
Abstract
Apoptosis plays a major role in controlling both the rate of sperm production and chromosomal abnormalities in adult male testes. However, little is known on the mechanisms controlling induction and execution of apoptosis under physiological conditions. In this work we have uncovered a major role for the cell death receptor Fas in both the extrinsic and intrinsic pathways in normal germ cell apoptosis. We show here that Fas levels increased significantly in a group of germ cell in 25 d old rats, which were identified as spermatocytes and only a few spermatogonia. In addition, we show that isolated spermatocytes expressing high levels of Fas display activation of caspase-8, -9, -3, -6 and -2, as well as increased levels of intracellular calcium and decreased pH, which coincides with stabilization of p53, and transcriptional activation of PUMA and Fas. Therefore, our data strongly suggests that transcriptional up regulation of Fas could predispose a group of spermatocytes to Fas ligand triggering apoptosis by the extrinsic and intrinsic pathway.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Normal spermatogenesis, the process by which male germ cells mature from spermatogonia via spermatocytes and spermatids to spermatozoa, involves a delicate balance between cell proliferation and death. Testicular disorders and infertility have been associated with deregulation of death cell pathways, which leads to an imbalance between proliferation and death [1–6].
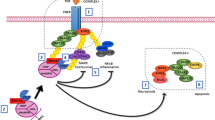
The Fas system has been implicated as a possible regulator of germ cell apoptosis in the rat testis [2, 7–10]. Fas (Apo/CD95) is a type I transmembrane receptor that belongs to the tumor necrosis factor (TNF)/nerve growth factor receptor family [11]. Binding of Fas to its ligand FasL forms a highly stable supramolecular cluster which promotes processing and stabilization of caspase-8 [12–14]. Once activated, caspase-8 can proteolytically activate the effectors caspases-3, -6 or -7, which degrade a large number of cellular proteins that ultimately kill the cell [15]. On the other hand, the intrinsic apoptotic pathway is activated by stress, starvation or radiation, and it starts with the assembly of a multimeric complex (apoptosome) involving Apaf-1, procaspase-9 and cytochrome c [16, 17]. Once activated, caspase-9 cleaves and activates caspases-3, -6 and -7. Based on the kinetics and patterns of caspase activation two types of cells can be defined. In type I cells, caspase -8 mediated activation of caspase-3 is sufficient to ensue execution of the final steps of apoptosis. In contrast, in type II cells caspase-8 mediated activation is less efficient and/or activation of the effectors caspases is inhibited by members of the inhibitor of apoptosis (IAP) protein family [11, 15, 18]. In these cells, a caspase-8 generated cleavage product of BID (tBID), a BH3-only proapoptotic protein, links the extrinsic and intrinsic pathways of apoptosis, contributing to apoptosis by inducing the release of cytochrome c, and other apoptogenic proteins from the mitochondria [17].
We recently showed that caspases-8, -9 and -3 are activated exclusively in germ cells during the first wave of spermatogenesis, with a maximum activity in rats of 25 d of age [19]. This peak in caspase activity coincides with the highest rate of germ cell apoptosis as evaluated by TUNEL. Since we found activity of caspases-8 and -9, it is very likely that germ cells behave like type II cells triggering both the extrinsic and intrinsic pathways. Although it is accepted that the Fas/FasL system is present in testes, its roles during germ cell apoptosis is not well defined [20, 21]. Fas receptors have been localized to germ cells, whereas FasL expression has been localized to Sertoli cells [8]. Several toxicants like mono-(2-ethylhexyl) phthalate or 2,5-hexanedione, induce up-regulation of FasL and therefore a massive germ cell apoptosis [7, 8, 22]. However mutant mice showing inactivation of Fas receptor or FasL show no evidence of abnormal spermatogenesis [20, 23, 24]. However, it is possible that in the absence of Fas receptors, germ cells could turn on alternative death pathways, which are not activated under normal physiological conditions.
p53 is a well-characterized tumor suppressor protein, which can induce apoptosis, either by inducing transcription of pro-apoptotic genes or by direct effects on mitochondrial membranes [25, 26]. p53 undergoes posttranslational modifications in response to oncogene-activated signaling pathways or to genotoxic stresses [27]. This modifications allow stabilization of p53, which accumulates in the nucleus and regulates target gene expression [26]. One important mechanism through which p53 mediates its biological response is transcriptional activation of proapoptotic target genes such as death receptors like Killer/DR5 or CD95/Fas, and proapoptotic Bcl-2 proteins like BAX, BID or PUMA [28]. p53 has been found to be expressed in primary spermatocytes and spermatogonia and, along with Fas, it is involved in a sequential mechanism of testicular germ cell apoptosis in cryptorchid testis [10, 29–31]. This evidence suggests that p53 may be linked to Fas expression, and probably to apoptosis, in spermatocytes and spermatogonia. However, there is no direct evidence showing that p53 stabilization/activation is related to Fas-mediated caspase activation and germ cell apoptosis under physiological conditions.
In this work we show for the first time that during the first wave of spermatogenesis in the rat, spermatocyte apoptosis is associated with activation/stabilization of p53 and up-regulation of Fas and PUMA, two transcriptional targets of p53. Apoptotic germ cells showed activation of caspases-8, -9, -3, -6, and -2, as well as increased levels of intracellular Ca2+ and decreased pH. Therefore, we propose that under physiological conditions, apoptotic spermatocytes behave like type II cells triggering both the intrinsic and extrinsic pathways. This way of execution of apoptosis is different from other cell death models proposed for germ cells, such as hormone deprivation, heat or genotoxic stress, where the main death mechanism activated seems to be the intrinsic apoptotic pathway. Thus, physiological germ cell death implies a new paradigm in apoptosis during mammalian spermatogenesis.
Materials and methods
Animals
Male Sprague-Dawley rats between 5 to 60 d of age were acquired from the Animal Facility of our Faculty. The rats were housed under a 12L:12D cycle and provided with water and rat chow ad libitum. The rats were killed by cervical dislocation after exposure to CO2 for 30 s. Investigations were conducted in accordance with the Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching, published by the Consortium for Developing a Guide for the Care and Use of Agricultural Animals in Agricultural Research and Teaching, First Edition, 1988; the National Research Council (NRC) publication Guide for Care and Use of Laboratory Animals (copyright 1996, National Academy of Science).
Chemicals and antibodies
Antibodies against Fas (sc-1023), BID (sc-11423), BARD1 (sc-11438), caspase-8 (sc-7890) and p53 (sc-126) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). Antibody against SCP3 was purchased from Abcam (Cambridge, UK) and antibody against vimentin (clone V9) was purchased from LabVision (Fremont, CA). Anti-Golgin160 was prepared as previously described [32]. Antibodies against active caspase-9 (cat 9506) and caspase-3 (Cat 9665) were purchased from Cell Signaling (Danvers, MA). 2′,7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein (BCECF) and Fluo-3 were purchased from Molecular Probes (Eugene, OR). Anti-rabbit and anti-mouse UltraVision Detection Systems were obtained from LabVision (Fremont, CA).
Immunohistochemistry
Fas receptors were assayed in paraffin embedded cross-sections of rat testis fixed in 4% paraformaldehyde (PFA). The samples were first treated with 3% H2O2 for 5 min, then, to prevent unspecific binding, a standard protein block System (Ultra V block, LabVision, Freemont, VA) was applied for 5 min. Primary antibody against Fas was applied at a concentration of 2 μg/ml and incubated overnight at 4°C in a humidified chamber after being washed twice for 5 min in a Tris–HCl buffer, pH 7.6 with 0.3 M NaCl and 0.1% Tween 20. Biotinylated secondary antibody, streptavidin–biotinylated–peroxidase complex, amplification reagent (biotinyl tyramide) and peroxidase-conjugated streptavidin were applied step by step for 15 min each. Afterwards, incubation slides were washed twice in a buffer for 3 min each. Finally, substrate-chromogen solution consisting of concentrated Tris–HCl and 0.8% H2O2 (substrate) and 3,3-diaminobenzidine tetrahydrochloride (DAB) solutions (chromogen) were applied for 5 min and washed in distilled water. Samples were observed under a phase contrast microscope (Optiphot-2, Nikon, Japan) and photographed with a digital camera (CoolPix 4500, Nikon, Japan).
Immunofluorescence
Rat testis were fixed in 4% PFA and embedded in paraffin. Sections 5–7 μm were re-hydrated and then unspecific binding sites were blocked by incubating sections for 1 h in a PBS plus 2% bovine serum albumin (PBS-BSA) solution. Tissue sections were then incubated overnight with an antibody against Fas (2 μg/ml) at room temperature in a humidified chamber. After washing in PBS, tissue sections were incubated with Alexa 488-goat anti-rabbit IgG (Molecular Probes, Eugene, OR) for 1 h at room temperature, washed, and mounted using a fluorescence protector medium (VectaShield, Burlingame, CA). Sections were observed under phase contrast and fluorescence microscopy (Optiphot-2, Nikon, Japan) and photographed with a digital camera (CoolPix 4500, Nikon, Japan).
Immunocytochemistry
Coverslips with Fas-positive cells were placed in PBS containing 2% formaldehyde and fixed for 1 h. Following fixation the samples were permeated for 60 min in PBS containing 1% Triton X-100. Nonspecific reactions were blocked by further incubation in PBS containing 2 mg/ml BSA and 130 mM glycine. Coverslips were incubated overnight with the antibody against Fas (2 μg/ml) or synaptonemal complex protein 3 (SCP3) (0.08 μg/ml) at room temperature in a humidified chamber. After extensive washing with PBS containing 0.1% Triton X-100, the samples were sequentially labeled with an anti-rabbit donkey antibody conjugated with Fluorescein-isotiociante (FITC) (Molecular Probes, Eugene, OR) for 1 h at room temperature. Following these incubations, coverslips were mounted in VectaShield mounting medium (Vector Labs, Burlingame, CA) and sealed with nail polish. Samples were observed under phase contrast and fluorescence microscopy (Optiphot-2, Nikon, Tokyo, Japan) and photographed with a digital camera (CoolPix 4500, Nikon, Tokyo, Japan).
Protein extraction and western blot
Protein extraction was made by homogenizing isolated seminiferous tubules in buffer A (1% Triton X-100, NaCl 1 M, EDTA 1 mM, PMSF 10 mg/ml, Tris-HCl 20 mM pH 7.0) and then centrifuged for 10 min at 9,300×g. The samples were run on a 12% polyacrylamide gel (SDS-PAGE) under reducing and denaturing conditions, and then transferred to nitrocellulose at 30 V overnight or 100 V for 1.5 h. Nitrocellulose was blocked with 2% BSA in PBS, pH 7.4, and then incubated overnight at 4°C with anti-Fas (2 μg/ml), anti-p53 (0.2 μg/ml), anti-SCP3 (0.008 μg/ml) anti-BARD1 (0.4 μg/ml), vimentin (0.4 μg/ml), caspase-3 (0.2 μg/ml), active caspase-9 (0.3 μg/ml) caspase-8 (0.4 μg/ml), BID (0.4 μg/ml) or anti-Golgin160 (0.8 μg/ml) antibodies. After extensive washing with PBS plus 0.05% Tween 20 (PBS-Tween), the membrane was incubated with a secondary antibody conjugated to peroxidase (KPL, Gaithersburg, Maryland) diluted 1:10.000 in PBS-BSA for 1 h at room temperature. Protein bands were revealed using the Super Signal West Pico chemiluminescent substrate (Pierce, Rockford, IL).
Isolation of Fas-positive and Fas-negative cells
Testes from 25 days-old rats were dissected into a Petri dish with EKRB (enriched Krebs-Ringer bicarbonate) medium containing 120.1 mM NaCl, 4.8 mM KCl, 25.2 mM NaHCO3, 1.2 mM KH2PO4 (pH 7.2), 1.2 mM MgSO4 z 7H2O, 1.3 mM CaCl2, supplemented with 11.1 mM glucose, 1 mM glutamine, 10 ml/L MEM essential amino acid solution (GIBCO/Invitrogen, Carlsbad, CA), 10 ml/l BME nonessential amino acid solution (GIBCO/Invitrogen, Carlsbad, CA), 100 mg/ml streptomycin, and 100 U/ml penicillin (K salt). Dry collagenase was then added at a final concentration of 0.5 mg/ml. The testes were then incubated for 15–45 min at 32°C with gentle stirring. Once the seminiferous tubules were dispersed in the medium, they were allowed to settle at the bottom of the dish, and the medium was aspirated and discarded. Germ and Sertoli cells were mechanically dissociated by aspirating the tubules with 18 G and 20 G syringes. Dissociated cells were incubated with anti-Fas antibody (5 μg/ml) in ERKB medium for 2 h at 33°C with gentle agitation. Then we added 50 mg of Protein A-conjugated with agarose (Sigma co, St Louis, MO) and the suspension was incubated for 1 h. The mixture was centrifuged at 600 g for 30 sec and the supernatant (Fas-negative cells) was kept in a separate tube. The pellet was washed twice with ERKB medium and the bound cells (Fas-positive cells) were released by adding a blocking peptide for the anti-Fas antibody.
Caspase activity measurements
Isolated Fas-negative and Fas-positive cell suspensions were homogenized in a buffer containing 1 M NaCl, 1 mM EDTA, 10 μg/ml PMSF, 1%Triton X-100, 20 mM Tris-HCl, pH 7.4. The colorimetric substrates for caspase-3 (Ac-DEVD-pNA), caspase-6 (Ac-VEID-pNA), caspase-8 (Ac-IETD-pNA), caspase-9 (Ac-LEHD-pNA) and caspase-2 (Ac-VDVAD-pNA) were purchased from Calbiochem (Darmstadt, Germany). Substrates were labeled with the chromophore p-nitroaniline (pNA) which is released upon caspase cleavage. Free pNA produces a yellow color that is monitored spectrophotometrically at 405 nm. The amount of yellow color produced upon cleavage is proportional to the amount of caspase activity present in the sample. To minimize cross reactivity, inhibitors of caspase-3 (DEVD-fmk), caspase-8 (IETD-fmk) or caspase-9 (LEHD-fmk) were included at a concentration of 1.5 μM when measuring the activities of caspase-8 and caspase-3, respectively [33].
Intracellular pH measurements
Fas-negative and Fas-positive cells were loaded with 2′, 7′-bis-(2-carboxyethyl)-5-(and-6)-carboxyfluorescein (BCECF) by incubating the cells in 1 μM BCECF-AM (acetoxymethyl ester) for 30 min at room temperature. The cells were subsequently washed by centrifugation and allowed to settle in a microscope chamber made of Plexiglas with a glass cover slip at the bottom (Ridout Plastics, San Diego, CA). Successive images were taken when the cells were excited with 440 and 500 nm lights, receiving emitted light above 520 nm. The digital images were processed using a public domain image processing software (ImageJ, NIH, USA). The digital ratio between the images obtained at 500/440 nm is related to intracellular pH by the formula pH=pKa+log[(R max−R)/(R−R min)] [34, 35].
Intracellular Ca2+ measurements
Fas-positive and Fas-negative cells were loaded with Fluo-3 by incubating the cells in 5 μM Fluo-3-AM (acetoxymethyl ester) for 1 h at room temperature. The cells were subsequently washed by centrifugation and allowed to settle in a microscope chamber made of Plexiglas with a glass cover slip at the bottom. Digital images were taken when the cells were excited with 440–490 nm, receiving emitted light above 520 nm. To obtain an estimated intracellular Ca2+ concentration, an image of Fluo-3 fluorescence was taken, followed by addition of 1 μM ionomycin, new images were taken, 2 mM MnCl2 added (quenching Fluo-3-Ca2+ fluorescence), new images were again taken, followed by addition of digitonin (20 μg/ml) in order to obtain a final background image. The digital images were processed using a public domain image processing software (Image J, NIH, USA). Intracellular Ca2+ concentrations were calculated using the formula described previously [36].
RT-PCR
Total RNA was isolated using Trizol-Reagent (Invitrogen, Carlsbad, CA). First, complementary DNA was made using 5 μg total RNA in the presence of Superscript III reverse Transcriptase (In Vitrogen, Carlsbad, CA) and random primers. After the RT reaction 51 μl of the incubation mixture were used as a template for the subsequent PCR reaction. Several primer sets were used to obtain the PCR products of Fas; 5′-GTGCAAGGCTCAAGGATGT-3′ and 5′TGTCAACCTGTCAGCCTG-3′; p53 5′-CACAACTGCACAGGGCATGT -3′ and 5′-ATATGAGCATCGAGCTCCCTCT-3′; PUMA 5′-TGCACTGATGGAGATACGGACTT-3′ and 5′-ACCATGAGTCCTTCAGCCCTC-3′ GAPDH 5′-TCCACCACCCTGTTGCTGTA-3′ and 5′-ACCACAGTCCATGCCATCAC-3′ in the same conditions as previously described [7, 8, 25, 37, 38].
Statistical analysis
For mean comparisons, we used analysis of variance (ANOVA). When the ANOVA test showed statistical differences, the Student–Newman–Keuls (SNK) test was used to discriminate between groups. The 2-test was used for comparison of frequencies. Statistical significance was defined as p < 0.05 [39].
Results
Immunolocalization of Fas during post-natal development of the rat testis
First, we evaluated Fas localization in paraffin-embedded tissue sections of rat testes from perinatal age to adulthood. The Fas antigen was never detected in testis during the perinatal or prepubertal period (5–15 days; Fig. 1(B, C)). At the pubertal period (25 days), Fas gave a strong signal in a few spermatogonia, but mostly in spermatocytes (Fig. 1(D)). At the young adult period (35 days), Fas staining was weakly observed in spermatogonia and spermatocytes (Fig. 1(F)). Rats 45 d old showed as few Fas-positive germ cells as shown for 35 d old testes (data not shown). Figure 1(G) shows the number of Fas positive cells throughout the studied period in relation to the number of seminiferous tubules. It was clearly seen that the highest number of Fas-positive cells was in 25 d old rats, in contrast to a low frequency of Fas-positive cells in adult rats (p < 0.001). Pre-incubation of the primary antibody with the blocking peptide did not show any reaction (Fig 1(A)).

Developmental immuno expression of Fas in rat testes: Testis sections were incubated with an antibody raised against Fas . (A) negative control obtained pre-incubating the primary antibody with a blocking peptide, (B) Postnatal Day 5, (C) Postnatal Day 15 testis, (D, E) Postnatal Day 25 testis, (F) Postnatal Day 35 testis, (G) Quantification of the number of Fas-positive germ cells per seminiferous tubules at each age. At least 100 tubules were quantified in three different animals for each age. The numbers represent the mean ± S.D. Fas gave a strong signal in spermatocytes (Fig. 1(E), arrow) and in a few spermatogonia (Fig. 1(E), arrowhead). Bar 100 μm
We also found that all the tubules that had Fas-positive cells were in stages VI–VIII (Fig. 1(D)). Most of the Fas-positive cells were found adjacent to the luminal space and they were classified as late pachytene spermatocytes according to size and chromatin condensation (Fig. 1(E), arrow). Furthermore, we occasionally found some strongly labeled cells attached to the basement membrane that according to their nuclear morphology were classified as spermatogonia (Fig. 1(E), arrowhead). Therefore, these results indicate that spermatogonia, but mostly spermatocytes, represent a distinct Fas-positive cell population in pubertal rats. We then studied whether Fas-positive germ cells were also apoptotic cells or if this receptor could be involved in a process unrelated to apoptosis. Hence, we assayed TUNEL and the presence of Fas in the same cross sections of 25 d old rat testis by confocal microscopy. Germ cells showed strong TUNEL labeling and seemed localized towards the lumen of the seminiferous tubules (Fig. 2(A)). All germ cells showed Fas signal (Fig. 2(B)). Superimposition of both images showed that 57 ± 5% of the TUNEL-positive cells co-localized with Fas (Fig. 2(D)). In order to determine whether the Fas levels differed between TUNEL–positive and TUNEL-negative cells, we made a densitometric analysis of Fas using the program Image J. Despite the fact that the mean level of Fas in TUNEL-positive cells appeared higher than TUNEL-negative cells this difference was not statically significant (Fig. 2(E)). Then, we quantified the levels of Fas in each cell according to a relative pseudo colored scale from an immunofluorescence image obtained using an anti-Fas antibody (Fig. 3(A, B)). Figure 3 shows the bright field image (Fig. 3(A)), the immunefluorescence image (Fig. 3(B)) and the pseudocolored intensity of Fas-positive cells in seminiferous tubules of 25 d old rat testis (Fig. 3(C)). The fluorescence intensity in Fig. 3(C) was adjusted to give a blue background corresponding to control samples with the antibody previously pre-adsorbed with the immunizing peptide (not shown). The gradient from blue to red (see bar scale) represents the intensity of Fas labeling in the tissue expressed in arbitrary counts per pixel. Light blue or green cells were classified as low intensity Fas expressing cells, yellow cells as medium intensity, and red colored cells as high intensity Fas expressing cells. Although Fas was expressed by all germ cells, two patterns of intensity were more evident: (a) seminiferous tubules with an open lumen with high intensity labeled cells in the ad luminal or luminal compartments, and (b) seminiferous tubules with a closed lumen and a high intensity Fas label on basal cells. Figure 3(D) shows that the largest percentage of labeled cells was ad luminal, followed by basally located cells. High intensity-labeled cells were located at the basal compartment followed by cells at the ad luminal compartment. Our results strongly suggest that there are germ cells expressing different levels of Fas in spermatogenic cells. Those cells expressing high levels of Fas are localized at the basal, ad luminal and luminal compartments of the seminiferous tubules. The majority of cells expressing Fas were localized to the ad luminal or luminal compartments in the seminiferous tubules, which according to immunohistochemistry, corresponded to spermatocytes (Fig. 1).

Co-localization of TUNEL and Fas expression in 25 d old rat testes: (A) Fas localization and (B) TUNEL were detected in the same paraffin-embedded cross tissue section of 25 d old rat testis. (C) Merge image showing that most of the TUNEL germ cells have a Fas signal at the plasma membrane. (D) Quantification of co-localization of TUNEL with Fas positive cells (gray bars). (E) Quantification of Fas level expression in TUNEL-positive and TUNEL-negative germ cells. Results show the quantification of 100 determinations at three different points around each cell, from three different rats (n=3)

Differential localization of germ cells according to their Fas levels: (A) Phase contrast, (B) immunofluorescence and (C) pseudo colored microphotograph of immunofluorescence against Fas in 25 day-old rat testis. The scale bar to the right shows the fluorescence intensity displayed by individual cells. (D) Quantitative analysis of Fas intensity according to low (green), medium (yellow) and high (red) fluorescence intensity. The graph shows that most of the germ cells (87%) expressing high levels of Fas are located towards the lumen of the seminiferous tubules. Bar 100 μm
Apoptosis in germ cells is associated with Fas up-regulation and caspase activation
We hypothesized that germ cells with high levels of Fas (mostly spermatocytes as judged by immunohistochemistry) in 25 d old rats, could trigger caspase activation and apoptosis. In order to characterize these cell populations, we attempted to isolate Fas (apoptotic)-positive cells from 25 d old rats. De-capsulated testes were treated with collagenase and then isolated seminiferous tubules were mechanically disrupted, washed and reacted with antiFas antibody. After extensive washing, the germ cells where immuno-separated using protein A-conjugated with agarose, a procedure similar to isolated GFRa-1-positive cells and modified as described in Methods (Van der Wee et al., 2001). Thus, we obtained two different cell populations, those bound (B) to the immunoaffinity beads (Fas-positive) and those not bound (NB) to the beads column (Fas-negative). In three replicate experiments, indirect immunofluorescence against Fas of immuno-separated (Fas-positive) cells, and released from Sepharose-protein A beads using the blocking peptide, showed that all cells presented a strong label on the plasma membrane (Fig. 4(A)). Immunofluorescence of Fas-positive cells with an antibody against SCP3, which is involved in the synaptonemal complex of meiotic cells, indicated that all isolated cells were meiotic spermatocytes (Fig. 4(B)). A control experiment without the primary antibody or with the antibody pre-adsorbed with the blocking peptide did not give any signals (Fig. 4(C)). A Western blot for SCP3 showed a clear band in both cell fractions, showing that the isolated Fas-positive cells corresponded to meiotic spermatocytes (Fig. 4(D)). Only Fas-negative cells showed a strong band for vimentin, a protein of the intermediate filaments which is present in Sertoli and myoid cells, but not in germ cells (Fig. 4(D)). In addition, Fas-positive but not Fas-negative cells showed a strong Fas band (Fig. 9(A)). Therefore, with reasonable certainty, we can affirm to have isolated meiotic spermatocytes expressing high levels of Fas from 25 d old rat testis.

Isolation and characterization of Fas-positive spermatocytes from 25-day-old rat testes: (A) Indirect immunofluorescence against the synaptonemal complex protein SCP3 and (B) Fas of immuno-isolated germ cells. The picture shows that all isolated germ cells displayed a strong and sharp surface reactivity against Fas and SCP3. (C) Negative control with primary antibody pre-incubated with the blocking peptide. Inserts are the bright field of each picture. (D) Western blot of the bound (B) and not bound (NB) germ cells to the Fas immunoaffinity column. Both cells populations displayed a sharp band of SCP3, but only the NB cells showed a strong reaction against vimentin

Caspase detection and activation in isolated Fas-positive germ cells from 25 d old rats: Fas-positive and Fas-negative cells were prepared from isolated seminiferous tubules and analyzed by SDS-PAGE using specific antibodies against different proteins as indicated to the left side of the figure. The relative molecular weight of each band is shown to the right side of the figure. The image is representative from three different sets of experiments. The Golgi protein Golgin-160 (Gol160) is shown as load control for all samples

Fas-positive cells have high caspase activity: Fas-negative and Fas-positive isolated germ cells from 25 d old rat testis were homogenized. Caspase activity was determined in the protein supernatant using a sensitive colorimetric method specific for each caspase. The graph shows the proportion of specific enzyme activity found in each cell population. The numbers represent the mean of three different experiments (* p < 0.05, ** p < 0.01, *** p < 0.001)

[Ca 2+] i levels in isolated Fas-positive germ cells from 25 d old rats testes: Cells pre-incubated with anti-Fas antibody and separated by binding to agarose-protein A beads (Fas+) and unbound cells (Fas−) where loaded with fluo-3 and subsequently placed in a microscope chamber in Krebs-Henseleit with DL-lactate (10 mM) at 33°C. Their [Ca2+]i concentrations were estimated by taking consecutive fluorescent images (excitation: 440–480 nm, emission: >520 nm) in basal conditions, in the presence of ionomycin (5 μM), MnCl2 and digitonin (20 μg/ml). [Ca2+]i was calculated as described by Kao et al. (1989)

pH i levels in isolated Fas-positive germ cells from 25 d old rats testes: The cells preincubated with anti-Fas antibody and separated by binding to agarose-protein A beads (Fas+) and unbound cells (Fas−) where loaded with BCECF and subsequently placed in a microscope chamber in Krebs-Henseleit with DL-lactate (10 mM) at 33°C. Their pHi values were estimated by taking consecutive fluorescent images with excitations at 380 and 340 nm and emission >410 nm. pHi was calculated as described by Bright et al. (1989)
Procaspase-8 cleavage is a hallmark of death-receptor activation. Procaspase-8 was present primarily as a band of approximately 55 kDa in both Fas-positive and Fas-negative cell fractions. On the other hand, Fas-positive cells showed processing of procaspase-8 efficiently cleaved to a fragment of approximately 20 kDa (Fig. 5). Full length procaspase-9 (46 kDa) is processed upon induction of apoptosis and yields fragments of 37/35 kDa. An antibody against active caspase-9 detected two strong bands around 37/35 kDa only in Fas-positive cells (Fig. 5). In both cell populations, procaspase-3 was present in a 32 kDa form. However, Fas-positive cells showed a strong band corresponding to the processed active form of caspase-3 (17 kDa) (Fig. 5). To estimate caspase-3 activity we determined cleavage of poly (ADP-ribose) polymerase-1 (PARP) a substrate of caspase-3. The full length form of PARP (116 kDa) was readily detected in Fas-negative cells, but it was almost completely cleaved to the 85 kDa form in Fas-positive cells (Fig. 5). Caspase-8 has been reported to cleave Bid, which then links the death receptor-mediated extrinsic pathway to the intrinsic mitochondrial apoptotic pathway. Full length Bid was present in both cell populations, but its cleaved active form (tBid of 15 kDa) showed a strong band only in Fas-positive cells (Fig. 5). The structural protein of the Golgi apparatus Golgin 160 was used a load control for all the samples. Since immunoblot analysis detects caspases and their proteolytic processing, but does not directly measure caspase activity, the activity of several caspases was assessed using a colorimetric substrate cleavage assay. Using the same extracts prepared for immunoblot analysis we found that Fas-positive cells had a significantly higher activity of initiator caspases-8, -9, and executioner caspases-3 and -6, than Fas-negative cells (Fig. 6). Caspase-2 may act upstream of the mitochondria to promote cytochrome c release and apoptosis. We detected caspase-2 activity in both cell fractions, but Fas-positive cells had a significantly higher proportion than Fas-negative cells (Fig. 6). Thus, our results strongly support the hypothesis that Fas-positive cells are germ cells (primary spermatocytes) engaged in caspase-mediated cell death.
Intracellular [Ca2+] and pH in immuno-isolated Fas-positive cells
To study whether the mentioned Fas expressing cell populations and caspase activation patterns were correlated with the changes in intracellular Ca2+ and pHi, we estimated these parameters in Fas-positive and Fas-negative cell populations. Figure 7 shows [Ca2+]i frequency distributions in Fas-positive and Fas-negative cells. Fas-positive cells presented a distribution skewed towards higher [Ca2+]i levels than those of Fas-negative cells. On the other hand, Fas-negative cells showed a relatively normal distribution of pHi values (Fig. 8) but Fas-positive cells presented a distribution with a subpopulation of cells showing pH values lower than pH 7.0, i.e., acid intracellular pH (Fig. 8).
Thus, our results show that a subpopulation of spermatocytes enters apoptosis by up-regulation of Fas, and activation of caspases 8, 9, 2, 3 and 6. These changes were correlated with an increased [Ca2+]I and a decreased pHi, composing the general picture a Fas-induced apoptotic death in this spermatocyte subpopulation.
Fas up-regulation is associated with p53 activation/stabilization in apoptotic spermatocytes
Fas up-regulation at the plasma membrane can be triggered either by an increase at the transcriptional level or by increasing trafficking from the trans-Golgi to the plasma membrane. In both pathways, p53 is somehow increasing the effective concentration of Fas at the plasma membrane [22]. In order to test whether the high levels of Fas correlated with its expression at the mRNA level, we measured Fas and p53 levels at the protein and RNA levels in both Fas-positive and Fas–negative cells. We found that Fas-positive cells showed higher levels of antigenic Fas than Fas-negative cells. The levels of Fas mRNA were also significantly higher in Fas-positive cells than in Fas-negative cells (Fig. 9(A, B, C)). Thus, Fas-positive cells are spermatocytes showing an increased Fas expression both at the transcriptional and translational levels.

Up-regulation of Fas, p53 and PUMA in apoptotic spermatocytes from 25 d old testes of rats: (A) Western blot showing a significantly higher level of Fas in Fas-positive cells than in Fas-negative cells. The transcription factor p53 showed similar levels in both cell populations, but a p53 regulator, BARD1, was expressed at higher levels in Fas-positive cells than in Fas-negative cells. Numbers show the size of the protein. (B) Semi-quantitative RT-PCR of Fas, p53 and PUMA in Fas-positive and Fas-negative cells. Numbers at the right side indicate the size of the amplicon. (C) Densitometric quantification of the RT-PCR showing significantly increased levels of PUMA and FAS mRNA in Fas-positive cells (* p < 0.05)
Immunoblot analysis showed that Fas-positive cells had higher levels of antigenic p53 than Fas-negative cells (Fig. 9(A)). However, mRNA levels were similar in both cell populations (Fig. 9(B, C)). We also examined the presence of BARD1, a protein that interacts with p53 and probably stabilizes p53 at the post-translational level (e.g., Feki et al., 2005). This protein was originally identified as a binding partner for BRCA1 and has also been implicated in the apoptotic pathway involving p53. BARD1 was strongly expressed in Fas-positive cells, but it was barely detected in Fas-negative cells (Fig. 9(A)). The structural protein of the Golgi apparatus Golgin 160 was used a load control for all the samples. Finally, we examined the role of PUMA (p53-upregulated mediator of apoptosis), a newly discovered pro-apoptotic member of the Bcl-2 family, and involved in p53-mediated mitochondria membrane permeabilization and apoptosis. Our results showed that PUMA mRNA was significantly higher in Fas-positive than in Fas-negative cells (Fig. 9(B, C)). Therefore, Fas-positive cells represent spermatocytes showing increased levels of p53, BARD1, Fas, and PUMA. These results, strongly suggest that p53, Fas, and the extrinsic apoptotic pathway, play a role in germ cell apoptosis during the first wave of spermatogenesis in the rat.
Discussion
The Fas system has been implicated as a possible regulator of germ cell apoptosis in the rat testis, especially under some pathological conditions such as; thermal stress, hormone deprivation or environmental toxicants [7, 20, 22, 40–43]. We recently showed that cell death during the first wave of spermatogenesis implied activation of multiple caspases, suggesting an active role of the receptor mediated apoptotic pathway in this process [19]. In this work, we showed that physiological apoptosis of spermatocytes during the first wave of spermatogenesis was associated with Fas up-regulation, an increase in [Ca2+]i, a decrease in pHi and activation of caspases, involving both the extrinsic and intrinsic pathways. Furthermore, this pattern of cell death induction in spermatocytes seems to involve activation/stabilization of p53, and transcriptional activation of some target genes like Fas and PUMA.
We observed differential expression of Fas during the first wave of spermatogenesis, with the highest number of germ cells expressing Fas in 25 d old rats. These findings agree with previous studies showing that the maximum number of TUNEL positive cells occurred in rats 3 weeks old [19, 44–46]. Similarly, these findings agree with our previous work showing activation of caspase-8 in 25-d old rats, and suggest a role for the death receptor pathway in germ cell apoptosis. Despite the apparent requirement of Fas and FasL for damage-induced germ cell apoptosis, lpr mice containing a spontaneous loss-of-function mutation in the Fas gene are fertile with apparently normal spermatogenesis [7, 20, 43]. Thus, the exact role of Fas in germ cell apoptosis is still very controversial and not yet clarified. It is possible that in the absence of Fas, death receptors like TRAIL or TNF-R, which are present in spermatocytes, could replace Fas triggering caspase activation [47–49]. On the other hand, Fas up-regulation could be a consequence of apoptosis and not be directly involved in caspase activation. Thus, it is possible that caspase-8, the first caspase paradigmatically activated by Fas, could be activated independently of death receptors, as occurs with anti-cancer drugs or ionizing irradiation [33, 50]. In this model, caspase-8 activation is mediated in a post-mitochondrial event by prior cleavage of caspase-6, followed by caspase-3 activation. In this context, caspase-8 activation is a consequence of the intrinsic pathway triggered by a still unknown signal.
In this paper, we propose a key role for Fas and the death receptor pathway in physiological germ cell apoptosis, an idea that is supported by several of our results. First, we found that most TUNEL positive cells (57 ± 5%) also showed localization of Fas in the plasma membrane. Second, immunohistochemistry showed two types of Fas-positive cells in 25 d old rats: ones having a weak signal, and others showing a strong signal. Those strongly labeled Fas-positive cells were identified as spermatocytes and localized towards the lumen of the seminiferous tubules. However, some spermatogonia located at the basal compartment of the seminiferous tubules were also strongly labeled with the anti-Fas antibody. A similar result was obtained using immunofluorescence against Fas. This image analysis showed that the largest percentage of labeled cells were ad-luminal spermatocytes, followed by basally located cells, most likely spermatogonia, as observed by immunohistochemistry. Thus, two independent immune techniques identified a population of spermatocytes expressing high levels of Fas located towards the ad-luminal compartment of the seminiferous tubules and also a population of basally located Fas positive spermatogonia. A similar situation has been observed in human vascular smooth muscle cells, which display a differential expression of Fas on their surface, making some of them respond to the Fas ligand, as a death stimulus, while other remain resistant [51]. Thus, this seems to be a common mechanism to prone a group of cells to a specific death receptor stimulus in different tissues. In addition, regulation of Fas, or Fas-signaling molecules, has been proposed as a major regulatory mechanism to stimulate apoptosis in several cell types [52].
Caspase-2 is one of the best conserved caspases across species [53]. This enzyme is unique among caspases because it has features of both initiator and effector caspases. Likewise, caspase-2, which is involved in stress-induced apoptosis, is recruited into a large protein complex that contains the death domain–containing protein PIDD, whose expression is induced by p53, and the adaptor protein RAIDD [54]. However, other evidence points towards a role of caspase-2 in death receptor-induced cell death [53]. Thus the role of caspase-2 remains controversial. In this work, we have identified caspase-2 activity in apoptotic germ cells, and our finding agrees with the work by Zheng et al. (2006) showing caspase-2 activation during the first wave of spermatogenesis in mice. Caspase-2 activity was highly concentrated in Fas-positive cells and only a small fraction of the activity was detected in Fas-negative cells, thus its function could be related to p53 activation [46, 53]. However, the role of this caspase in spermatogenesis remains controversial because caspase-2 knockout male mice do not show major reproductive defects [55]. Thus, it is possible that caspase-2 activation is related to a non-apoptotic process or its role in apoptosis could be overcome by other caspases in caspase-2 knockout mice. On the other hand, it is possible that caspase-2 activation may be important in the absence of other apoptotic stimuli, like ablation of Fas receptor in lrp mice.
A key component in this hypothesis of Fas-mediated germ cell death in spermatogenesis is the up-regulation of Fas in apoptotic germ cells. We showed here that Fas levels were higher in apoptotic spermatocytes at the protein and mRNA levels as compared to non apoptotic spermatocytes in 25 days-old rats. In addition, we showed that two transcriptional regulators of Fas, p53 and BRAD1, were activated or stabilized in apoptotic germ cells. A fact that agrees with evidence demonstrating that expression of the tumor-suppressor p53 is important in germ cell apoptosis [29, 56–59]. p53 deficient mice display lower levels of spontaneous and induced germ cell apoptosis as compared to control animals. In addition, these mice express reduced levels of Fas in the testes, suggesting that p53 helps control Fas expression [10, 23, 31]. We have shown here, for the first time, that under physiological conditions p53 and its regulatory protein BARD1, were elevated in a population of apoptotic spermatocytes. BARD1 plays a role as mediator of apoptosis by binding to and stabilizing p53. Similarly, BARD1-repressed cells are resistant to apoptosis [60, 61]. It has been previously shown that spermatogonia express full-length BARD1 mRNA and spermatocytes express predominantly a shorter spliced form BARD1ß which lacks the BRCA1-interacting RING finger but maintains its pro-apoptotic activity [62]. Therefore, BARD1 stabilization seems to be a step involved in spermatocytes apoptosis during spermatogenesis.
p53 functions in the nucleus regulating pro-apoptotic genes, whereas cytoplasmic p53 directly activates pro-apoptotic Bcl-2 proteins to permeate mitochondria and initiate apoptosis . It has been shown that a tripartite nexus between Bcl-xL, cytoplasmic p53, and PUMA coordinates these distinct p53 functions. Nuclear p53 caused expression of PUMA, which then displaced p53 from Bcl-xL allowing p53 to induce mitochondrial permeabilization [25]. In addition, PUMA induces apoptosis, in part, through the BAX-dependent generation of super oxide and hydrogen peroxide [63]. Thus, PUMA expression could lead to both caspase-dependent and caspase-independent germ cell apoptosis. In addition, some evidence suggests that transcriptional up-regulation of PUMA, induced by p53, modulates endoplasmic reticulum calcium pool depletion-induced apoptosis via Bax activation [64]. We have shown here for the first time expression and possible participation of PUMA in germ cell apoptosis. It is possible that PUMA expression may help to promote the intrinsic pathway of apoptosis and, along with cleaved BID (tBID), to induce the release of mitochondrial proapoptic factors like cytochrome c and triggering caspase-9 activation.
Activation of apoptosis has been classically associated with an increase in intracellular Ca2+ concentration ([Ca2+]i), and a decreased intracellular pH (pHi). In fact, apoptosis has been linked to an increased [Ca2+]i, either as a required event for calpain-mediated caspase-3 activation in some cells or as an activator of DNA degradation in other cells [65, 66]. Furthermore, a decrease in pHi seems to precede some apoptotic changes in several cell lines [67]. Isolated Fas positive cells showed activated caspases and higher levels of [Ca2+]i and lower pHi, a pattern of biochemical and physiological markers characteristic of apoptotic cells, reaffirming the notion that up-regulation of Fas is associated with apoptosis induction in these cells.
It is has been shown that stimuli like heat or hormone deprivation promotes only the intrinsic pathway of apoptosis in pachytene spermatocytes [24]. These stimuli led to mitochondria permeabilization, release of cytochrome c, smac/diablo and activation of caspase-9 in apoptotic spermatocytes of heated rat testis [24, 68, 69]. We have shown here that under physiological conditions Fas up regulation and caspase-8 activation appear to play a major role in germ cell demise. We also showed caspase-9 activation and cleavage of the pro-apoptotic protein BID, linking both caspase cascades. Thus, it seems that under physiological conditions germ cells undergoing apoptosis behave like type II cells, where the intrinsic and extrinsic apoptotic pathways are activated. It has been suggested that germ cell apoptosis could be triggered by calpain activation in a caspase-independent mechanism [70]. Our work strongly suggests a role for caspases during germ cell apoptosis during the first wave of spermatogenesis. However, our results showed that approximately 70% of the TUNEL-positive germ cells showed co-localization with FAS. Thus, it is possible to conceive that two independent pathways could trigger germ cell death. On the other hand, several studies show that Fas activation can also stimulate free radical production that could contribute to the observed cell death [71]. Therefore, although the fact that caspase activation and Fas expression seems well established by the published data and this work, the real role of free radicals, caspase-independent mechanisms and how they interplay with caspase-dependent pathways in order to bring about germ cell death remains to be elucidated.
Conclusion
In conclusion, we have demonstrated a novel mechanism of germ cell apoptosis during the first round of spermatogenesis. These results could help to design new strategies oriented to modulate the rate of male germ cell production in mammals.
References
Eguchi J, Koji T, Nomata K, Yoshii A, Shin M, Kanetake H (2002) Fas-Fas ligand system as a possible mediator of spermatogenic cell apoptosis in human maturation-arrested testes. Hum Cell 15(1):61–68
Francavilla S, D′Abrizio P, Cordeschi G, Pelliccione F, Necozione S, Ulisse S, et al (2002) Fas expression correlates with human germ cell degeneration in meiotic and post-meiotic arrest of spermatogenesis. Mol Hum Reprod 8(3):213–220
Francavilla S, D′Abrizio P, Rucci N, Silvano G, Properzi G, Straface E, et al (2000) Fas and Fas ligand expression in fetal and adult human testis with normal or deranged spermatogenesis. J Clin Endocrinol Metab 85(8):2692–2700
Furuchi T, Masuko K, Nishimune Y, Obinata M, Matsui Y (1996) Inhibition of testicular germ cell apoptosis and differentiation in mice misexpressing Bcl-2 in spermatogonia. Development 122(6):1703–1709
Knudson CM, Tung KS, Tourtellotte WG, Brown GA, Korsmeyer SJ (1995) Bax-deficient mice with lymphoid hyperplasia and male germ cell death. Science 270(5233):96–99
Yamamoto CM, Hikim AP, Lue Y, Portugal AM, Guo TB, Hsu SY, et al (2001) Impairment of spermatogenesis in transgenic mice with selective overexpression of Bcl-2 in the somatic cells of the testis. J Androl 22(6):981–991
Lee J, Richburg JH, Shipp EB, Meistrich ML, Boekelheide K (1999) The Fas system, a regulator of testicular germ cell apoptosis, is differentially up-regulated in Sertoli cell versus germ cell injury of the testis. Endocrinology 140(2):852–858
Lee J, Richburg JH, Younkin SC, Boekelheide K (1997) The Fas system is a key regulator of germ cell apoptosis in the testis. Endocrinology 138(5):2081–2088
Pentikainen V, Erkkila K, Dunkel L (1999) Fas regulates germ cell apoptosis in the human testis in vitro. Am J Physiol 276(2 Pt 1):E310–316
Yin Y, Stahl BC, DeWolf WC, Morgentaler A (2002) P53 and Fas are sequential mechanisms of testicular germ cell apoptosis. J Androl 23(1):64–70
Krammer PH (2000) CD95's deadly mission in the immune system. Nature 407(6805):789–795
Henkler F, Behrle E, Dennehy KM, Wicovsky A, Peters N, Warnke C, et al (2005) The extracellular domains of FasL and Fas are sufficient for the formation of supramolecular FasL-Fas clusters of high stability. J Cell Biol 168(7):1087–1098
Sanchez-Gomez MV, Alberdi E, Ibarretxe G, Torre I, Matute C (2003) Caspase-dependent and caspase-independent oligodendrocyte death mediated by AMPA and kainate receptors. J Neurosci 23(29):9519–9528
Scaffidi C, Fulda S, Srinivasan A, Friesen C, Li F, Tomaselli KJ, et al (1998) Two CD95 (APO-1/Fas) signaling pathways. Embo J 17(6):1675–1687
Riedl SJ, Shi Y (2004) Molecular mechanisms of caspase regulation during apoptosis. Nat Rev Mol Cell Biol 5(11):897–907
Budihardjo I, Oliver H, Lutter M, Luo X, Wang X (1999) Biochemical pathways of caspase activation during apoptosis. Annu Rev Cell Dev Biol 15:269–290
Hengartner MO (2000) The biochemistry of apoptosis. Nature 407(6805):770–776
Scaffidi C, Schmitz I, Zha J, Korsmeyer SJ, Krammer PH, Peter ME (1999) Differential modulation of apoptosis sensitivity in CD95 type I and type II cells. J Biol Chem 274(32):22532–22538
Moreno RD, Lizama C, Urzua N, Vergara SP, Reyes JG (2006) Caspase activation throughout the first wave of spermatogenesis in the rat. Cell Tissue Res 325(3):533–540
Richburg JH, Nanez A (2003) Fas- or FasL-deficient mice display an increased sensitivity to nitrobenzene-induced testicular germ cell apoptosis. Toxicol Lett 139(1):1–10
Richburg JH, Nanez A, Williams LR, Embree ME, Boekelheide K (2000) Sensitivity of testicular germ cells to toxicant-induced apoptosis in gld mice that express a nonfunctional form of Fas ligand. Endocrinology 141(2):787–793
Chandrasekaran Y, McKee CM, Ye Y, Richburg JH (2006) Influence of TRP53 status on FAS membrane localization, CFLAR (c-FLIP) ubiquitinylation, and sensitivity of GC-2spd (ts) cells to undergo FAS-mediated apoptosis. Biol Reprod 74(3):560–568
Embree-Ku M, Venturini D, Boekelheide K (2002) Fas is involved in the p53-dependent apoptotic response to ionizing radiation in mouse testis. Biol Reprod 66(5):1456–1461
Sinha Hikim AP, Lue Y, Diaz-Romero M, Yen PH, Wang C, Swerdloff RS (2003) Deciphering the pathways of germ cell apoptosis in the testis. J Steroid Biochem Mol Biol 85(2–5):175–182
Chipuk JE, Bouchier-Hayes L, Kuwana T, Newmeyer DD, Green DR (2005) PUMA couples the nuclear and cytoplasmic proapoptotic function of p53. Science 309(5741):1732–1735
Lu X (2005) p53: a heavily dictated dictator of life and death. Curr Opin Genet Dev 15(1):27–33
Levine AJ, Hu W, Feng Z (2006) The P53 pathway: what questions remain to be explored? Cell Death Differ 13(6):1027–1036
Yu J, Zhang L (2005) The transcriptional targets of p53 in apoptosis control. Biochem Biophys Res Commun 331(3):851–858
Schwartz D, Goldfinger N, Kam Z, Rotter V (1999) p53 controls low DNA damage-dependent premeiotic checkpoint and facilitates DNA repair during spermatogenesis. Cell Growth Differ 10(10):665–675
Sjoblom T, Lahdetie J (1996) Expression of p53 in normal and gamma-irradiated rat testis suggests a role for p53 in meiotic recombination and repair. Oncogene 12(12):2499–2505
Yin Y, Stahl BC, DeWolf WC, Morgentaler A (1998) p53-mediated germ cell quality control in spermatogenesis. Dev Biol 204(1):165–171
Griffith KJ, Chan EK, Lung CC, Hamel JC, Guo X, Miyachi K, et al (1997) Molecular cloning of a novel 97-kd Golgi complex autoantigen associated with Sjogren's syndrome. Arthritis Rheum 40(9):1693–1702
Ding HF, Lin YL, McGill G, Juo P, Zhu H, Blenis J, et al (2000) Essential role for caspase-8 in transcription-independent apoptosis triggered by p53. J Biol Chem 275(49):38905–38911
Bright GR, Whitaker JE, Haugland RP, Taylor DL (1989) Heterogeneity of the changes in cytoplasmic pH upon serum stimulation of quiescent fibroblasts. J Cell Physiol 141(2):410–419
Rink TJ, Tsien RY, Pozzan T (1982) Cytoplasmic pH and free Mg2+ in lymphocytes. J Cell Biol 95(1):189–196
Kao JP, Harootunian AT, Tsien RY (1989) Photochemically generated cytosolic calcium pulses and their detection by fluo-3. J Biol Chem 264(14):8179–8184
Sun XY, Li FX, Li J, Tan YF, Piao YS, Tang S, et al (2004) Determination of genes involved in the early process of embryonic implantation in rhesus monkey (Macaca mulatta) by suppression subtractive hybridization. Biol Reprod 70(5):1365–1373
Yamauchi H, Katayama K, Ueno M, Uetsuka K, Nakayama H, Doi K (2004) Involvement of p53 in 1-beta-D-arabinofuranosylcytosine-induced rat fetal brain lesions. Neurotoxicol Teratol 26(4):579–586
Sokal RR (1995) Biometry: the principles and practice of statistic in biological research. W. H. Freeman, New York, p 887
Mishra DP, Shaha C (2005) Estrogen-induced spermatogenic cell apoptosis occurs via the mitochondrial pathway: role of superoxide and nitric oxide. J Biol Chem 280(7):6181–6196
Nair R, Shaha C (2003) Diethylstilbestrol induces rat spermatogenic cell apoptosis in vivo through increased expression of spermatogenic cell Fas/FasL system. J Biol Chem 278(8):6470–6481
Richburg JH (2000) The relevance of spontaneous- and chemically-induced alterations in testicular germ cell apoptosis to toxicology. Toxicol Lett 112–113:79–86
Richburg JH, Nanez A, Gao H (1999) Participation of the Fas-signaling system in the initiation of germ cell apoptosis in young rat testes after exposure to mono-(2-ethylhexyl) phthalate. Toxicol Appl Pharmacol 160(3):271–278
Boulogne B, Olaso R, Levacher C, Durand P, Habert R (1999) Apoptosis and mitosis in gonocytes of the rat testis during foetal and neonatal development. Int J Androl 22(6):356–365
Rodriguez I, Ody C, Araki K, Garcia I, Vassalli P (1997) An early and massive wave of germinal cell apoptosis is required for the development of functional spermatogenesis. Embo J 16(9):2262–2270
Zheng S, Turner TT, Lysiak JJ (2006) Caspase 2 activity contributes to the initial wave of germ cell apoptosis during the first round of spermatogenesis. Biol Reprod 74(6):1026–1033
Boekelheide K, Lee J, Shipp EB, Richburg JH, Li G (1998) Expression of Fas system-related genes in the testis during development and after toxicant exposure. Toxicol Lett 102–103:503–508
Grataroli R, Vindrieux D, Gougeon A, Benahmed M (2002) Expression of tumor necrosis factor-alpha-related apoptosis-inducing ligand and its receptors in rat testis during development. Biol Reprod 66(6):1707–1715
Suominen JS, Wang Y, Kaipia A, Toppari J (2004) Tumor necrosis factor-alpha (TNF-alpha) promotes cell survival during spermatogenesis, and this effect can be blocked by infliximab, a TNF-alpha antagonist. Eur J Endocrinol 151(5):629–640
Jimbo A, Fujita E, Kouroku Y, Ohnishi J, Inohara N, Kuida K, et al (2003) ER stress induces caspase-8 activation, stimulating cytochrome c release and caspase-9 activation. Exp Cell Res 283(2):156–166
Chan SW, Hegyi L, Scott S, Cary NR, Weissberg PL, Bennett MR (2000) Sensitivity to Fas-mediated apoptosis is determined below receptor level in human vascular smooth muscle cells. Circ Res 86(10):1038–1046
Bentele M, Lavrik I, Ulrich M, Stosser S, Heermann DW, Kalthoff H, et al (2004) Mathematical modeling reveals threshold mechanism in CD95-induced apoptosis. J Cell Biol 166(6):839–851
Zhivotovsky B, Orrenius S (2005) Caspase-2 function in response to DNA damage. Biochem Biophys Res Commun 331(3):859–867
Tinel A, Tschopp J (2004) The PIDDosome, a protein complex implicated in activation of caspase-2 in response to genotoxic stress. Science 304(5672):843–846
Bergeron L, Perez GI, Macdonald G, Shi L, Sun Y, Jurisicova A, et al (1998) Defects in regulation of apoptosis in caspase-2-deficient mice. Genes Dev 12(9):1304–1314
Allemand I, Anglo A, Jeantet AY, Cerutti I, May E (1999) Testicular wild-type p53 expression in transgenic mice induces spermiogenesis alterations ranging from differentiation defects to apoptosis. Oncogene 18(47):6521–6530
Beumer TL, Roepers-Gajadien HL, Gademan IS, van Buul PP, Gil-Gomez G, Rutgers DH, et al (1998) The role of the tumor suppressor p53 in spermatogenesis. Cell Death Differ 5(8):669–677
Fujisawa M, Shirakawa T, Fujioka H, Gotoh A, Okada H, Arakawa S, et al (2001) Adenovirus-mediated p53 gene transfer to rat testis impairs spermatogenesis. Arch Androl 46(3):223–231
Kilinc F, Guvel S, Kayaselcuk F, Aygun C, Egilmez T, Ozkardes H (2004) p53 expression and apoptosis in varicocele in the rat testis. J Urol 172(6 Pt 1):2475–2478
Irminger-Finger I, Busquets S, Calabrio F, Lopez-Soriano FJ, Argiles JM (2006) BARD1 content correlates with increased DNA fragmentation associated with muscle wasting in tumour-bearing rats. Oncol Rep 15(6):1425–1428
Fabbro M, Savage K, Hobson K, Deans AJ, Powell SN, McArthur GA, et al (2004) BRCA1-BARD1 complexes are required for p53Ser-15 phosphorylation and a G1/S arrest following ionizing radiation-induced DNA damage. J Biol Chem 279(30):31251–31258
Feki A, Jefford CE, Durand P, Harb J, Lucas H, Krause KH, et al (2004) BARD1 expression during spermatogenesis is associated with apoptosis and hormonally regulated. Biol Reprod 71(5):1614–1624
Liu Z, Lu H, Shi H, Du Y, Yu J, Gu S, et al (2005) PUMA overexpression induces reactive oxygen species generation and proteasome-mediated stathmin degradation in colorectal cancer cells. Cancer Res 65(5):1647–1654
Luo X, He Q, Huang Y, Sheikh MS (2005) Transcriptional upregulation of PUMA modulates endoplasmic reticulum calcium pool depletion-induced apoptosis via Bax activation. Cell Death Differ 12(10):1310–1318
Scoltock AB, Bortner CD, St JBG, Putney JW Jr, Cidlowski JA (2000) A selective requirement for elevated calcium in DNA degradation, but not early events in anti-Fas-induced apoptosis. J Biol Chem 275(39):30586–30596
Sharma M, Sahu K, Dube A, Gupta PK (2005) Extracellular pH influences the mode of cell death in human colon adenocarcinoma cells subjected to photodynamic treatment with chlorin p6. J Photochem Photobiol B 81(2): 107–113
Collins MK, Furlong IJ, Malde P, Ascaso R, Oliver J, Lopez Rivas A (1996) An apoptotic endonuclease activated either by decreasing pH or by increasing calcium. J Cell Sci 109(Pt 9): 2393–2399
Vera Y, Diaz-Romero M, Rodriguez S, Lue Y, Wang C, Swerdloff RS, et al (2004) Mitochondria-dependent pathway is involved in heat-induced male germ cell death: lessons from mutant mice. Biol Reprod 70(5):1534–1540
Vera Y, Rodriguez S, Castanares M, Lue Y, Atienza V, Wang C, et al (2005) Functional role of caspases in heat-induced testicular germ cell apoptosis. Biol Reprod 72(3):516–522
Coureuil M, Fouchet P, Prat M, Letallec B, Barroca V, Dos Santos C, et al (2006) Caspase-independent death of meiotic and postmeiotic cells overexpressing p53: calpain involvement. Cell Death Differ
Leist M, Jaattela M (2001) Four deaths and a funeral: from caspases to alternative mechanisms. Nat Rev Mol Cell Biol 2(8):589–598
Acknowledgments
This work was financed by a grant from FONDECYT (1040800). C.L and I.A. are PhD fellows from the National Commission for Scientific Research and Technology (FONDECYT 1040800). We thank Mrs Connie Mefalle for her excellent assistance in English grammar.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lizama, C., Alfaro, I., Reyes, J.G. et al. Up-regulation of CD95 (Apo-1/Fas) is associated with spermatocyte apoptosis during the first round of spermatogenesis in the rat. Apoptosis 12, 499–512 (2007). https://doi.org/10.1007/s10495-006-0012-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10495-006-0012-1