
Abstract
Livestock production in the tropics contributes significantly to global greenhouse gas emissions, so better understanding the role of silvopastoral systems (SPS) in mitigating such emissions is necessary. The aim of this study was to evaluate the amounts of carbon stored in the biomass and soil organic carbon (SOC) components of a Leucaena leucocephala cum Panicum maximum silvopasture system (SPS) compared to a deciduous tropical forest (DTF), and a grass monoculture (GM) in Michoacán, Mexico. The above- and below-ground biomass were measured by destructive sampling in the SPS and GM, while previously reported allometric equations were used to quantify biomass stocks in the DTF. The SOC concentration up to 30 cm was determined by dry combustion method. The SPS and DTF contained more aboveground biomass (41.8 ± 3.30 and 36.7 ± 5.72 Mg DM ha−1) compared to GM (8.0 ± 0.76 Mg DM ha−1). However, the SPS exhibited greater belowground biomass (16.4 ± 1.95 Mg DM ha−1) than the other systems. The DTF had the highest SOC fraction in all depth classes with values ranging from 3.1 ± 0.07% to 3.7 ± 0.06%, respectively, compared to the other systems. The total carbon stocks in SPS was similar to DTF (120.7 ± 10.97 vs. 120.9 ± 6.38 Mg C ha−1) but was significantly higher than GM (78.2 ± 8.41 Mg C ha−1). In dry tropical conditions, SPS displays enormous potential for increasing biomass and soil carbon stocks compared to the GM and can thus be used as a greenhouse gas mitigation strategy in livestock production systems.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Global climate change, caused by rising levels of carbon dioxide (CO2) and other greenhouse gases (GHG) in the atmosphere, and food security are recognized as the most serious problems of human society of this century (Feliciano et al. 2018). Worldwide, annual CO2 emissions into the atmosphere from fossil fuel combustion and cement production reached 9.5 Pg C yr−1 in 2011, and land use, particularly deforestation to initially establish monoculture farming and the subsequent planting of pasture for livestock (Feliciano et al. 2013; Villanueva-López et al. 2016), accounted for 0.9 Pg C yr−1 of net CO2 emissions (IPCC 2013). Extensive pastureland for animal grazing is the dominant land use in many parts of the tropics with almost 80% of forest area cleared to establish pasture (McGroddy et al. 2015). While the global net carbon uptake of terrestrial ecosystems was 60 Pg C yr−1, only 1 Pg C yr−1 is considered long-term carbon storage, so the role of land use systems in stabilizing CO2 levels and reducing the carbon footprint of existing production practices has attracted considerable scientific attention as a fundamental approach to mitigate global climate change (Ibrahim et al. 2007; Nair 2012). Due to the fast increase in pastureland area, any strategy that reduce GHG emissions or increase in carbon storage in these ecosystems would have a large impact on the carbon balance at a regional to national level (McGroddy et al. 2015).
On a global level, tropical forests are the ecosystems with the greatest net primary productivity, they play an important role not only in regulating the global climate by storing significant quantities of CO2 and other greenhouse gases (Jose 2009), but also, they offer a diversity of foods (foliages and fruits) that allow the animals to vary their diet and increase their level of production particularly in regions of the dry tropics with scarce forages. The tropical forest in Mexico spans 79.29 million hectares, of which 2.68 million hectares are low, seasonally dry tropical forest (INEGI 2005). In the state of Michoacán (southwest Mexico), low deciduous tropical forests (DTFs) are characterized by the presence of three well-defined strata: the canopy, shrub, and herbaceous levels. The canopy has an average height of only 3 m, with the dominant species belonging to the Burseraceae and Fabaceae families. However, these forests have experienced high rates of deforestation and degradation, mainly due to land use conversion to monoculture pastures for ruminant livestock, which affects the capacity of these ecosystems to maintain their functions and provide environmental services. Additionally, degradation of more than 65% of the pasture is contributing to the increase in atmospheric CO2 (Ibrahim et al. 2007).
In the search for strategies to increase carbon reservoirs in livestock production systems of the tropical region, and promote participatory adoption and adaptation strategies by small producers, silvopastoral systems (SPS) have been proposed as an alternative to monoculture pastures that will allow for sustainable production and, at the same time, produce direct economic benefits for farmers (Tapia-Coral et al. 2005). These livestock production systems integrate the management of trees and/or shrubs with that of grasses and animals to promote a series of ecological interactions as well as create economic differences between trees and other system components (Broom et al. 2013; Casanova-Lugo et al. 2014). These systems offer numerous benefits, including the protection of the soil from erosion and the introduction of greater numbers of trees and shrubs, thereby increasing the quantity of organic material (Partey 2011) and consequently improving soil properties.
The incorporation of shrubs and trees to open grazing systems increases the above and belowground biomass as well as other nutrients to the soil. Higher soil organic carbon can increase soil water holding capacity and improve other soil properties like cation exchange capacity, porosity and infiltration (Cubillos et al. 2016). Incorporation of leguminous species such as Leucaena leucocephala (Lam) de Wit., enhance symbiotic nitrogen fixation that provide a limiting nutrient for the fodder plants and reduce the burden of purchasing synthetic nitrogen fertilizers (Cubillos et al. 2016).
In addition, high-quality food is available for livestock throughout the year (Broom et al. 2013; Feliciano et al. 2018), which contributes to reducing deforestation in tropical regions and the concentration of greenhouse gases in the atmosphere (Matos et al. 2012; Mbow et al. 2014; Luedeling et al. 2014). Furthermore, the micro-environmental conditions in these systems are improved due to an increase in shaded areas (Elevitch 2006), and greater animal and plant biodiversity is maintained (Jose 2012; Broom et al. 2013). In recent years, SPS have also been recognized for their potential to store large quantities of carbon (Jose 2009; Lorenz and Lal 2014) and atmospheric nitrogen in their biomass and soil (Munroe and Isaac 2014) compared to crop and pasture monocultures (Bambrick et al. 2010; Djomo et al. 2011; Feliciano et al. 2018), which is due to the rapid growth and high productivity of these systems. In developing countries particulary in tropical regions, as is the case of Mexico this storage represents an economic opportunity for small farmers if opportunities to sell carbon sequestered through agroforestry systems to industrialised countries become more widespread (Feliciano et al. 2018), and above all, it contributes increase yield and to the resilience of livestock production systems (Hoosbeek et al. 2016; Coulibaly et al. 2017; Waldron et al. 2017). FAO considered silvopastoral system as one of the climate-smart agricultural practices that provides economic diversity and protection from erosion in addition to carbon uptake benefits (Harvey et al. 2014).
It was recently reported that approximately 10,000 hectares of SPS have been established in the states of Michoacán, Campeche, San Luis Potosi, Veracruz, Tamaulipas, Chiapas, Nayarit, Quintana Roo, and Yucatan, Mexico (Broom et al. 2013). These systems often involve rotational grazing and the use of electric fences in pastures cultivated with the Cunningham cultivar of Leucaena leucocephala in addition to a diverse set of tropical grasses, such as the Tanzania cultivar of Panicum maximum Jacq. and Cynodon plectostachyus (K. Schum.) Pilg. However, despite the widespread use of these systems in the Mexican tropics relative to other systems, little is known about the diverse environmental services they offer, such as carbon storage. Better understanding about the effects of trees and shrubs on carbon storage and soil nutrient dynamics of pasturelands will help improve the theoretical basis for agroecosystem management that guides to harmonizing food production systems with environmental and biodiversity conservation (Casals et al. 2014). Therefore, the general objective of this study was to determine the role of SPS in biomass and soil carbon storage in comparison to a DTF and a C. plectostachyus grass monoculture (GM) in Michoacán, Mexico.
Materials and methods
Study area
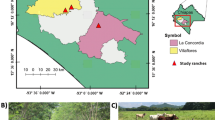
The study was carried out at La Concha in the municipality of Apatzingán, Michoacán, Mexico (Fig. 1); located at 19°08″N, 102°35″W at an elevation of 325 masl. The climate is dry with a summer rainfall pattern, an average annual rainfall of 924 mm, and an average temperature of 28.3 °C (data taken from a local meteorological station, Fig. 2). The soils are predominantly vertisols, based on the classification of the World Reference Base for Soil Resources (WRB 2007), and they have a pH of 7.2–7.7, a soil bulk density (BD) of 0.98–1.37 g cm−3, a total nitrogen content ranging from 1.5 to 1.7 mg N g−1, available phosphorus content from 0.05 to 0.06 mg P g−1, a potassium content from 5.2 to 5.8 mg K g−1, and an organic carbon content from 21.10 to 33.40 mg C g−1.

Location of the study area: a location of state Michoacán in the Southwest of Mexico; b tropical dry region in the Municipality of Apatzingán; c deciduous tropical forest; d silvopastoral system; e grass monoculture

Monthly variation in temperature and rainfall pattern at Apatzingán, Michoacán, Mexico
System descriptions
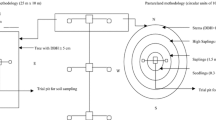
Three ecosystems were selected: a deciduous tropical forest (DTF), a grass monoculture (GM), and a silvopasture system (SPS), and we established five 100 m2 (10 m × 10 m) carbon monitoring plots in each of these systems. Table 1 presents the biophysical characteristics of the ecosystems. The low DTF represents the regional native vegetation and has been regenerating for approximately 20 years old. During the study, the average tree density was 557 individuals per ha with 17–32 tree species per site. The low DTF trees with the highest relative importance values include Cordia elaeagnoides (A. DC.), Apoplanesia paniculata (C. Presl), Randia capitata (DC.), Caesalpinia platyloba (S. Watson), Capparis asperifolia (C. Presl), and Simira mexicana (Bullock Steyerm), which the local communities use for various purposes including fodder, firewood, timber, tools, medicine, pollinators attraction, fruit production, and as living fences and posts.
The GM consists of African star grass (C. plectostachyus) that was planted in 2005 by scattering seed. Management consists of a 5-day occupation period followed by 40–65 days of rest throughout the year with a stocking rate of 2 animal units/ha. During the study period, gravity-fed irrigation was applied every 15 days during the dry season and suspended during the rainy season. In addition, organic fertilizer (bovine manure) was applied at a rate of 100 kg ha−1 yr−1 every six months, and no weed control was performed.
The SPS consists of six-year-old Leucaena leucocephala cv. Cunningham as tree component and Panicum. maximum cv. Tanzania grasses, mixed in an area that was previously a more than 15 years old GM. The distance between the rows of L. leucocephala was 1.60 m, and the distance between plants was 0.20 m, yielding a density of approximately 36,000 plants per ha. The gaps were planted by scattering P. maximum cv. Tanzania seeds. This system is grazed to fatten cattle to produce approximately 350 to 450-kg animals (stocking rate of 4.0 animal units per ha). The SPS is grazed for 3 days and allowed to rest for 35 to 45 days in the rainy and dry seasons, respectively. Gravity-fed irrigation was applied every 15 days during the dry season.
Biomass estimation
Trees in the DTF sampling units were identified using field guides and dichotomous keys. The diameter at breast height (DBH) was measured for all trees with a DBH exceeding 10 cm, and tree biomass (TB) was indirectly estimated using an allometric equation Eq. (1) developed by Chave et al. (2005). This equation was developed by using a compilation of database from a wide range of tropical trees, including tropical dry forests of Mexico. The equation includes small as well as big trees ranging from 5 to 138 cm DBH (Chave et al. 2005). The tree DBH data from our study lie within the range of the data used to generate the equation. In addition, the use of species specific wood density data along with DBH and tree height, increases the applicability of the equation to a wide range of tropical ecosystems. The wood density data of the species were obtained from published reports (Reyes et al. 1992).
where AGB is the above-ground biomass (Mg DM ha−1); ρ is the specific density (g cm−3); DBH (cm) is measured at a height of 1.37 m; and H is the total height of the individual tree (m).
Root biomass was estimated using an allometric equation Eq. (2) developed by Cairns et al. (1997) that has been widely used by several authors (Nakakaawa et al. 2009; Soto-Pinto et al. 2010; Schmitt-Harsh et al. 2012) and is an accepted method in the land use manual by the IPCC (2007):
where BGB is the below-ground (root) biomass (Mg DM ha−1), and AGB is the above-ground biomass (Mg DM ha−1).
To estimate the herbaceous biomass in the DTF, grass biomass of a specific area was estimated using the square frame sampling method. In each sampling unit, three samples were collected from a 1 m × 1 m area marked with a square PVC frame in the dry season. The biomass in the square was cut at ground level and weighed in its fresh state in the field, and at the same time, the leaf litter was collected and weighed. Afterward, the material was dried in a forced-air-circulation oven at 60 °C for 48 h to determine the amount of dry matter (DM).
Destructive sampling was performed to determine the tree biomass in the SPS. Fifteen L. leucocephala plants were randomly selected (three in each permanent sampling plot) and carefully extracted from the soil manually, including the roots, as described by Casanova-Lugo et al. (2010). Afterward, the plants were separated into leaves, branches, stems, and roots, and each part was weighed in its fresh state in the field. Then, samples weighing approximately 300 g were collected from each of these components and dried in a forced-air-circulation oven at 60 °C to determine the amounts of DM.
To examine productivity, the herbaceous biomass and leaf litter in the SPS and GM systems were sampled using a square frame. Six sampling units were marked with the 1 m × 1 m square PVC frame, and the grass within the square was cut at ground level and weighed. Afterward, the samples were separated into leaves, stems, and dry material. The leaf litter within the square was also collected and weighed.
The grass root biomass was extracted by excavation inside the square (Casanova-Lugo et al. 2010) to a depth of 30 cm, and the roots were separated from the soil by pressure washing with water. The roots were then dried in a forced-air-circulation oven at 60 °C until they attained a constant weight; the final weights were recorded and extrapolated to surface area units. As the carbon content is commonly assumed to be 45–50% of the biomass (Brown 2002), all biomass estimates were multiplied by 0.5, and this value was multiplied by the planting density of the respective systems.
Soil organic carbon concentration
Soil organic carbon (SOC) was measured by excavating and sampling three pits measuring 0.5 m × 0.5 m across and 30 cm deep, and soil samples were obtained from three depth intervals: 0–10, 10–20, and 20–30 cm. An undisturbed sample was collected from each depth interval in a metal cylinder with a volume of 502.6 cm3, and these samples were transferred to the laboratory, where they were oven dried to 105 °C for 72 h and then weighed to determine the soil bulk density obtained using the cylinder method. The SOC concentration (%) was determined by the dry combustion method using a Leco CNS® element analyzer (Leco Corporation St. Joseph, MI, US), and the SOC in each depth interval was calculated using equation Eq. (3), as proposed by Xu et al. (2011):
where PC is the percentage of carbon; BD is the soil bulk density (g cm−3); and SD is the sampling depth (m).
Carbon storage in each system
The total carbon stocks associated with each land use was determined by using equation Eq. (4), which several authors (Nakakaawa et al. 2009; Soto-Pinto et al. 2010; Schmitt-Harsh et al. 2012) have applied to various tropical agro-ecosystems:
where AGB is the carbon stored in the above-ground biomass; BGB is the carbon stored in the below-ground biomass; Lit is the ground litter; and SOC is the soil organic carbon.
Finally, the rate of carbon fixation Mg C ha−1 yr−1 (mean annual biomass increment) was determined based on the biomass carbon density divided by the age of each system.
Statistical analysis
The carbon storage data were analyzed by one-way ANOVA using PROC GLM in SAS (SAS Institute Inc 2004), and two-way ANOVA was used to analyze the effects of the system, the soil depth, and the interaction between these factors on the SOC. Data were log-transformed to comply with the assumptions of normality and homoscedasticity of residuals when necessary. In all cases, when significant differences were noted between the carbon reservoirs of the systems, Tukey’s test was performed at a 5% level of significance to identify type I errors.
Results
Biomass estimation
We found that the SPS and DTF contained more aboveground biomass (41.8 ± 3.30 and 36.7 ± 5.72 Mg DM ha−1, respectively) than the GM (8.0 ± 0.76 Mg DM ha−1) (F = 41.91; P = 0.001) in Michoacán, Mexico (Table 2). The SPS exhibited greater belowground biomass (16.4 ± 1.95 Mg DM ha−1) than the other systems (F = 32.82; P = 0.001). The same pattern was noted for total biomass; the SPS had the greatest quantity of biomass (58.2 ± 4.77 Mg DM ha−1) during the evaluation period (F = 45.31; P = 0.001; Table 2). Furthermore, the aboveground biomass of the SPS represented 71.9% of the total biomass, and its root biomass represented 28.1% of the total. In contrast, the aboveground biomass of the DTF and GM systems represented 80.6 and 85.3% of the total, respectively, and the root biomass represented 19.4 and 14.7%, respectively.
The aboveground tree component represent 77% of the total carbon in TDF (Fig. 3), followed by roots (19%), herbaceous plants (2%) and ground litter (2%). In the case of the silvopastoral system, herbaceous plants represent the highest proportion of total carbon (33%), followed by trees/shrubs (31%), roots (28%) and the ground litter (9%). In contrast, the herbaceous components had the highest proportion of total carbon (59%) in the grass monoculture system, followed by the ground litter (27%) and roots biomass (15%).

Proportions of different components to the total biomass for each of the systems: silvopasture, grass monoculture and tropical deciduous forests in Michoacán, Mexico
Soil organic carbon
The SOC of the different depth classes, 0–10, 10–20, and 20–30 cm, differed significantly between the evaluated systems (F = 1.59; P = 0.021). Compared to the other systems, the DTF had the highest SOC fraction in all depth classes with values ranging from 3.1 ± 0.07 to 3.7 ± 0.06% respectively (Fig. 4).

Soil organic carbon fractions (%) at various depths in three agroecosystems in the municipality of Apatzingán, Michoacán. SPS, silvopastoral system; GM, grass monoculture; DTF, deciduous tropical forest
Soil bulk density
There was a statistically significant increase in BD with soil depth (F = 5.54; P = 0.004). In the 0–10-cm layer, the GM had the highest BD with a value of 1.39 34 ± 0.05 g cm−3. In the 10–20-cm layer, the higher BD values were observed in GM compared to SPS and DTF (1.40 ± 0.10 and 1.18 ± 0.04 g cm−3, respectively). In the 20–30-cm layer, BD was higher in the SPS and GM (1.31 ± 0.02 and 1.26 ± 0.11 g cm−3, respectively) compared with the DTF, with a value of 1.06 g cm−3 (Fig. 5).

Soil bulk density at various depths in the agroecosystems in the municipality of Apatzingán, Michoacán. SPS, silvopastoral system; GM, grass monoculture; DTF, deciduous tropical forest
Total carbon storage
The results indicated that carbon storage in the biomass (above- and below-ground) differed significantly among agro-ecosystems (one-way ANOVA, F = 16.41; P = 0.004), with carbon storage being higher in the SPS and DTF (29.1 ± 4.45 and 22.6 ± 3.48 Mg C ha−1, respectively) than in the GM (4.7 ± 0.62 Mg C ha−1). Furthermore, the SPS and DTF contained statistically similar amounts of SOC (91.6 ± 4.92 and 98.3 ± 2.90 Mg C ha−1, respectively), which were greater than that in the GM (73.4 ± 4.58 Mg C ha−1) (F = 9.25; P = 0.001; Table 3). Therefore, the SPS and DTF systems stored greater amounts of carbon than the GM in total (F = 7.02; P = 0.027). The mean annual biomass increment was significantly higher in the SPS (4.8 ± 0.74 Mg C ha−1 yr−1) compared to the DTF (1.5 ± 0.23 Mg C ha−1 yr−1) and GM (0.5 ± 0.06 Mg C ha−1 yr−1) (F = 70.17; P < 0.001; Table 3). However, this calculation is not considered as an indicator of carbon accumulation in GM because it can vary due to the seasonality of the herbaceous biomass.
Discussion
Biomass productivity in the three ecosystems
The mean annual biomass increment in our study was higher in the SPS compared to the DTF and GM, which is consistent with the results of several other studies that found greater productivity in tropical SPS compared to other land uses. In certain contexts, the productivity of tropical SPS can equal or exceed the productivity of natural ecosystems (Trumbmore et al. 1995; Montagnini and Nair 2004; Nair 2012). The annual carbon removal rate of 4.8 Mg DM ha−1 can significantly help livestock farmers to mitigate their net greenhouse gas emissions at farm level. This value is two hundred percent higher than the adjacent tropical forests and shows a huge potential to improve the largescale grass monoculture based livestock system in Michoacán and other parts of the tropics. The analyses indicated that the tree biomass stock of the DTF was nearly twice that of the SPS. Although the silvopastoral systems store a lower tree biomass compared to the adjacent forests, they have other advantages because we found that they promoted a greater amount of herbaceous biomass accumulation and increased the roots as well as litter carbon accumulation. The SPS stored almost four times the herbaceous biomass of the GM and almost 20 times that of the DTF. In addition, the root biomass and leaf litter of the SPS were higher than that of the GM and DTF. In this study, we did not measure the carbon input through annual litter production and fine root turnover, which we expect to increase with incorporation of trees and shrubs. On the other hand, the silvopastoral systems could increase the quality of the forage biomass since the inclusion of L. leucocephala improves the protein contribution to the animals, given that the latter is a limiting element in conventional livestock systems. In the same way, the silvopastoral systems could favor the physicochemical characteristics of the soil because they have a higher root biomass, which could reduce the soil compaction caused by the trampling of the animals, as well as promote a greater recycling of nutrients through the senescence of fine roots.
A study by Bacab and Solorio (2011) in Tepalcatepec, Michoacán, demonstrated that the biomass produced by a SPS consisting of L. leucocephala and P. maximum exceeded that of a GM (P. maximum), which is consistent with the results of the present study; however, in this case, the biomass values were inferior. These differences in biomass production may be attributed to various factors such as the type of management in each system (i.e., grazing and the frequency of irrigation), the species, the age of the system, and the season (Montagnini and Nair 2004). Furthermore, although the below-ground contribution of the plants (roots) was not reported, Bacab and Solorio (2011) found that the median value of 16.3 Mg DM ha−1 represented approximately 30% of the total biomass of the system, which is close to that found in this study (28.1%). This is also consistent with the findings of Casanova-Lugo et al. (2010) in Yucatan Peninsula, Mexico, who showed that the root biomass of a SPS can represent 10–40% of the total biomass of the system. In a SPS with Gliricidia sepium (Jacq. Kunth ex Walp.) trees and Brachiaria brizantha (Hochst. ex A. Rich.) grass in the humid tropic of Tabasco, Villanueva-López et al. (2015) also reported that tree roots represent more than 20% of the total biomass. However, the roots of woody species in semiarid regions can represent up to 60% of the total biomass; the low availability of soil moisture promotes the growth of roots to search for water and other nutrients (Mun and Whitford 1997). Moreover, the presence of trees in the pastures is likely to enhance soil C sequestration not only in the topsoil, but also in the subsoil due to bioturbation and turnover of the fine roots as reported by Haile et al. (2010) and Casals et al. (2014).
Soil organic carbon
The higher amount of SOC in the DTF and SPS than in the GM is consistent with the results of several studies that found greater amounts of soil organic material in DTF than in land devoted to other uses (Matamala et al. 2003). In this regard, Feliciano et al. (2018) notes that carbon benefits are greater in agroforestry systems tropical climates when compared to agroforestry systems located in other climates, both in terms of soil (2.23 Mg C ha−1 yr−1) and above ground (4.85 Mg C ha−1 yr−1). In terms of land use change, the greatest above ground carbon sequestration (12.8 Mg C ha−1 yr−1) occurs when degraded land is replaced by improved fallow and the greatest soil carbon sequestration (4.38 Mg C ha−1 yr−1) results from the transition of a grassland system to a silvopastoral system. Similarly, the presence of trees and shrubs in SPS increases the turnover of fine roots in the deeper soil horizons compared to GMs, which increase the total SOC stocks. However, several studies demonstrated a clear decrease in SOC with depth due to the effects of roots in the surficial soil (depth of 0–30 cm); a higher fine-root density contributed to a greater and faster rate of nutrient recycling (Tapia-Coral et al. 2005). However, the vegetation type also exerts a significant effect on the quantity and type of nutrients recycled by fine roots (Munroe and Isaac 2014), and the variations in SOC can also be directly linked to site-specific edaphic conditions, including the soil texture, land use history, age of the system, structure of the agroecosystem, inputs and outputs of organic material, and capacity of the soil to retain nutrients (Aryal et al. 2015).
Soil bulk density
The higher BD in the GM in our study could be attributed to the shallow root system of the grasses; in contrast, tree roots extend farther laterally and vertically, which facilitates greater nutrient and water uptake, thereby improving the physical structure of the soil (Matamala et al. 2003). In addition, the higher soil BD in the GM can be explained by soil trampling due to extensive livestock ranching. Furthermore, the relatively low fine-root turnover and absence of trees can alter the soil physico-chemical and biological properties, resulting in more compact soil in the GM (Manning et al. 2006; Villanueva-López et al. 2014). Carter (2002) reported that maintaining adequate levels of organic material helped decrease the soil BD and resistance to soil compaction. Our results are similar to those reported by Matos et al. (2012) who found soil BD values of 1.3 and 1.5 g cm−3 for the soils at depths of 0–10 and 10–20 cm, respectively, in an SPS of apple trees associated with Dactylis glomerata (L.) grass and values of 1.4 and 1.7 g cm−3 in a GM. However, Ibrahim et al. (2007) reported that the soil BD was lower in improved Brachiaria humidicola (Rendle) Schweickerdt grassland due to the rapid incorporation of organic material into the soil, which could contribute to the improvement of the soil profile and lead to a greater concentration of organic material. However, poor grazing management could result in a high concentration of livestock per unit area and a consequent increase in the degree of soil compaction in a GM, where the absence of trees slows the rate of soil recovery.
Carbon storage
The greater carbon storage in the SPS and DTF than in the GM can be attributed to the presence of trees and shrubs in the first two agroecosystems and their larger quantities of woody material; this conclusion was also reached by previous authors (Montagnini and Nair 2004; Amézquita et al. 2010; Nair 2012). Carbon storage can be strongly controlled by the composition and structure of the vegetation within the ecosystem (Aryal et al. 2014; Mattsson et al. 2015). For example, leguminous trees and shrubs improve the accumulation of organic material in the soil due to their ability to fix high quantities of atmospheric nitrogen, such as L. leucocephala, which fixes as much as 550 kg N ha−1 yr−1 (Leblanc et al. 2007; Munroe and Isaac 2014).
In a study performed in Colombia by Amézquita et al. (2010), a tropical forest and an SPS consisting of L. leucocephala and C. plectostachyus both stored high total amounts of carbon (134 and 138 Mg C ha−1, respectively), which significantly exceeded the total in a degraded pasture (97.0 Mg C ha−1). Likewise, Andrade et al. (2008) found that in an SPS with the trees Pithecellobium saman (Jacq.) Benth., Diphysa robinioides (Benth. ex Benth. & Oerst), and Dalbergia retusa (Hemsl.) in association with B. brizantha and Hyparrhenia rufa (Nees) Stapf pastures, the carbon in the above- and below-ground phytomass varied between 3.5 and 12.5 Mg C ha−1 in treeless pasture controls and the SPS, respectively. The total SOC in the upper 0.6 m in the SPS averaged 110 Mg C ha−1, which was approximately 38% higher than that in the GM. B. brizantha appeared to stimulate the production of tree roots, which, in turn, was highly correlated with total SOC, resulting in annual increases in total SOC of up to 9.9 Mg C ha−1. In a study of an SPS consisting of L. leucocephala and Andropogon gayanus Kunth., Miranda et al. (2008) reported carbon storage values of 64.4 and 38.3 Mg C ha−1, respectively, for a total of 102.7 Mg C ha−1. However, in this case, the carbon storage values were less than the present study, which was probably due to the differences in the edaphic and climate conditions and the ages of the systems. Together, these results suggest that native tropical forests and SPS, if well managed, can be important in the recovery of degraded pasture due to their high potential to capture atmospheric carbon.
Other researchers also agree that the benefits of SPS are more pronounced when tree and shrub densities are greater than 10,000 individuals per ha (Nair 2012; Broom et al. 2013; Casanova-Lugo et al. 2014). In addition to incorporating leaf litter and dead roots into the soil matrix, these systems promote greater soil nitrogen availability, which increases their capacity to store carbon for relatively short periods (Matamala et al. 2003). However, this aspect of SPS has not been studied extensively given that most nitrogen decomposition and liberation coefficients associated with the leaf litter and fine roots of leguminous trees with silvopastoral potential are unknown, particularly in seasonally dry tropical areas with intensive management regimes.
Conclusions
Silvopastoral systems can significantly increase biomass productivity and consequently improve carbon storage in the biomass and soil, thus allowing for more environmentally sustainable livestock production in dry tropical environments. The results of this study showed that the silvopastoral systems can store up to eight times more carbon in short time, compared to the livestock systems in pasture monocultures. SPS stored similar amounts of carbon in their biomass and soil as dry tropical forests. The mean annual carbon accumulation rates in the silvopasture systems were higher than that of grass monoculture and the adjacent deciduous tropical forests, giving insights into net greenhouse gas mitigation potential at the farm level. In addition to greenhouse gas mitigation, the establishment of SPS increase forage production and reduce natural resources degradation. However, it is necessary to conduct longer-term study on carbon and other nutrient dynamics of the silvopastoral systems including their fluxes from one pool to another to assist farmers and policy makers in designing sustainable livestock production systems.
References
Amézquita MC, Murgueitio E, Ibrahim M, Ramírez B (2010) Carbon sequestration in pasture and silvopastoral systems compared with native forests in ecosystems of tropical America. Grassland carbon sequestration: management, policy and economics. In: Abberton M, Conant R, Batello C (eds) Proceedings of the workshop on the role of grassland carbon sequestration in the mitigation of climate change, Roma, pp 153–155. http://www.fao.org/docrep/013/i1880e/i1880e06.pdf. Accessed 3 May 2014
Andrade HJ, Brook R, Ibrahim M (2008) Growth, production and carbon sequestration of silvopastoral systems with native timber species in the dry lowlands of Costa Rica. Plant Soil 308:11–22. https://doi.org/10.1007/s11104-008-9600-x
Aryal DR, De Jong BHJ, Ochoa-Gaona S, Esparza-Olguin L, Mendoza-Vega J (2014) Carbon stocks and changes in tropical secondary forests of southern Mexico. Agr Ecosyst Environ 195:220–230. https://doi.org/10.1016/j.agee.2014.06.005
Aryal DR, De Jong BHJ, Ochoa-Gaona S, Mendoza-Vega J, Esparza-Olguin L (2015) Successional and seasonal variation of litterfall and associated nutrient transfer in semi-evergreen tropical secondary forests of SE Mexico. Nutr Cycl Agroecosyst 103(1):45–60. https://doi.org/10.1007/s10705-015-9719-0
Bacab PHM, Solorio SFJ (2011) Forage offer and intake and milk production in dual purpose cattle managed under silvopastoral systems in Tepalcatepec, Michoacan. Trop Subtrop Agroecosyst 13(3):271–278. www.revista.ccba.uady.mx/ojs/index.php/TSA/article/view/1367
Bambrick AD, Whalen JK, Bradley RL, Cogliastro A, Gordon AM, Olivier A, Thevathasan NV (2010) Spatial heterogeneity of soil organic carbon in tree-based intercropping systems in Quebec and Ontario, Canada. Agrofor Syst 79:343–353. https://doi.org/10.1007/s10457-010-9305-z
Broom DM, Galindo FA, Murgueitio E (2013) Sustainable, efficient livestock production with high biodiversity and good welfare for animals. Proc R Soc B 280:2013–2025. https://doi.org/10.1098/rspb.2013.2025
Brown S (2002) Measuring carbon in forests: current status and future challenges. Environ Pollut 116:363–372. https://doi.org/10.1016/s0269-7491(01)00212-3
Cairns MA, Brown S, Helmer EH, Baumgarder GA (1997) Root biomass allocation in the world’s upland forests. Oecologia 111:1–11. https://doi.org/10.1007/s004420050201
Carter MR (2002) Soil quality for sustainable land management: organic matter and aggregation interactions that maintain soil functions. Agron J 94:38–47. https://doi.org/10.2134/agronj2002.3800
Casals P, Romero J, Rusch GM, Ibrahim M (2014) Soil organic C and nutrient contents under trees with different functional characteristics in seasonally dry tropical silvopastures. Plant Soil 374:643–659. https://doi.org/10.1007/s11104-013-1884-9
Casanova-Lugo F, Caamal-Maldonado J, Petit-Aldana J, Solorio-Sánchez F, Castillo-Caamal J (2010) Carbon storage in biomass of Leucaena leucocephala and Guazuma ulmifolia both in intercropping and monoculture systems. Revista Forestal Venezolana 54(1):45–50. http://www.saber.ula.ve/bitstream/123456789/31644/1/articulo4.pdf
Casanova-Lugo F, Petit-Aldana J, Solorio-Sánchez F, Parsons D, Ramírez-Avilés L (2014) Forage yield and quality of Leucaena leucocephala and Guazuma ulmifolia in mixed and pure fodder banks systems in Yucatan, Mexico. Agrofor Syst 88:29–39. https://doi.org/10.1007/s10457-013-9652-7
Chave J, Andalo C, Brown S, Cairns MA, Chambers JQ, Eamus D, Fölster H, Fromard F, Higuchi N, Kira T, Lescure JP, Nelson BW, Ogawa H, Puig H, Riéra B, Yamakura T (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145:87–99. https://doi.org/10.1007/s00442-005-0100-x
Coulibaly JY, Chiputwa B, Nakelse T, Kundhlande G (2017) Adoption of agroforestry and the impact on household food security among farmers in Malawi. Agric Syst 155:52–69. https://doi.org/10.1016/j.agsy.2017.03.017
Cubillos AM, Vallejo VE, Arbeli Z, Terán W, Dick RP, Molina CH, Molina E, Roldan F (2016) Effect of the conversion of conventional pasture to intensive silvopastoral systems on edaphic bacterial and ammonia oxidizer communities in Colombia. Eur J Soil Biol 72:42–50. https://doi.org/10.1016/j.ejsobi.2015.12.003
Djomo AN, Knohl A, Gravenhorst G (2011) Estimations of total ecosystem carbon pools distribution and carbon biomass current annual increment of a moist tropical forest. For Ecol Manag 261:1448–1459. https://doi.org/10.1016/j.foreco.2011.01.031
Elevitch CR (2006) Traditional trees of Pacific Islands: their culture, environment, and use. Permanent agriculture resources (PAR), Holualoa, United States of America. http://agroforestry.org/projects/traditional-tree-initiative. Accessed 24 June 2014
Feliciano D, Slee B, Hunter C, Smith P (2013) Estimating the contribution of rural land uses to greenhouse gas emissions: a case study of North East Scotland. Environ Sci Policy 25:36–49. https://doi.org/10.1016/j.envsci.2012.08.007
Feliciano D, Ledoa A, Hillier J, Nayaka DR (2018) Which agroforestry options give the greatest soil and above ground carbon benefits in different world regions? Agr Ecosyst Environ 254:117–129. https://doi.org/10.1016/j.agee.2017.11.032
Haile SG, Nair VD, Nair PKR (2010) Contribution of trees to carbon storage in soils of silvopastoral systems in Florida, USA. Glob Change Biol 16:427–438. https://doi.org/10.1111/j.1365-2486.2009.01981.x
Harvey CA, Chacon M, Donatti CI, Garen E, Hannah L, Andrade A, Bede L, Brown D, Calle A, Chara J, Clement C, Gray E, Hoang MH, Minang P, Rodriguez AM, Seeberg-Elverfeldt C, Semroc B, Shames S, Smukler S, Somarriba E, Torquebiau E, van Etten J, Wollenberg E (2014) Climate-smart landscapes: opportunities and challenges for integrating adaptation and mitigation in tropical agriculture. Conserv Lett 7:77–90. https://doi.org/10.1111/conl.12066
Hoosbeek MR, Remme RP, Rusch GM (2016) Trees enhance soil carbon sequestration and nutrient cycling in a silvopastoral system in south-western Nicaragua. Agroforest Syst. https://doi.org/10.1007/s10457-016-0049-2
Ibrahim M, Chacón M, Cuartas C, Naranjo J, Ponce G, Vega P, Casasola F, Rojas J (2007) Carbon storage in soil and tree biomass in different land use systems in cattle-dominated landscapes in Colombia, Costa Rica and Nicaragua. Agroforestería en las Américas 45:27–36. http://www.cipav.org.co/pdf/red%20de%20agroforesteria/Articulos/almacenamiento%20de%20carbono%20en%20el%20suelo%20y%20la%20biomasa%20arborea.pdf
INEGI (2005) Conjunto de datos vectoriales de la carta de uso del suelo y vegetación: escala 1:250,000. Serie III (continuo nacional). Instituto Nacional de Estadística, Geografía e Informática, Michoacán. http://www.inegi.org.mx/geo/contenidos/recnat/usosuelo/Default.aspx. Accessed 10 May 2014
IPCC (2007) Climate change 2007. Contribution of working group I to the fourth assessment report of the intergovernmental panel on climate change. IPCC WG1 AR4 Final Report. http://www.ipcc.ch/pdf/assessment-report/ar4/wg1/ar4_wg1_full_report.pdf. Accessed 4 Jan 2015
IPCC (2013) Climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the intergovernmental panel on climate change, Cambridge, United Kingdom and New York, NY, USA. https://doi.org/10.1017/cbo9781107415324. Accessed 19 Apr 2014
Jose S (2009) Agroforestry for ecosystem services and environmental benefits: an overview. Agrofor Syst 76:1–10. https://doi.org/10.1007/s10457-009-9229-7
Jose S (2012) Agroforestry for conserving and enhancing biodiversity. Agrofor Syst 85(1):1–8. https://doi.org/10.1007/s10457-012-9517-5
Leblanc HA, McGraw RL, Nygren P (2007) Dinitrogen-fixation by three neotropical agroforestry tree species under semi-controlled field conditions. Plant Soil 291(1–2):199–209. https://doi.org/10.1007/s11104-006-9186-0
Lorenz K, Lal R (2014) Soil organic carbon sequestration in agroforestry systems. A review. Agron Sustain Dev 34:443-454. https://doi.org/10.1007/s13593-014-0212-y
Luedeling E, Kindt R, Huth NI, Koenig K (2014) Agroforestry systems in a changing climate—challenges in projecting future performance. Curr Opin Environ Sustain 6:1–7. https://doi.org/10.1016/j.cosust.2013.07.013
Manning AD, Fischer J, Lindenmayer DB (2006) Scattered trees are keystone structures—implications for conservation. Biol Cons 132:311–321. https://doi.org/10.1016/j.biocon.2006.04.023
Matamala R, González-Meler MA, Jastrow JD, Norby RJ, Schlesinger WH (2003) Impacts of fine root turnover on forest NPP and soil C sequestration potential. Science 302:1385–1387. https://doi.org/10.1126/science.1089543
Matos ES, Freese D, Mendonca ES, Slazak A, Reinhard FH (2012) Carbon, nitrogen and organic C fractions in topsoil affected by conversion from silvopastoral to different land use systems. Agrofor Syst 81:203–211. https://doi.org/10.1007/s10457-010-9314-y
Mattsson E, Ostwald M, Nissanka SP, Pushpakumara DKNG (2015) Quantification of carbon stock and tree diversity of homegardens in a dry zone area of Moneragala district, Sri Lanka. Agrofor Syst 3:435–445. https://doi.org/10.1007/s10457-014-9780-8
Mbow C, Smith P, Skole D, Duguma L, Bustamante M (2014) Achieving mitigation and adaptation to climate change through sustainable agroforestry practices in Africa. Curr Opin Environ Sustain 6:8–14. https://doi.org/10.1016/j.cosust.2013.09.002
McGroddy ME, Lerner AM, Burbano DV, Schneider LC, Rudel TK (2015) Carbon stocks in silvopastoral systems: a study from four communities in southeastern Ecuador. Biotropica 47(4):407–415. https://doi.org/10.1111/btp.12225
Miranda T, Machado R, Machado H, Brunet J, Duquesne P (2008) Economic valuation of environmental goods and services in two ecosystems of cattle use. Zootec Trop 26(3):187–189. https://tspace.library.utoronto.ca/handle/1807/64190
Montagnini F, Nair PKR (2004) Carbon sequestration: an underexploited environmental benefit of agroforestry systems. Agrofor Syst 61–62(1–3):281–295. https://doi.org/10.1023/b:agfo.0000029005.92691.79
Mun HT, Whitford WG (1997) Changes in mass and chemistry of plant roots during long-term decomposition on a Chihuahuan Desert watershed. Biol Fertil Soils 26(1):16–22. https://doi.org/10.1007/s003740050336
Munroe JW, Isaac ME (2014) N2-fixing trees and the transfer of fixed-N for sustainable agroforestry: a review. Agron Sustain Dev 34(2):417–427. https://doi.org/10.1007/s13593-013-0190-5
Nair PKR (2012) Carbon sequestration studies in agroforestry systems: a realitycheck. Agrofor Syst 86:243–253. https://doi.org/10.1007/s10457-011-9434-z
Nakakaawa C, Aune J, Vedeld P (2009) Changes in carbon stocks and tree diversity in agro-ecosystems in south western Uganda: What role for carbon sequestration payments? New Forest 40(1):19–44. https://doi.org/10.1007/s11056-009-9180-5
Partey ST (2011) Effect of pruning frequency and pruning height on the biomass production of Tithonia diversifolia (Hemsl) A. Gray. Agrofor Syst 83(2):181–187. https://doi.org/10.1007/s10457-010-9367-y
Reyes G, Brown S, Chapman J, Lugo AE (1992) Wood densities of tropical tree species. United States Department of Agriculture, Forest Service Southern Forest Experimental Station, NewOrleans, Louisiana. General Technical Report SO-88. https://doi.org/10.2737/so-gtr-88
SAS Institute Inc (2004) SAS/STAT ® 9.1 User’s Guide. SAS Institute Inc., Cary, NC
Schmitt-Harsh M, Evans TP, Castellanos E, Randolph JC (2012) Carbon stocks in coffee agroforests and mixed dry tropical forests in the western highlands of Guatemala. Agrofor Syst 86(2):141–157. https://doi.org/10.1007/s10457-012-9549-x
Soto-Pinto L, Anzueto M, Mendoza J, Jimenez-Ferrer G, De Jong B (2010) Carbon sequestration through agroforestry in indigenous communities of Chiapas, Mexico. Agrofor Syst 78:39–51. https://doi.org/10.1007/s10457-009-9247-5
Tapia-Coral SC, Luiza FJ, Wandelli E, Fernandes ECM (2005) Carbon and nutrient stocks in the litter layer of agroforestry systems in central Amazonia, Brazil. Agrofor Syst 65:33–42. https://doi.org/10.1007/s10457-004-5152-0
Trumbmore SE, Davidson EA, Barbosa de Camargo P, Nepstad DD, Martinelli LA (1995) Belowground ciclin of carbon in forest and pastures of eastern Amazonia. Global Biogeochem Cycles 9:515–528. https://doi.org/10.1029/95gb02148
Villanueva-López G, Martínez-Zurimendi P, Ramírez-Avilés L, Casanova-Lugo F, Jarquín-Sánchez A (2014) Influence of livestock systems with live fences of Gliricidia sepium on several soil properties in Tabasco, Mexico. Cien Inv Agr 2:175–186. http://www.scielo.cl/pdf/ciagr/v41n2/art04.pdf
Villanueva-López G, Martínez-Zurimendi P, Casanova-Lugo F, Ramírez-Avilés L, Montañez-Escalante PI (2015) Carbon storage in livestock systems with and without live fences of Gliricidia sepium in the humid tropics of Mexico. Agrofor Syst 6:1083–1096. https://doi.org/10.1007/s10457-015-9836-4
Villanueva-López G, Casanova-Lugo F, Martínez-Zurimendi P, Parsons D, Aguilar-Solis L (2016) Effect of live fences of Gliricidia sepium on CO2 fluxes in tropical livestock systems. Soil Use Manag 32:553–564. https://doi.org/10.1111/sum.12311
Waldron A, Garrity D, Malhi Y, Girardin C, Miller DC, Seddon N (2017) Agroforestry can enhance food security while meeting other sustainable development goals. Trop Conserv Sci 10:1–6. https://doi.org/10.1177/1940082917720667
WRB (2007) The world reference base for soil resources. First update 2007. Reports on Global Soil Resources No. 103. FAO, Rome. http://www.fao.org/fileadmin/templates/nr/images/resources/pdf_documents/wrb2007_red.pdf. Accessed 27 Sept 2014
Xu X, Liu W, Zhang C, Kiely G (2011) Estimation of soil organic carbon stock and its spatial distribution in the Republic of Ireland. Soil Use Manag 27(2):156–162. https://doi.org/10.1111/j.1475-2743.2011.00342.x
Acknowledgements
We thank the Fundación Produce Michoacán A.C. for financially supporting this project, and we are also grateful to the National Council of Science and Technology for the support provided under infrastructure Project Number 270666. We thank Juan Carlos Gómez and Sergio Piñón, who contributed their experience and support, thereby facilitating the field work, and comments and suggestions from Fabien Charbonnier significantly improved the study. We acknowledge the comments from two anonymous reviewers on an earlier version of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
López-Santiago, J.G., Casanova-Lugo, F., Villanueva-López, G. et al. Carbon storage in a silvopastoral system compared to that in a deciduous dry forest in Michoacán, Mexico. Agroforest Syst 93, 199–211 (2019). https://doi.org/10.1007/s10457-018-0259-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-018-0259-x