient and nutrient excretion of rainbow trout (Oncorhynchus mykiss Walbaum). Aquacult Int 17, 143–157 (2009). https://doi.org/10.1007/s10499-008-9187-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10499-008-9187-5