
Abstract
In tropical areas of Mexico, Leucaena leucocephala is widely used in silvopastoral systems. However, little information exists on other native woody species of high forage potential, such as Guazuma ulmifolia. The aim of this study was to evaluate the components of biomass, forage yield and quality, and availability of N in fodder banks of L. leucocephala, G. ulmifolia, and a mixture of both species during dry and rainy seasons, under sub-humid tropical conditions. The experimental unit was a 5 × 10 m plot, containing three rows with 2 m between rows; each row had 20 plant positions with 0.50 m between plants. Within each plant position there was either a single plant, in the case of pure-crop, or two plants, in the case of mixed of both species. A complete randomized block design with three repetitions was used. In both seasons, there were a significantly greater proportion of leaves in the G. ulmifolia fodder banks (71 %) and in mixed fodder banks (69 %) than in L. leucocephala fodder banks (64 %). Consequently, these systems had leaf-to-stem ratios of 2.4, 2.2 and 1.9, respectively. The forage yield of fodder banks was not influenced by season. The mixed fodder bank had greater forage yield (5.1 t DM ha−1) than the L. leucocephala fodder bank (3.4 t DM ha−1) in each season. Additionally, the mixed fodder bank accumulated more forage yield during the experimental period (10.2 t DM ha−1 year−1) than G. ulmifolia (9.0 t DM ha−1 year−1) or L. leucocephala (6.9 t DM ha−1 year−1). The concentrations of CP, C and C:N were not influenced by season. Forage NDF and ADF concentrations were greater in the rainy season (476 g kg−1 DM) compared with the dry season (325 g kg−1 DM). Mixed fodder banks had the greatest N yield (185.9 kg ha−1) and consequently the greatest availability of N (371.8 kg N ha−1 year−1). We conclude that mixed fodder banks of L. leucocephala and G. ulmifolia are a better option for improving productivity and forage quality in comparison with pure fodder banks in Yucatan, Mexico.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In the humid and sub-humid tropics the increase of extensive farming has led to a rapid loss of natural resources. Some of the associated problems include deforestation, overgrazing of grasslands, un-controlled fires, productivity and biodiversity loss, water pollution, and vulnerability to climate change (Thornton et al. 2007; Murgueitio et al. 2011). Therefore, it is important to use strategies to solve these problems and mitigate natural resource degradation. Tree and shrub species not only contribute to reforestation and restoration of degraded areas (Verchot et al. 2007) but also provide an opportunity to overcome the problem of low availability and quality of forage during the dry season, and to improve animal performance (Olivares-Pérez et al. 2013). Often the nutritional value of leaves and fruits of these species is greater than from forage grasses (Piñeiro-Vázquez et al. 2013).
The Yucatan Peninsula has a great diversity of tree and shrub species which can be used for multiple purposes. For example, Guazuma ulmifolia Lam. (Sterculiaceae) and Leucaena leucocephala (Lam) de Wit. (Fabaceae) have been incorporated into agricultural systems as living fences, shade, firewood and forage (Casanova-Lugo et al. 2010; Murgueitio et al. 2011; Ortega-Vargas et al. 2013). Studies have described the incorporation of L. leucocephala in silvopastoral systems (Kiwia et al. 2009; Murgueitio et al. 2011), but little information exists on the use of other potentially useful woody species in the tropics of southern Mexico. Species grown together in mixed stands can promote benefits such as regulation of damaging organisms, improvement of nutrient recycling, increase of biomass production, improvement of organic matter supply, soil erosion protection, and increased product quality such as improved forage production and quality (Altieri and Nicholls 2003).
Therefore, the objective of this study was to evaluate the biomass conformation and forage yield and quality of L. leucocephala and G. ulmifolia, in pure and mixed fodder banks, during the dry and rainy season in a sub-humid tropical zone.
Materials and methods
Site characteristics
The experiment was carried out at the Campus of Biological and Agricultural Sciences of the University of Yucatan (UADY) in Yucatan state, Mexico from January to December 2009 (21°51′ N and 89°41′ W; Fig. 1).

Study site located in Xmatkuil, Yucatan, Mexico (http://www.seduma.yucatan.gob.mx/ordenamiento-ecologico/index.php)
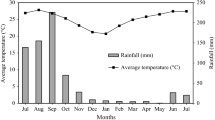
The area where the experiment was located is climatically classified as Aw0 (i.e. warm and sub-humid; according to Garcia 1988), with an average annual rainfall of 953 mm. The annual mean temperature is 26.5 °C, with annual maximum and minimum ranges from 36 to 40 and from 14 to 16 °C, respectively (Garcia 1988). Climatic data during the experimental period (Fig. 2) was recorded daily with a Hobo® data logger. Mean, maximum, and minimum annual temperatures and total rainfall during this period were 24.2, 37.2, 17.0 °C and 721 mm, respectively. The area was located within a karst plateau, which is characterized by a flat or gently rolling relief. Soils are shallow, heterogeneous, rocky (limestone) and clay-loam, with a pH of 7.5–7.8 (Bautista et al. 2011).

Minimum and maximum air temperatures, and rainfall at the study site. Vertical arrows (↓) indicate time of biomass harvesting (Data taken from the microclimatic station at the Campus of Biological and Agricultural Sciences. University of Yucatan, Mexico in 2009)
Management of experimental plots
The fodder banks systems formed by woody species native from Yucatan were established during the rainy season of 2004. Plots of pure L. leucocephala or G. ulmifolia, and plots of a mixture of both species (L. leucocephala and G. ulmifolia) were arranged in a randomized complete block design with three replicates; each experimental unit was 5 × 10 m (Fig. 3). Plots were established using seedlings of 30-cm height, with 2.0 m between rows and 0.5 m between plants. Each planting position had either one seedling, in the pure fodder bank treatments, or two seedlings, in the mixed fodder bank treatments (one of each tree species) (Fig. 3).

Diagram of the experimental plots
In October 2008, 3 months before data collection, a cleaning cut was undertaken in all the plots, at 1-m height, to avoid residual effects of previous management and to reduce the heterogeneity between trees. Three months later, five plants of each species were randomly selected, from the central rows of each plot in the three replicates, to assess forage yield and quality. This included fifteen plants from each pure fodder bank (monoculture) and 30 plants (15 of each species) from the mixed forage bank.
Above ground biomass was harvested every 3 months; two times in the dry season (January and April 2009) and two times in the rainy season (July and October 2009), at a cutting height of 1.0 m, following the methodology described by Pezo and Ibrahim (1998). During the dry season, drip irrigation was applied in the morning for 3 h (approximately 1 l h−1 plant−1), two times per week.
Biomass components and forage yield
After harvesting, the biomass from each fodder bank was weighed fresh. Harvested material from each bank was pooled and three sub-samples (of approximately 500 g each) were randomly taken. These sub-samples were divided into leaves, edible (<0.5 cm diameter) and woody stems (>0.5 cm diameter), which were dried at 60 °C in an fan-forced oven to constant weight. Only leaves and edible stems were considered for calculating forage yield.
Seasonal forage yield was obtained by adding the yield from both harvests. Leaf and stem proportions per season were estimated from an average of the two harvests within a season.
Chemical analysis
Forage sub-samples (leaves and edible stems) were ground using an electric Thomas-Wiley mill to a particle size of 1 mm and analyzed for neutral detergent fiber (NDF) and acid detergent fiber (ADF) using an ANKOM (Macedon, NY) A200 fiber analyzer. Fractions of carbon (C) and nitrogen (N) were estimated using a Leco CN 2000 elemental analyzer (Leco Instruments, Inc., St. Joseph, MI) and C-to-N ratio was calculated. N yield (t N ha−1) was calculated for each harvest period by multiplying the N fraction by the forage yield.
Statistical analysis
Forage and N yield data were analyzed with a repeated measures ANOVA model using PROC MIXED (SAS 2004) to examine the effect of the season, the fodder bank type, and their interactions. For biomass components (data were transformed to square root) and chemical composition, a multivariate analysis of variance (MANOVA) was applied, using PROC GLM (SAS 2004). Where significant differences were found, means were compared using Tukey’s statistic (P ≤ 0.05).
Results
Biomass composition
Leaf proportion was greater in the G. ulmifolia (71 %) and mixed fodder banks (69 %) than L. leucocephala fodder banks (66 %, P < 0.001) (Table 1). The proportions of edible stems (12–14 %) and woody stems (17–20 %) were similar (P > 0.05) among fodder banks. G. ulmifolia and mixed fodder banks had a greater (P < 0.001) leaf-to-stem ratio (2.4 and 2.2, respectively) than L. leucocephala fodder bank (1.9). Biomass components also were influenced by season. Leaf proportion and leaf-stem ratio were greater (P < 0.01) in the rainy season (70 % and 2.3, respectively) than in the dry season (67 % and 2.0, respectively). In contrast, the proportion of edible stems was greater (P < 0.001) in the dry season (16 %) than the rainy season (11 %). The proportion of woody stems (18 %) was similar among seasons. Overall, fodder banks had an average edible fraction (leaves and edible stems) of 82 %.
Forage yield
Mixed fodder banks had greater (P < 0.05) forage yield (5.1 t DM ha−1) than L. leucocephala (3.4 t DM ha−1). G. ulmifolia fodder banks had a similar forage yield (4.5 t DM ha−1) to the other fodder banks (Table 2). There was no effect of season on forage yield (Table 2, P > 0.05). Average forage yield per season was 4.3 t DM ha−1. Throughout the year, mixed fodder banks system accumulated more forage (10.2 t DM ha−1 year−1) than G. ulmifolia (9.0 t DM ha−1 year−1) or L. leucocephala (6.9 t DM ha−1 year−1) in monocultures (Fig. 4).

Annual forage yield of L. leucocephala and G. ulmifolia in pure and mixed fodder banks under tropical conditions of Yucatan. Error bars represent the standard error of the mean. Means labelled by different letters are significantly different (Tukey’s statistic, P ≤ 0.05)
Forage quality
Leucaena fodder banks had greater (P < 0.01) concentrations of CP (228 g kg−1 DM) and C (445 g kg−1 DM) than mixed (179.5, 432.4 g kg−1 DM) and G. ulmifolia fodder banks (145, 425.3 g kg−1 DM). On the other hand, G. ulmifolia fodder banks had a greater C:N ratio (18.4) than mixed (15.1) and L. leucocephala (12.2) fodder banks (P < 0.001). The concentrations of NDF and ADF in the forage were similar in all fodder banks (P > 0.05, Table 3).
The mean concentrations of CP, C, and C:N ratio were 184, 434 g kg−1 DM, and 15.2, respectively, and were not influenced by season (P > 0.05). However, forage NDF and ADF concentrations were greater (P < 0.01) in the rainy season, 476 and 325 g kg−1 DM, respectively) than in the dry season, 427 and 244 g kg−1 DM, respectively. There was no interaction between fodder bank types and season (P > 0.05, Table 3).
Nitrogen yield
There was no significant (P > 0.05) effect of season or the interaction between fodder banks and season on N yield (Table 4). N yield averaged across seasons was 129.8 kg N ha−1 season−1. However, there was a significant (P < 0.001) effect of type of fodder bank on N yield. Mixed fodder banks had greater N yield (185.9 kg N ha−1 season−1) than L. leucocephala (99.2 kg N ha−1 season−1) or G. ulmifolia (104.3 kg N ha−1 season−1) fodder banks.
In addition, mixed fodder banks accumulated the greatest annual N yield (371.8 kg N ha−1 year−1), followed by L. leucocephala (250.3 kg N ha−1 year−1) and G. ulmifolia (208.5 kg N ha−1 year−1) fodder banks (P < 0.05; Fig. 5).

N yield from L. leucocephala and G. ulmifolia pure and mixed fodder banks under tropical conditions of Yucatán. Error bars represent the standard error of the mean. Means labelled by different letters are significantly different (Tukey’s statistic, P ≤ 0.05)
Discussion
Biomass composition
The lower proportion of leaves and leaf to stem ratio in the dry season, compared to the rainy season, could be associated with the relationship between plant growth and climatic conditions. In particular, temperature and photoperiod likely played an important role in plant growth and development. Temperature recorded early in the dry season (February to March) was lower than during the raining season (Fig. 2). Also, during this period the photoperiod is shorter than in the rainy season (the longest night occurs on the 21st of December, while the longest day is on the 21st of June). These two factors (low temperature and short photoperiod) could have promoted flowering. Consequently, as a result of this stimulus, transportation of carbohydrates favored stem elongation and flower formation at the expense of leaf formation. It is known that during the dry season, both of the studied trees species flower and produce seed in the region (Latt et al. 2000; Ortega-Vargas et al. 2013).
Among all fodder banks, the biomass consisted of 68 % leaf and a smaller proportion of edible (13 %) and woody stems (18 %), and the mean leaf-to-stem ratio was 2.2. Consequently, the edible fraction was approximately 80 % of the harvested biomass (Table 1). These results are similar to those reported by Petit-Aldana (2011) who found that when cut at 3-month intervals, L. leucocepahala, G. ulmifolia, and a mixture of both species had an average of 61 % leaves, 19 % edible stems, 20 % woody stems, and leaf-to-stem ratio of 1.7. Additionally, they found that the edible fraction varied from 74 to 88 % depending on the season. The results are also consistent with those reported by Casanova-Lugo et al. (2010), who reported that 6 year-old L. leucocephala and G. ulmifolia fodder banks, pruned at three-month intervals, produced similar leaf proportions to the current study.
G. ulmifolia and mixed fodder banks had the greatest proportion of leaves and, consequently, the greatest leaf-to-stems ratio (Table 1). This is likely related to intrinsic characteristics of these species and their resilience to severe pruning.
Forage yield
Mixtures of forage tree species can provide important benefits to the productivity of the system. Mixed tree crops may have advantages compared with monoculture systems, mainly in terms of increased biomass production per unit area (Rosales-Méndez and Gill 1997). Additionally, they are fast-growing and better able to tolerate long dry periods than grasses. Therefore, the inclusion of trees should extend forage production further into the dry season. In a study by Reyes-Garcia (2009) in pure and mixed fodder banks, at densities of 10,000 and 20,000 plants ha−1, mixed banks had a forage yield of 5.8 t DM ha−1 yr−1, harvested at four-month intervals, whereas pure forage crops of G. ulmifolia and L. leucocephala yielded almost 40 % less (3.8 and 3.5 t ha−1 yr−1, respectively). Further studies, on similar soil conditions and cutting at three-month intervals, showed that the mixed fodder bank produced the greatest amount of forage (7.0 t ha−1 yr−1) (Petit-Aldana 2011).
In the present study, greater forage yield was obtained from mixed fodder banks than from pure crops, especially L. leucocephala fodder bank, which yielded almost 50 % lower than the mixed fodder bank. However, mixed fodder bank yield was only 13 % higher than the G. ulmifolia fodder bank (Fig. 4). This result is consistent with the report of Petit-Aldana (2011), who found that the edible biomass of mixed fodder banks increased by 50 % compared with pure plots of L. leucocephala, and 30 % compared with pure crop of G. ulmifolia. These yields increments could be partly due to plant density, since there were twice as many plants in the mixed fodder bank than in the pure crop banks. Increasing plant density results in more yield up to a point, after which yield could decline due to plant competition for growing resources (e.g. water, nutrients). Another factor likely to be involved in the higher yield in the mixed fodder bank compared to pure crop banks is a beneficial relationship between L. leucaena and G. ulmifolia, as suggested by Pretzsch and Schütze (2009) for other tree species. This advantage of the crop mixture over the pure crops could be a consequence of different root structures, especially depth of rooting. Also, an important factor for the co-existence of two species in the same niche is that the species must use resources differently, which suggests that mixture fodder banks may utilize resources more completely than the single species, leading to a greater overall productivity. Nevertheless, this account only for part of the results, since the soils of the area have limited resources, and some level of competition in the mixed crops could be expected, inducing a yield reduction of each associated individual plant, despite the a superior total yield. Another mechanism which could explain this advantage of the mixture crop is “facilitation”, which involves the improvement of N availability, either by litter decomposition and/or N biological fixation (Freitas et al. 2010). Although in the current work N-fixation was not measured, L. leucocephala is known for its ability to fix large amounts on N (Danso et al. 1992), especially under low fertile and calcareous soils such as those found in the experimental site. Also, L. leucocephala foliage may have recycled N to the system since it is rapidly degraded in the soil. The results shows that L. leucocephala and G. ulmifolia are compatible species, whose association increases overall biomass yield. We hypothesize that this advantage was due to the different rooting pattern and depth (as has been shown by Tamayo-Chim et al. 2012), and the N-fixing ability of the legume.
There was no influence of season on forage yield of fodder bank type (pure and mixed), suggesting that these species have some tolerance to periods of drought and periodic pruning, as indicated by Tamayo-Chim et al. (2012) and Casanova-Lugo et al. (2010), and that the supplementary irrigation was enough to encourage plant growth. However, Petit-Aldana (2011) found that in the dry season fodder tree species subject to periodic pruning reduce their production by 6 to 30 %, compared with the rainy season, due to depletion of carbohydrate reserves and thus delayed recovery.
Forage quality
The different tree fodder banks showed differences in dry matter and CP concentrations, which could be caused by genotype, environmental factors, and management, such as pruning (Casanova-Lugo et al. 2010; Petit-Aldana 2011; Ortega-Vargas et al. 2013).
In Matanzas, Cuba, González-García et al. (2009) found that the CP concentration of the edible fraction of L. leucocephala ranged from 190 to 234 g kg−1 DM. For G. ulmifolia, in Chiapas, Mexico, Jiménez-Ferrer et al. (2008) reported CP values ranging from 136 to 149 g kg−1 DM. Ayala-Burgos et al. (2006) found that the CP concentrations ranged from 63 to 310 g kg−1 DM for L. leucocephala and 26 to 198 g kg−1 DM for G. ulmifolia. In Yucatan, Mexico, Reyes-García (2009) reported that the CP concentrations of L. leucocephala fodder banks were 183 and 188 g kg−1 DM for cutting intervals of 4 and 6 months, respectively. In the same study, CP concentrations were 111 and 106 g kg−1 DM for G. ulmifolia fodder banks, and 155 and 149 g kg−1 DM for mixed fodder banks for cutting intervals of 4 and 6 months, respectively. Under similar soil and climatic conditions, Petit-Aldana (2011) found CP concentrations of 195 g kg−1 DM for L. Leucocephala, 124 g kg−1 DM for G. ulmifolia, and 163 g kg−1 DM for the mixture of both species, with a harvest interval of 3 months. These results demonstrate the variability in CP concentration, but also the greater CP concentration in L. leucocephala than G. ulmifolia.
In this study (Table 3), pure fodder banks of L. leucocephala had a 57 % higher CP concentration than the pure fodder bank of G. ulmifolia and a 27 % higher CP concentration than the mixed system. L. leucocephala had the greatest concentration of CP due its ability to fix atmospheric N, which is a great advantage for forage production in the tropics (Danso et al. 1992). However, both forage tree species (either in pure or mixed crops) provide greater protein concentrations than typical for tropical pastures (e.g. 7 % CP, Jackson and Ash 1998). In addition, these tree species maintain CP concentrations content above 12 % even in the dry season, a level recommended to select good quality forage (Olivares-Pérez et al. 2013; Ortega-Vargas et al. 2013).
NDF is a measure of fiber concentration in forage, and includes cellulose, hemicellulose and lignin, and is negatively correlated with dry matter intake (Piñeiro-Vázquez et al. 2013). ADF is a similar measure, but does not include hemicellulose and it is empirically correlated with digestibility. NDF and ADF can vary with species, age of tissue, management, and climatic variations (Grewal and Abrol 1986; Larbi et al. 2005; Ortega-Vargas et al. 2013).
Petit-Aldana (2011) reported that average NDF and ADF concentrations in L. leucocephala, and G. ulmifolia and the mixture of both species were similar (470 and 330 g kg−1 DM). However, in the rainy season they reported greater values than in the dry season for NDF (464 and 417 g kg−1 DM), and for ADF (360 and 290 g kg−1 DM). In agreement with these authors, in the current study, NDF and ADF concentrations were similar among the fodder banks (average 450 and 285 g kg−1 DM), and there were significant differences between seasons (Table 3).
NDF and ADF values in this study (Table 3) are lower than those of typical tropical grasses (Rosales-Méndez and Gil 1997), highlighting their superior quality for feeding. Grasses have a greater concentration of structural material and a lower N concentration in comparison to the foliage of trees, which gives trees an advantage in terms of digestibility, forage intake, and the effect on animal production (Jackson and Ash 1998; Olivares-Pérez et al. 2013; Ortega-Vargas et al. 2013).
C concentration is an indicator of photosynthetic production as well as the ability of plants to store it in their tissues (Hamburg 2000). Brown et al. (2007) stated that approximately 50 % of the dry weight of any plant species consists of C. However, Kirby and Potvin (2007) argued that this content varies between species and between different plant parts and it is generally greater in woody tissues, mainly due to the greater lignin concentration (Larbi et al. 2005). In the current study, the concentration of C in the edible biomass of L. leucocephala pure fodder bank system was greater (445 g kg−1 DM) than G. ulmifolia pure fodder bank (425 g kg−1 DM) and the mixture fodder banks (432 g kg−1 DM) (Table 3). These values are within the range of other studies; for example, Petit-Aldana (2011) reported that C concentration in the edible fraction of L. leucocephala, G. ulmifolia and M. oleifera fodder banks was on average 427 g kg−1 DM, and that L. leucocephala had the highest concentration of C among these species (438 g kg−1 DM). Similarly, Alvarado et al. (2007) reported that C concentrations of Gliricidia sepium and Erythrina sp. foliage were 440 and 433 g kg−1 DM, respectively, when they were pruned to 80-cm height.
Numerous studies report that the C: N ratio and concentrations of lignin and polyphenols in the plant are factors in the decomposition process (Mafongoya et al. 1998; Valenzuela-Solano and Crohn 2006). Plant material with low lignin and high N content generally decays faster than material with more lignin and less N. Species with high C:N (>25) will require more time for degradation. In contrast, species with low C:N will require less time to be degraded. For example, L. leucocephala has a low C:N ratio due to its ability to fix N from the atmosphere (Petit-Aldana 2011). In addition, C:N ratio influences rumen digestibility, and therefore, the level of methane production (Hindrichsen et al. 2005).
In general, the C:N values reported in this study are adequate, which confirms their potential use in animal nutrition. The tree species studied have high nutritional properties that can improve animal diets. They can also be used to achieve ecological and economic targets, by reducing external inputs and improving nutrient cycling in the animal production system (Petit-Aldana 2011). However, this depends not only on the characteristics of the species, but also on site resource availability, such as light, water, soil nutrients, and management of the agro-ecosystem (Casanova-Lugo et al. 2010).
Nitrogen yield
Nitrogen is often the most limiting element in tropical animal production systems (Graham and Vance 2003). The use of tropical trees and shrubs could contribute significantly to the sustainability of these systems by making them less dependent on external inputs (Verchot et al. 2007; Olivares-Pérez et al. 2013). This study found that although L. leucocephala had greater total N yield than G. ulmifolia, the mixed forage banks had the greatest total N yield (371 kg ha−1) (Fig. 5). The yield of N depends mainly on the selected species and management practices (i.e. irrigation, fertilization, pruning, etc.) that have a role in the production and quality of forage (Grewal and Abrol 1986; Larbi et al. 2005). Leguminous trees that fix N have an advantage over other species in accumulating N, and subsequently also in biomass production (Danso et al. 1992; Freitas et al. 2010).
Conclusions
Mixed fodder banks have the potential to be developed in the growing conditions of Yucatan, and likely in other tropical regions, mainly due to their high forage yields and ability to provide high N yield to the whole system. Overall, leaf proportion and CP, important attributes that determine forage quality, were better in the mixture bank than in pure fodder banks. Further studies are needed to assess the economics of establishment because the greater density of plants in the mixed fodder banks implies increased cost.
References
Altieri MA, Nicholls CI (2003) Soil fertility management and insect pests: harmonizing soil and plant health in agroecosystems. Soil Till Res 72:203–211
Alvarado MF, Rodríguez JC, Cerrato M (2007) Concentración de carbono y nitrógeno a seis frecuencias de poda en Gliricidia sepium y Erythrina sp. Tierra Trop 3(2):211–220
Bautista F, Palacio-Aponte G, Quintana P, Zinck JA (2011) Spatial distribution and development of soils in tropical karst areas from the Peninsula of Yucatan, Mexico. Geomorphology 135:308–321
Brown S, Hall M, Andrasko K, Ruiz F, Marzoli W, Guerrero G, Masera O, Dushku A, DeJong B, Cornell J (2007) Baselines for land-use change in the tropics: application to avoided deforestation projects. Mitig Adapt Strat Glob Change 12:1001–1026
Casanova-Lugo F, Ramírez-Avilés L, Solorio-Sánchez FJ (2010) Effect of pruning interval on foliage and root biomass in forage tree species in monoculture and in association. Trop Subtrop Agroecosyst 12:33–41
Danso SKA, Bowen GD, Sanginga N (1992) Biological nitrogen fixation in trees in agro-ecosystems. Plant Soil 141(1–2):177–196
Freitas AD, Sampaio EV, Santos CE, Fernandes AR (2010) Biological nitrogen fixation in tree legumes of the Brazilian semi-arid caatinga. J Arid Environ 74(3):344–349
García E (1988) Modificaciones al sistema de clasificación climática de Köppen. Universidad Nacional Autónoma de México (UNAM), Mexico
González-García E, Cáceres O, Archimède H, Santana H (2009) Nutritive value of edible forage from two Leucaena leucocephala cultivars with different growth habit and morphology. Agroforest Syst 77:131–141
Graham PH, Vance CP (2003) Legumes: importance and constraints to greater utilization. Plant Phys 131:872–877
Grewal SS, Abrol IP (1986) Agroforestry on alkali soils: effect on some management practices on initial growth, biomass accumulation and chemical composition of selected tree species. Agrofor Syst 4(3):221–232
Hamburg S (2000) Simple rules for measuring changes in ecosystem carbon in forestry-offset projects. Mit Adapt Strat Glob Chan 5(1):25–37
Hindrichsen IK, Wettstein HR, Machmüller A, Jörg B, Kreuzer M (2005) Effect of the carbohydrate composition of feed concentratates on methane emission from dairy cows and their slurry. Env Mon Assess 107(1–3):329–350
Jackson J, Ash AJ (1998) Tree-grass relationships in open eucalypt woodlands of northeastern Australia: influence of trees on pasture productivity, forage quality and species distribution. Agrofor Syst 40(2):159–176
Jiménez-Ferrer G, López-Carmona M, Nahed-Toral J, Ochoa-Gaona S, de Jong B (2008) Fodder trees and shrubs of the north–tzotzil region of Chiapas. Mexico Vet Méx 39(2):199–213
Kirby KR, Potvin C (2007) Variation in carbon storage among tree species: implications for the management of a small-scale carbon sink project. For Ecol Manage 246:208–221
Kiwia A, Imo M, Jama B, Okalebo JR (2009) Coppicing improved fallows are profitable for maize production in striga infested soils of western Kenya. Agrofor Syst 76:455–465
Larbi A, Anyanwu NJ, Oji UI, Etela I, Gbaraneh LD, Ladipo DO (2005) Fodder yield and nutritive value of browse species in West African humid tropics: response to age of coppice regrowth. Agrofor Syst 65(3):197–205
Latt CR, Nair PKR, Kang BT (2000) Interactions among cutting frequency, reserve carbohydrates, and post-cutting biomass production in Gliricidia sepium and Leucaena leucocephala. Agrofor Syst 50(1):27–46
Mafongoya P, Giller K, Palm C (1998) Decomposition and nutrient release patterns of prunings and litter of agroforestry trees. Agrofor Syst 38:77–97
Murgueitio E, Calle Z, Uribe F, Calle A, Solorio B (2011) Native trees and shrubs for the productive rehabilitation of tropical cattle ranching lands. For Ecol Manage 261:1654–1663
Olivares-Pérez J, Avilés-Nova F, Albarran-Portillo B, Castelán-Ortega OA, Rojas-Hernández S (2013) Use of three fodder trees in the feeding of goats in the subhumid tropics in Mexico. Trop Anim Health Prod 45:821–828
Ortega-Vargas E, López-Ortiz S, Burgueño-Ferreira JA, Campbell WB, Jarillo-Rodríguez J (2013) Date of pruning of Guazuma ulmifolia during the rainy season affects the availability, productivity and nutritional quality of forage during the dry season. Agrofor Syst. doi:10.1007/s10457-013-9608-y
Petit-Aldana JC (2011) Fodder tree species in association to improve productivity and nutrients cycling. Campus of biological and agricultural sciences. University of Yucatan, Merida, p 137
Pezo D, Ibrahim M (1998) Sistemas Silvopastoriles. Módulo de Enseñanza Agroforestal No. 2. Proyecto Agroforestal CATIE/GTZ, Turrialba, p 258
Piñeiro-Vázquez AT, Ayala-Burgos AJ, Chay-Canul AJ, Ku-Vera JC (2013) Dry matter intake and digestibility of rations replacing concentrates with graded levels of Enterolobium cyclocarpum in pelibuey lambs. Trop Anim Health Prod 45:577–583
Pretzsch H, Schütze G (2009) Transgressive over yielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: evidence on stand level and explanation on individual tree level. Eur J Forest Res 128(2):183–204
Reyes-García J (2009) Efecto de la edad de rebrote sobre el rendimiento, composición química y digestibilidad del forraje de árboles (monocultivos y asociados). Equiparabilidad de las técnicas In vitro e In situ para estimar la digestibilidad de la MS. Tesis de Maestría en Ciencias. Universidad Autónoma de Yucatán, Mexico, p 57
Rosales-Méndez M, Gill M (1997) Tree mixtures within integrated farming systems. Livest Res Rural Dev 9(4):56–66
SAS (2004) Statiscal Analysis System, Users. SAS Institute, Cary
Tamayo-Chim M, Reyes-García C, Orellana R (2012) A combination of forage species with different responses to drought can increase year-round productivity in seasonally dry silvopastoral systems. Agrofor Syst 84(2):287–297
Thornton P, Herrero M, Freeman A, Mwai O, Rege E, Jones P, McDermott J (2007) Vulnerability, climate change and livestock—research opportunities and challenges for poverty alleviation. J SAT Agri Res 4(1):23
Valenzuela-Solano C, Crohn D (2006) Are decomposition and N release from organic mulches determined mainly by their chemical composition? Soil Biol Biochem 38:377–384
Verchot LV, Van Noordwijk M, Kandji S, Tomich T, Kandji M, Ong C, Albrecht A, Mackensen J, Bantilan C, Anupama KV, Palm C (2007) Palm C (2007) climate change: linking adaptation and mitigation through agroforestry. Mitig Adapt Strat Glob Change 12:901–918
Acknowledgments
The authors are grateful to Consejo Nacional de Ciencia y Tecnología (CONACyT) for the project FORDECyT, key 117072. We would like to thank the CONACyT for the support granted to M.Sc. Fernando Casanova Lugo to perform his Doctoral studies at Autonomous University of Yucatan, Mexico.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Casanova-Lugo, F., Petit-Aldana, J., Solorio-Sánchez, F.J. et al. Forage yield and quality of Leucaena leucocephala and Guazuma ulmifolia in mixed and pure fodder banks systems in Yucatan, Mexico. Agroforest Syst 88, 29–39 (2014). https://doi.org/10.1007/s10457-013-9652-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-013-9652-7