
Abstract
The tick Dermacentor everestianus mainly distributed in Tibet, China and Nepal, and can transmit some pathogens causing great damages in Qinghai-Tibet Plateau. This study investigated the life cycle and development characteristics of D. everestianus under field conditions. The average duration of the whole life cycle of D. everestianus was 124.4 days, with the host available in the field plot. Under natural conditions, the mean feeding, preoviposition and oviposition period of female ticks were 6.1, 17.9 and 21.2 days, respectively. The incubation time of eggs was the longest phase in the life cycle of the ticks (26 days on average). Moreover, the weight of engorged females was highly positively correlated with the number of the eggs that were laid (r = 0.81, P < 0.05). The reproductive efficiency index and reproductive fitness index in females were 7.3 and 5.9, respectively. The above findings suggest that the tick D. everestianus have evolved well adaptability to the highland areas.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
As obligatory blood-feeding arthropods, ticks are important vectors of human and animal pathogens (Estrada-Peña and Jongejan 1999). Dermacentor everestianus Hirst is mainly distributed in Tibet, China and Nepal, and commonly dwells in alpine desert above an altitude of 4000 m in Tibet (Teng and Jiang 1991). Adult ticks mainly parasitize medium to large-sized mammals (sheep, blue sheep, yaks and horse), whereas larvae and nymphs mainly feed on rodents and lagomorphs (Teng and Jiang 1991). D. everestianus can transmit some pathogens including Francisella tularensis, Rickettsia raoultii and Anaplasma ovis, causing human health problems and economic losses in livestock industries in Qinghai–Tibet Plateau (Yu et al. 2015). Recently, the tick D. everestianus has been morphologically re-described (Apanaskevich et al. 2015), and regarded as taxonomic valid with D. abaensis and D. birulai being the junior synonyms (Chen et al. 2014). It is phylogenetically clustered with D. nuttalli and D. silvarum based on mitochondrial sequences (Chen et al. 2014; Yu et al. 2018).
Under laboratory conditions, the tick D. everestianus can complete one generation within 110.2 days (ranged 87–136 days) under laboratory conditions (Jin et al. 2017). However, little is known about the life cycle and biological characteristics of D. everestianus under field conditions, which is important for understanding its adaption to the hostile environments in high altitude areas, and further evaluating its potential influences on human health and livestock production. Hence, the current study investigated the life cycle and development characteristics of this tick species under field conditions in the Tibet Autonomous Region of China. The results will be helpful in accumulating knowledge on ecological adaptation of this tick to Tibet plateau, and providing references for the constructive control of D. everestianus and related tick-borne pathogens.
Materials and methods
Study site
The study was conducted from March 2014 to March 2015 in Damxung County (90°45′–91°31′E, 29°31′–31°04′N), north of Lhasa City located in the center of the Tibet Autonomous Region, southwest of China. A 150 × 150 cm field plot (90°51′737″E, 30°25′325″N, and altitude 4353 m) was chosen in a natural tick habitat to study the life cycle and biological characteristics of D. everestianus (Fig. 1). A 10 × 10 cm water-filled drain encircled the plot to prevent tick escape, and a wooden fence was enclosed to prevent other animals from entering. The plot was adjacent to the habitat of D. everestianus and covered with approximately 5 cm scattered bushes including Kobresia schoenoides and Sabina pingii.

Study site (red dot). (Color figure online)
During the study period, monthly minimum, maximum, and mean ambient temperatures, as well as relative humidity were obtained using hygrothermographs (Qingsheng Electronic Technology, China) that were planted in the plot and located 50 cm above the ground (Fig. 2).

Monthly minimum, maximum, and mean air temperature (T, °C) and relative humidity (RH, %) from March 2014 to March 2015, at the field plot in Tibet Autonomous Region, China. (Color figure online)
Observation of the biology of adult ticks
From March to early-April 2014, the free-living adult ticks of D. everestianus were collected by flag-dragging from the vegetation (K. schoenoides, S. pingii and Caragana spinifera), and weighed individually. These adult ticks (50 females and 50 males) were fed on the ears of white rabbits (Oryctolagus cuniculus domesticus) maintained in iron cages (40 × 40 × 50 cm) in the field plot as described previously (Liu et al. 2016), and monitored daily (9:00, 14:00 and 18:00). To facilitate the collection and observation of the ticks, one end of the cloth bag was tightly attached to rabbit ears by adhesive tape, and the other end of the bag was sealed to prevent the ticks from escaping. All the rabbits were fed with rabbit pellets and provided water ad libitum under natural lighting and climate conditions.
When fully engorged female ticks detached from rabbits, they were individually collected, weighed and placed in separate glass tubes (1.5 × 12 cm). The tubes were placed in the plot, covered with an approximately 4 cm thick leaf layer and closed using gauze netting with proper ventilation. They were examined twice daily to record the preoviposition periods, oviposition periods and the mass of the laid eggs. After the onset of oviposition, the deposited eggs were collected daily, weighted and put into glass tubes separately. In order to observe the incubation period, 1000 eggs were counted and placed in separate individual tubes. Preoviposition periods were defined as the number of days from detachment to the beginning of oviposition, and oviposition periods were defined as the number of days from the beginning to the end of oviposition. The reproductive efficiency index (REI) was calculated as the ratio of the number of eggs laid to the weight of the engorged female. The reproductive fitness index (RFI) was calculated as the ratio of the number of eggs hatched into larvae to the weight of engorged females (Drummond and Whetstone 1970; Chilton 1992).
Observation of the biology of larvae and nymph ticks
To assess the prefeeding periods, freshly hatched larvae (n = 200) and nymphs (n = 100) were placed on the ears of the rabbits with cloth bags and checked daily. Prefeeding periods were defined as the number of days from hatching or molting to the beginning of attachment. In order to determine the feeding periods, 100 unfed larvae and nymphs were weighted, put into cloth bags (6 × 4 × 15 cm) and checked daily during May and June 2014. To determine the premolting and molting periods, naturally detached engorged immature ticks were collected daily, weighed and placed individually in glass tubes closed with gauze netting. The glass tubes were covered with 4 cm thick leaf layer and placed in the plot under natural lighting and climate conditions.
Survival of free-living Dermacentor everestianus in the field plot
From May to July 2014, 100 newly hatched larvae and 100 newly emerged nymphs were placed in separate glass tubes and checked daily to determine their survival ability without feeding. In July 2014, one hundred newly molted adult ticks were confined in large glass tubes to evaluate the overwinter survival rate of the ticks. All glass tubes that were placed in the plot were covered with an approximately 4 cm thick leaf layer. Survival was determined after returning these adult ticks to the laboratory in March in the next year and incubating them for 24 h (25 ± 1 °C, 70% RH, 6-h light and 18-h dark).
Statistical analysis
All data were processed using Microsoft Excel 2010 (Microsoft, Redmond, WA, USA). The correlation between the number of eggs laid and the weight of engorged females was assessed by linear regression analysis using SPSS 20.0 (IBM, Armonk, NY, USA). All charts were prepared using GraphPad Prism (GraphPad software, La Jolla, CA, USA).
Results
Life cycle of Dermacentor everestianus under field conditions
The average duration of prefeeding, feeding, and preoviposition of adults were 4.9, 6.1, and 17.9 days, respectively. Oviposition occurred in early April, 2014. The duration of oviposition and the average incubation period of eggs were 21.2 days (range 17–31 days) and 26 days (range 24–29 days). Larvae first appeared in the confined plot in late April, 2014. The nymphs and adults appeared in mid-May and late June, 2014, and the mean developmental duration of larvae and nymphs was 17.1 days (range 14–20 days) and 31.2 days (range 26–38 days), respectively. Overall, these results indicate that D. everestianus requires an average of 124.4 days (range 105–150 days) to complete the whole life cycle from unfed adult to the next generation adult (between March and September, 2014) (Table 1), and the incubation period of eggs is the longest time during the development of D. everestianus (Table 1; Fig. 3).

Different developmental stage parameters during the life cycle of Dermacentor everestianus under field conditions. (Color figure online)
Oviposition of adult Dermacentor everestianus in field plot
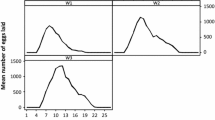
The mean period of oviposition of females lasted for 21.2 days (range 17–31 days), and the average number of eggs laid was 2237.5 (range 1287–2975) (Table 2). After the onset of oviposition, the number of eggs laid increased gradually during the first 4 days and reached the peak on the fifth day, and declined quickly afterward (Fig. 4). The REI and RFI were 7.3 and 5.9, respectively (Table 2). The hatchability of eggs was 81.2%. The number of eggs laid was positively correlated with the weight of engorged females (r = 0.81, P < 0.05) (Fig. 5).

Mean (± SEM) daily number of eggs deposited by females of Dermacentor everestianus (based on 15 females) under field conditions

Relationship between the weight of engorged females of Dermacentor everestianus and number of eggs laid (based on 15 females) under field conditions
Feeding behavior and survival of Dermacentor everestianus
The fresh larvae, nymphs and adults required 2.4, 3.5 and 4.9 days to attach to the hosts, and the engorgement periods were 2.9, 4.4 and 6.1 days, respectively. Both the nymphal and adult D. everestianus fed slowly on the first day after attachment, then engorged quickly and dropped in the following 2 days. In contrast, larvae fed quickly on the first day, and fully engorged on the second day. Copulation is the prerequisite for rapid engorgement of females. Compared with the body weight of unfed females, the mean body weight of the engorged females increased by 41.5-folds. However, the mean body weight of engorged males only increased by 1.3-folds when compared with that of unfed males. The average weight ratios of engorged larvae and nymphs to unfed ones were 11.3 and 26.3, respectively (Table 3).
From May 2014 to March 2015, 100 freshly hatched larvae and 100 freshly molted nymphs were tested for the survival period in the plot. The larvae and nymphs could survive 22–31 days and 36–50 days without feeding. In March 2015, the survival rate of adult ticks was 71.2%, indicating that the adults survived for about 9 months.
Discussion
The flexible life cycle strategies of arthropods reflect their adaptive response to variable environmental conditions (Visser et al. 2016). Due to the extremely hostile environments, Qinghai–Tibetan Plateau is regarded as a unique natural region unit in the word due to its extremely hostile environment, where the tick D. everestianus mainly dwells in alpine desert above altitude of 4000 m (Teng and Jiang 1991). The current study elucidated the life cycle of D. everestianus under nature conditions, which provides basic knowledge about the adaptation to high altitude areas of this tick species.
Under field condition, D. everestianus required an average of 124.4 days (range 105–150 days) to complete its life cycle, which is longer than the life cycle of the tick (110.2 days on average) observed under laboratory condition (22 ± 1 °C, 75 ± 5% RH, 16-h light and 8-h dark) (Jin et al. 2017). The prolonged life cycle under field condition was also observed in D. silvarum (Yu et al. 2010) and D. marginatus (Cao et al. 1991). The environment in Tibet plateau was featured by poor oxygen, drought and low temperature (Fig. 2). Such conditions could reduce the metabolic activity of the tick, leading to increased life cycle duration (Branagan et al. 1973).
In Tibet plateau, the incubation of eggs usually occurs in April, when the temperature and relative humidity start arising. The incubation period (26 days on average) was the longest developmental stage in the life cycle of D. everestianus. Similar results were also obtained in Amblyomma rotundatum (Rodrigues et al. 2010) and Hyalomma rufipes (Chen et al. 2012). The long incubation period of eggs will keep the emergence of larvae in late April and May which is consistent with the activity of hosts. In the field, the REI and RFI of D. everestianus were 7.3 and 5.9, respectively (Table 2), which are close to the values observed under laboratory condition (Jin et al. 2017). As the oviposition sites are usually covered with decaying vegetation, where the temperature and relative humidity are relatively stable (Estrada-Peña and de la Fuente 2014). The hatching rate of eggs in D. everestianus reached 81.2%, indicating that this tick species is adapted to the environment of Tibet plateau.
Diapause (behavioral and morphogenetic) is a genetically determined behavior and the induction and termination of diapause is mostly under photoperiodic control (Belozerov 1982; Oliver 1989). In our study, the adult D. everestianus that appeared in July 2014 did not start questing until the next March. Even when these ticks were artificially placed on the ears of rabbits, the females did not feed to engorgement. These findings suggest that unfed adult D. everestianus starts behavioral diapause in July, which can ensure them survive through winter. In contrast, the pre-oviposition period of females (17.9 days on average) was prolonged under field condition, which was not observed under standard laboratory condition (6.8 days on average) (Jin et al. 2017). This may be attributed to the complex interactions among photoperiod, temperature, relative humidity and other ecological factors (Frenot et al. 2001; Padgett and Lane 2001). Host availability may be another reason for prolonged pre-oviposition that can ensure the appearance of larvae in late April and May when the host is available.
When oviposition began, the number of egg mass of D. everestianus was low in the first 2–4 days, followed by a peak on the fifth day, and declined gradually afterward (Fig. 4). This trend of oviposition is mainly affected by the daily temperature and relative humidity (Despins 1992). Similar pattern was reported in D. everestianus under laboratory condition (Jin et al. 2017) and in H. tibetensis under field condition (Liu et al. 2016). The weight and number of egg masses laid are positively correlated with the weight of engorged female ticks (Snow and Arthur 1966). The linear positive correlation between the egg masses and the weight of engorged female (r = 0.81) was also observed in D. everestianus under field condition. The weight ratios of females, nymphs and larvae from unfed to engorgement were 41.5-, 26.3- and 11.3-fold, respectively (Table 3), which were similar to the values observed under laboratory condition (Jin et al. 2017).
Under field condition, the unfed larvae and nymphs of D. everestianus could survive 22–31 days and 36–50 days, respectively. The life span of unfed adults is about 9 months, and they can survive winter successfully, indicating that D. everestianus requires at least one year to complete one generation under field condition. Our findings provide a starting point for future investigation on seasonal population dynamics of the tick D. everestianus.
References
Apanaskevich DA, Duan W, Apanaskevich MA, Filippova NA, Chen J (2015) Redescription of Dermacentor everestianus Hirst (Acari: Ixodidae), a parasite of mammals in mountains of China and Nepal with synonymization of D. abaensis Teng and D. birulai Olenev. J Parasitol 100(3):268–278. https://doi.org/10.1645/13-369.1
Belozerov VN (1982) Diapause and biological rhythms in ticks. In: Obenchain FD, Galun R (eds) Physiology of ticks. Pergamon Press, Oxford, pp 469–500
Branagan D (1973) The developmental periods of the ixodid tick Rhipicephalus appendiculatus Neum. under laboratory conditions. Bull Entomol Res 63(1):155–168. https://doi.org/10.1017/S0007485300050951
Cao HL, Ye RY, Xiao H, Riziwan (1991) Observations on the Biology of Dermacentor marginatus (Acariformes: Ixodidae). Endem Dis Bull 3:117–122. (in Chinese)
Chen Z, Li Y, Liu Z, Yang J, Yin H (2012) The life cycle of Hyalomma rufipes (Acari: Ixodidae) under laboratory conditions. Exp Appl Acarol 56(1):85–92. https://doi.org/10.1007/s10493-011-9490-0
Chen Z, Li YQ, Ren QY, Luo J, Liu ZJ, Zhou X, Liu GY, Luo GX, Yin H (2014) Dermacentor everestianus Hirst, 1926 (Acari: Ixodidae): phylogenetic status inferred from molecular characteristics. Parasitol Res 113(10):3773–3779. https://doi.org/10.1007/s00436-014-4043-1
Chilton NB (1992) An index to assess the reproductive fitness of female ticks. Int J Parasitol 22(1):109–111. https://doi.org/10.1016/0020-7519(92)90086-Z
Despins JL (1992) Effects of temperature and humidity on ovipositional biologyand egg development of the tropical horse tick Dermacentor (Anocentor) nitens. J Med Entomol 29(2):332–337. https://doi.org/10.1093/jmedent/29.2.332
Drummond RQ, Whetstone TM (1970) Oviposition of the Gulf Coast tick. J Econ Entomol 63(5):1548–1551. https://doi.org/10.1093/jee/63.5.1547
Estrada-Pen˜a A, de la Fuente J (2014) The ecology of ticks and epidemiology of tick-borne viral diseases. Antivir Res 108:104–128. https://doi.org/10.1016/j.antiviral.2014.05.016
Estrada-Peña A, Jongejan F (1999) Ticks feeding on humans: a review of records on human-biting Ixodoidea with special reference to pathogen transmission. Exp Appl Acarol 23(9):685–715. https://doi.org/10.1023/A:1006241108739
Frenot Y, de Oliviera E, Gauthier-Clerc M, Deunff J, Bellido A, Vernon P (2001) Life cycle of the tick Ixodes uriae in penguin colonies: relationships with host breeding activity. Int J Parasitol 31(10):1040–1047. https://doi.org/10.1016/S0020-7519(01)00232-6
Jin S, Wang TH, Li T, Liu M, Li T, Yang XL, Wang H, Jia QY, Yu ZJ, Liu JZ (2017) Life cycle of Dermacentor everestianus Hirst, 1926 (Acari: Ixodidae) under laboratory conditions. Korean J Parasitol 55(2):193–196. https://doi.org/10.3347/kjp.2017.55.2.193
Liu M, Li T, Yu ZJ, Gao XH, Zuo CW, Wang RR, Li NX, Wang H, Liu JZ (2016) Characterization of the life cycle of the tick Haemaphysalis tibetensis under field conditions in Qinghai–Tibet plateau. Exp Appl Acarol 69(1):107–115. https://doi.org/10.1007/s10493-016-0020-y
Oliver JH (1989) Biology and systematics of ticks (Acari: Ixodidae). Annu Rev Ecol Syst 20:397–430. https://doi.org/10.1146/annurev.es.20.110189.002145
Padgett K, Lane RS (2001) Life cycle of Ixodes pacificus (Acari: Ixodidae): timing of developmental processes under field and laboratory conditions. J Med Entomol 38(5):684–693. https://doi.org/10.1603/0022-2585-38.5.684
Rodrigues DS, Maciel R, Cunha LM, Leite RC, de Oliveira PR (2010) Amblyomma rotundatum (Koch, 1844) (Acari: Ixodidae) two-host life-cycle on Viperidae snakes. Rev Bras Parasitol Vet 19(3):174. https://doi.org/10.1590/S1984-29612010000300009
Snow KR, Arthur DR (1966) Oviposition in Hyalomma anatolicum anatolicum (Koch 1844) (Acarina: Ixodidae). Parasitology 56(3):555–568
Teng GF, Jiang ZJ (1991) Economic insect fauna of China, Fasc 39 Acari: Ixodidae. Science Press, Beijing. (in Chinese)
Visser B, Lann CL, Snaas H, Verdeny-Vilalta O, Harvey JA (2016) Divergent life history strategies in congeneric hyperparasitoids. Evol Ecol 30(3):535–549. https://doi.org/10.1007/s10682-016-9819-6
Yu ZJ, Zheng HY, Chen Z, Zheng B, Liu JZ (2010) The life cycle and biological characteristics of Dermacentor silvarum Olenev (Acari: Ixodidae) under field conditions. Vet Parasitol 168(3):323–328. https://doi.org/10.1016/j.vetpar.2009.11.010
Yu ZJ, Wang H, Wang TH, Sun WY, Yang XL, Liu JZ (2015) Tick-borne pathogens and the vector potential of ticks in China. Parasit Vectors 8:24. https://doi.org/10.1186/s13071-014-0628-x
Yu ZJ, Zhang S, Wang TH, Yang X, Wang H, Liu JZ (2018) The mitochondrial genome and phylogenetic analysis of the tick Dermacentor everestianus hirst, 1926 (acari: ixodidae). Syst Appl Acarol 23(7):1313–1321. https://doi.org/10.11158/saa.23.7.8
Acknowledgements
This study was supported by National Natural Science Foundation of China (31672365), and Youth Foundation of Department of Education of Hebei Province (QN2018027).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Li, T., Liu, M., Zhang, TT. et al. The life cycle and development characteristics of Dermacentor everestianus (Acari: Ixodidae) under field conditions in Qinghai–Tibet Plateau. Exp Appl Acarol 76, 513–522 (2018). https://doi.org/10.1007/s10493-018-0325-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-018-0325-0