
Abstract
Hyphaene thebaica Mart. (doum palm) is an agroforestry tree with high ecological and economic value, but currently its populations are harvested excessively, which is likely to increase in the future. This study assessed the current status of this species with regard to increasing landscape modification and human pressure in Benin. We compared the structure of adult palms in farmlands to those within the Biosphere Reserve of Pendjari (BRP). In addition, spatial patterns and sex ratio of the species were compared between both land use types. Results showed that mean diameter (adult palms) and density (adult palms and seedlings) were significantly higher (P < 0.001) in BRP than in farmlands. However, no significant differences were noticed for doum palm height and density of juveniles (P > 0.05). The pair correlation function showed globally a random pattern for all palm life stages, albeit with a weak aggregation in farmlands. In the BRP, a strong aggregated pattern is observed for seedlings, whereas all other palm life stages showed globally a random pattern. Moreover, no spatial association was observed within palm life stages and between palm life stages and other tree species, but did exist between females and seedlings in the BRP. The sex ratio did not depart from 0.5 in both land use types. We conclude that in spite of the land use difference, the doum palm species is still well preserved. However, rapid land-use intensifications may lead to increasing pressure on the species populations in the future.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Agroforestry systems have received much attention in conservation research because they help to reduce the rates of conversion of natural habitat by providing a more productive, sustainable alternative to traditional agricultural systems that may involve clearing natural habitats (Bhagwat et al. 2008; Teklehaimanot 2003). These systems provide ideal study areas for investigating spatial patterning of tree populations, as they contain relatively few tree species and have simple dynamics, reducing their complexity. Agroforestry is currently among the most widespread land use management systems in sub-Saharan Africa (Teklehaimanot 2003) and holds several tree species, including Hyphaene thebaica Mart.
Hyphaene thebaica (commonly called doum palm), native to the semi-arid Sahelian zone of West Africa is an important tree species growing wild throughout the dry regions of tropical Africa, the Middle East and Western India (Fanshawe 1966). The species is a valuable source of a variety of products for many people, especially those living in its vicinity (Hsu et al. 2006). Local farmers harvest the fruit for food, the foliage for local crafts, the stem for construction material, and the roots for medicinal use (Moussa et al. 1998). All these uses involve overexploitation of vital organs of individuals of the species resulting in a significant reduction in the doum palm populations present in the region (Moussa et al. 1998). Doum palm is also considered as an indicator of fertile soils (Giffard 1966; Jahiel 1993), often resulting in the clearance of the species stands for agricultural needs.
Many ecological methods have been developed and used recently for assessing the ecological status of agroforestry trees (Dawson et al. 2009; Isaac et al. 2009). These approaches include quantification of the population structure and determination of the spatial patterns of the species. Indeed, several ecological investigations focused on the population structure and dynamics of some useful tree species in Sub-Saharan Africa (Gaoue and Ticktin 2008; Glèlè Kakaï et al. 2011; Ouédraogo et al. 2013). However, few of those ecological studies have specifically addressed the need for a detailed understanding of savanna palm, such as doum palm stand structure.
Spatial pattern is a crucial aspect of natural vegetation because it affects the dynamics of the plants themselves and of a range of other organisms with which they interact (Gelfand et al. 2010). The spatial scale at which pattern is known to affect processes range from the neighborhood of an individual plant, to as broad as landscapes, where it may affect biodiversity and ecosystem functions (Turner 1989).
The type of reproduction and mating system constitute important factors shaping the dynamics of a given plant species at individual or population levels (Pannell 2002). Sex ratio is known to affect both the growth rates and the evolutionary trajectories of wild populations (Sapir et al. 2008) because it affects the successful mating probability of a female. Although genetic and environmental factors are recognized as determinant drivers of individual sex and population-level sex ratios, anthropogenic impacts are also likely to shape the population sex ratio in a given area.
Human disturbances are among environmental factors defining spatial patterns of species (Lykke 1998). The significant effect of human disturbance, such as logging, land clearing, and/or fire on population structure and spatial patterns of tropical species has been highlighted extensively (Djossa et al. 2008; Haarmeyer et al. 2013; Haugo et al. 2010). At the landscape level, potentially widespread and increased disturbances, such as plant cutting, can have an obvious effect on stand structure and spatial organization, especially for trees (Haugo et al. 2010). As the abundance of an organism is dependent on the number of individuals that survive and reproduce, any factors that affect survival or reproduction will affect abundance and spatial arrangement (Jorgensen and Fath 2008). Consequently, anthropogenic disturbances could have a serious impact on the spatial pattern of doum palm. Understanding the population patterns and underlying intrinsic factors is critical for the conservation and sustainable management of this agroforestry species.
The objective of this study was to assess the effects of human disturbance on the structural characteristics of doum palm. More specifically, we tested the following five hypotheses with regard to doum palm populations within farmland habitats and the BRP:
-
1.
practices in farmlands negatively affect the structural parameters and population structure of doum palm;
-
2.
doum palm life stages show clumped patterns in BRP, but random pattern in farmlands;
-
3.
adult palms and seedlings show a positive spatial association in both land use types;
-
4.
doum palm life stages and other trees species show no spatial association in both land use types;
-
5.
the sex ratio of doum palm is female-biased.
Materials and methods
Study area
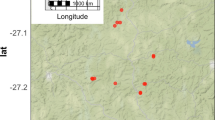
The study was undertaken in the vicinity and within the BRP, with a particular focus on the hunting zone where doum palm exhibits higher occurrence (Fig. 1). The region is located in the extreme northwest of Benin in the district of Atacora (10°40′–11°28°N and 0o°7′-2°10′E), covering an area of 4661.4 km2 and includes the National Park of Pendjari (2660.4 km2), the hunting zone of Pendjari (1750 km2) and the hunting zone of Konkombri (251 km2). The Pendjari is the primary river carrying water throughout the year, and runs through the National Park of Pendjari and the hunting zone of Pendjari (Djossa et al. 2008). The park is located in the Sudanian zone with one rainy season (April–May to October). The total rainfall averages 1000 mm with 60 % falling between July and September (Sinsin et al. 2004), and one dry season (November to March–April). The BRP is bordered by many small villages. Population density in this area is low compared to the majority of the country, and the main activities of the local population are agriculture and livestock rearing (INSAE 2015).

Map of study area: a Benin in Africa, b the Biosphere reserve of Pendjari in Benin; and c the position of samples plots in the hunting zone
Sampling and data collection
Structural parameters and stand structure of doum palm
Previous fieldwork helped to distinguish three successive life stages: seedlings, juveniles (which bear mature leaves but are not able to reproduce), and adults (females and males). Those life stages have been considered in the study.
For assessing the impact of land use types structural parameters and population structure of doum palm (hypothesis 1), thirty 1-ha square plots (100 m × 100 m) were installed in different habitats that were assigned to two different land-use types: farmlands and BRP. These land-use types differed according to the level of human pressure. Human disturbance was lowest in the BRP and was limited to Non Timber Forest Products (NTFP) harvesting and management fire, whereas it was highest in farmlands and was characterized by NTFP-harvesting, fire for agricultural land clearing, and livestock grazing. Within each ha-plot, sex and diameter (for adult palms), and total height were collected.
Spatial patterns and spatial association among doum palm stages and other tree species
For assessing the spatial patterns of doum palm (hypotheses 2 and 3), three different stands size were chosen both in farmlands and in the BRP: 150 × 150 m (stand 1), 200 × 200 m (stand 2) and 400 m x 400 m (stand 3) (Figs. 2 and 3). Each plot was subdivided into quadrats of 50 × 50 m (Azihou 2013), delimited by poles that created a grid system used for mapping. All palms (seedlings, juveniles and adults) and all other tree species exceeding 10 cm in diameter at breast height were mapped. Data were collected on the distance of each adult palm to its neighbours (or other tree species), the azimuth of each tree to the nearest neighbour, and its slope using respectively a rangefinder, a compass and a clinometer. The following characteristics were recorded for each stand: soil texture, human disturbance (cattle grazing, past logging and agricultural clearing), and fire occurrence (through checking for presence of fire tracks).
For the spatial association between doum palm stages and all other tree species (exceeding 10 cm in diameter at breast height; hypothesis 4), data were also collected on the distance of each adult palm to other tree species, the azimuth of each tree to the nearest neighbour, and its slope.
Sex ratio of doum palm
For the sex ratio (hypothesis 5), the number of male and female doum palms was collected in each of the plot used for spatial pattern analyses.
Data analysis
Assessing impact of land use types on doum palm populations’ structure
We calculated the following dendrometric parameters:
-
Diameter (D) of the multi-stem doum palms:
\(D = \sqrt {\sum\nolimits_{i = 1}^{w} {dfi^{2} } }\) with w: number of stems and dfi: diameter of each stem.
-
Height (Ht) of the doum palms:
\(Ht = di * \left( { { \tan } \alpha_{ 1i} + { \tan } \alpha_{ 2i} } \right)\) with di: estimated distance of the falling tree and α1i and α2i: the higher and lower angles estimated with a clinometer SUUNTO.
Then, we calculated densities (i.e. the average number of trees per plot expressed in trees/ha) and displayed population structure (life stages) in both land use types. We used an analysis of variance (ANOVA) (Zar 1999) to depict possible difference in density of life stages between farmlands and BRP, and we used a log-linear analysis to test if the proportion of trees encountered in each life stage differed between land use types.
Densities were also estimated per height class and the distribution in height classes was displayed in the form of a histogram. To compare height class distributions, we used the coefficient of skewness of the distribution (Condit et al. 1998; Leakey et al. 2005). Spearman’s rank correlation was used to identify the environmental parameters and human disturbance that correlated with seedlings, juveniles and adult palm density. All variables evaluated were either continuous or ordinal (Gaoue and Ticktin 2009).
Assessing spatial patterns of doum palm life stages, spatial association among doum palm stages and other tree species
For spatial analysis, Arpent 1.3.d. software was used to convert distance, azimuth and slope data to the x,y coordinate of each tree (Lejeune 2001). We used the standardized Morisita index with a 95 % confidence interval to assess the global spatial pattern for doum palm life stages (Krebs 1999). The deviation from random expectation was tested using critical values of the Chi squared distribution with n − 1 degrees of freedom. Of the emerging techniques in spatial statistics, the most informative descriptor of pattern structure is the neighbourhood density function g(r) (Law et al. 2009) which describes the plant’s-eye perspective of neighbourhood density at an increasing distance r around a focal plant, averaged across the population sampled. Then, for each individual plot we used the pair-correlation function (Stoyan and Stoyan 1996) to test (1) whether observed samples can be considered as having a regular, random or aggregated pattern; and (2) whether the relationship between two types of sampled points suggests spatial association, repulsion, or independence. The estimate of g is a useful statistic summarizing one aspect of points’ clusters. Assuming the point process is stationary, we can define the cumulative distribution function of the nearest-neighbour distance for a typical point in the pattern as:
where K′(r) is the derivative of K(r), the reduced second moment function (aka “Ripley’s K function”; Ripley 1981) of the point process. The pair correlation g(r) is the probability of observing a pair of points separated by a distance r, divided by the corresponding probability for a Poisson process. If densities are independent at a given distance, then g(r) ∼ 1. Values of g(r) < 1 suggest inhibition between points; values of g(r) > 1 suggest clustering or attraction at distance r. The cross-type pair-correlation function is a generalization of the pair-correlation function to multitype point patterns. For a stationary multitype point process, the cross-type pair-correlation function between marks i and j is formally defined as (Baddeley and Turner 2005):
where \(K_{i,j}^{'}\) is the derivative of the cross-type K function Ki,j (r) of the point process. When a process of type i points was independent from one of type j, thus Kij (r) would be equal to \(\pi r^{2}\). For any marked point pattern in which the points of type i are independent of the points of type j, the theoretical value of the cross-type pair-correlation function is g i,j (r) = 1. The tests of spatial pattern/association were all constructed in the same way. The estimated function is compared to the theoretical function under the null hypothesis, H0 (complete spatial randomness for spatial pattern tests or spatial independence for spatial association tests), through a test statistic whose expected null-hypothesis value is 1 at all distances. Test significance is estimated through a Monte Carlo procedure (Barot et al. 1999). Rejection limits for the test are estimated as the simulation envelopes under H0, of spatial patterns of the same density as the observed sample. We used 499 simulations to compute tests at a 5 % confidence level (Diggle and Ribeiro 2007). In case that g(r) for a given scale r was outside the simulation envelopes, the null hypothesis was rejected. For a completely spatially random Poisson marked point process, p(r) = λiλj so gi,j(r) = 1. Indeed for any marked point pattern in which the points of type i are independent of the points of type j, the theoretical value of the cross-type pair correlation is gi,j(r) = 1.
For univariate point patterns, g(r) = 1 indicates randomness, while g(r) > 1 and g(r) < 1 indicate clumping and regularity, respectively. For a multitype point pattern, gi,j(r) = 1 indicates spatial independence, whereas gi,j(r) > 1 and gi,j(r) < 1 indicates a positive association (attraction) and a negative association (repulsion). Bivariate analyses assessed whether interactions among palm life stages and between palm life stages and other tree species occurred under a null hypothesis of spatial independence (Goreaud and Pélissier 2003).
Assessing doum palm sex ratio
Deviations of a sex ratio (male/female) from 1:1 were tested in each of the three stands using an exact binomial test. Statistical analyses were performed in R 3.1.0 (R Core Team 2013) using base functions and the packages vegan (Oksanen et al. 2007) and spatstat (Baddeley and Turner 2005) (Figs. 2, 3).

Spatial pattern of doum palm life stages (seedlings and adults) and other tree species in the three stands size of farmlands

Spatial pattern of doum palm life stages (seedlings and adults) and other tree species in the three stands size of the Biosphere Reserve of Pendjari
Results
Structural parameters and stand structure of doum palm in different land use types
Comparison of dendrometric traits revealed that adult palms diameters were significantly larger in BRP than in farmlands (Table 1; P < 0.001). In addition, density of adult palms and seedlings were significantly higher in BRP than in farmlands (Table 1; P < 0.001). However, no significant differences were observed for both tree height and density of juveniles (P > 0.05).
Comparison of life stage structure between land use types (Fig. 4) revealed that all doum palm life stage (seedlings, juveniles, and adults) densities were significantly higher in BRP than in farmlands (P < 0.001). However there was no significant difference between densities of juveniles in both land use types (P > 0.05).

Life stage distribution of doum palm in farmlands versus Biosphere Reserve of Pendjari in Benin. BRP Biosphere Reserve of Pendjari
As for doum palm height, comparison between farmlands and BRP revealed a coefficient of skewness of 0.1 for farmlands and 0.55 for BRP (Fig. 5) indicating a right skewed distribution. In addition, the density of doum palm was higher in the BRP than in farmlands, except for individuals of mean height = 2.75 m. No individual taller than 14.75 m was found in farmlands.

Frequency of doum palm height in farmlands versus Biosphere Reserve of Pendjari in Benin
There was a negative correlation between doum palm densities (juveniles and adults) and both human disturbance and fire (Table 2). As for seedlings of the species, their density was negatively correlated with the occurrence of fire (Table 2).
Spatial patterns of doum palm life stages, spatial association among doum palm stages and other tree species
Assessment of the global spatial patterns of palm life stages and other tree species showed a random distribution (Table 3). However, assessment of univariate point pattern revealed different scenarios (Table 4; Fig. 6). The pair-correlation function suggested a global random distribution for all palm life stages albeit with a weak aggregation within a 0–10 m radius in farmlands. This result holds for all three stands in farmlands. However, in the BRP, a strong aggregated distribution was observed for seedlings between 0 and 5 m, whereas all other palm life stages suggested a global random distribution in all three stands (Table 5).

Results of the point pattern analyses for seedlings (a), adults × seedlings (b) and adults × trees (c) in stand 3 in BRP. The horizontal dashed line shows the pair-correlation function expected from a Poisson process. Dotted lines are approximate 95 % confidence envelopes for the null hypothesis, obtained from 499 independent simulations
When considering spatial association between palm life stages, there was global independence for all stand sizes in the farmlands. For the BRP, the same trend was also observed between adults (females and males), and between adults and seedlings. However a spatial association was observed between females and seedlings in all stands.
Other tree species were randomly distributed in both land-use types, and results suggested no association with all palm life stages (Tables 4, 5).
Sex ratio of doum palm
The sex-ratio of doum palm was not significantly different from 0.5 (P > 0.05) in the two land use types, although adult male palms dominated some plots in the farmlands (Table 6). This result indicated an equal representation of both sexes in the two land use types.
Discussion
Land use influence on the stand structure of doum palm
Our study investigated differences between the population structure of doum palm in farmlands and the BRP. Density of adults and seedlings of doum palm were higher in BRP than in farmlands. In addition, adult doum palms were larger in BRP than in the farmlands. These results supported the hypothesis that practices in the farmlands have a negative impact on doum palm and suggest the potential for conservation of the species in BRP. Unfortunately, this opportunity should not be taken for granted, as human disturbance beyond biosphere reserves influence biodiversity within them (Hansen and DeFries 2007). As stated by Kiringe et al. (2007), weak enforcement and ineffective management plague many of Africa’s current protected areas. As such, effectiveness of PA in biodiversity conservation is questionable.
The negative influence of farmlands on the doum palm stand structure could be explained by the presence of traditional agriculture; hence, the vegetation is often cleared with local tools and burned for crop growth. This phenomenon is also the case for other palm species occurring in agroforestry systems (Barot et al. 2000; Laris and Wardell 2006). Consequently, seedlings of the palm species rarely persist in those systems. Some adult trees are intentionally preserved because they provide fruits for local people, as is the case for other agroforestry trees (Djossa et al. 2008; Schumann et al. 2011). These fruits, once fallen on the ground, could also help in recruitment of seedlings of the species. Nevertheless, the land preparation for agriculture constitutes a permanent threat for the seedlings of doum palm in farmlands. Our results match previous findings stipulating that individuals of some agroforestry tree species in young age classes occurred at lower numbers in farmlands when compared to BRP (Kelly et al. 2004; Schumann et al. 2011).
No significant differences were observed when comparing the height of the doum palm in both land use types. This could be explained by the fact that reduction in the availability of croplands lead farmers to shorten the fallows cycle. As H. thebaica is a long-life species, the same trees are encountered along time with little recruitment if not logged for other purposes (Kahn and Luxereau 2008). This constitutes a rescue strategy for the species in its distributional areas.
Land use influence on doum palm spatial patterns
A random distribution was observed for doum palm life stages, albeit with a weak aggregation between 0 and 10 m in farmlands, thus partially supporting our second hypothesis that doum palm shows clumped patterns in BRP, but random distributions in farmlands. The same pattern has been observed for other palm and dioecious species (Barot et al. 1999; Gibson and Menges 1994; Sterner et al. 1986). The species’ spatial pattern could be attributed to patchy habitat variation, to the limited dispersal of seeds, or to neutral processes that disregard species-specific traits (Harms et al. 2001; Hubbell 2001). This weak aggregation of seedlings can also be related to human influence on the landscape. Indeed land preparation for agricultural needs leads to high seedling death and modifying the species’ natural structure. However in the BRP, the species showed aggregation of offspring, as the ripe fruits tend to fall close to the mother tree (i.e. gravity-dispersed) and grow successfully due to physical features (i.e. substrate type, moisture, light, temperature). Homogeneity of the macro environment may also permit the establishment of the species with varying degrees of success. This phenomenon supports the hypothesis that aggregation corresponds with absence or minimum disturbance and diminishes with higher disturbance.
Our results partially supported the idea of spatial association between seedlings and adult palms (but only for female in the BRP). These findings match those from other studies that have shown a positive association between females and seedlings in dioecious species (Barot and Gignoux 2003; Condit et al. 2000; Montesinos et al. 2007). This proves true as seed dispersal is constrained by a number of ecological factors, including density and aggregation of individuals, and effects of habitat heterogeneity (Boll et al. 2005; Vormisto et al. 2004). However, distribution of seedlings was independent from those of other palm life stages in both land use types. This could also be explained by human modification of the landscape, leading to disturbance of the species’ normal distribution.
Our results support the hypothesis that other tree species showed no association with all doum palm life stages in both land use types. This suggests that human factors do not have a particular influence on the spatial structure of doum palms and other tree species in both land use types. In addition, competition for light or food and natural selection among species could lead to absence of attraction between competing species.
Land use impacts on doum palm sex ratio
The sex ratio is significantly male-biased in the plot 1 in farmland. However, in larger plots the sex ratio did not significantly differ from 0.5. These findings do not support our hypothesis that the sex ratio of doum palms is female biased. This result may suggest a high level of pressure on the species in plot 1. In fact, field observations revealed an intense agricultural practice in the area, leading to the cutting of almost all trees. In other areas, adult individuals of both sexes are cut, leading to an equal number of males and females in the different land use types. However the ratio of felling should not be occulted.
Implications of the study
Our spatial analysis provides evidence that disturbance may contribute to an irregular distribution pattern of doum adult palms, leading to increased pressure on the species. Another important aspect is the fragmentation of the landscape. Doum palm is located in rural areas, less likely to experience rapid urbanization. However, as noticed by the Food and Agricultural organization in its report, part of the BRP is converted to agricultural lands, due to rapid population growth in surrounding areas and because of a reduction in soil fertility. This could be an important threat to the conservation of doum palm, although some remnant individuals are kept in agroforestry systems. Thus, only agricultural practices that maintain a sufficient number of male and female individuals of doum palm can significantly contribute to its persistence in agroforestry systems.
Conclusion
Comparison of population structure between land use types indicated negative impacts of practices in farmlands on doum palm stand structure. The low density of juveniles in both land use types suggested recruitment problems for this development stage of the species, thus raising doubts for long term conservation of the species. Our results also showed an overall tendency towards a less aggregated pattern throughout the doum palm life stages in both land use types. A spatial association is noticed between seedlings and female adults, but only in the biosphere reserve. Although adult male palms tended to dominate some plots in the farmlands, sex-ratio was not significantly different from 0.5 in all sampled stands.
Overall, this study contributed novel information about the structure and spatial patterns of doum palm life stages and trees in stands. We recommend that permanent plots should be set up and data collected for a longer period of time (i.e. at least 5 years) to effectively assess the population status of this species. This approach could help to identify potential processes that affect different doum palm life stages and further define more reliable long term conservation strategies for doum palms.
References
Azihou AF (2013) Ecology of isolated trees in tropical savannas: modelling of plant species distribution and colonization of new habitats through long-distance dispersal and facilitation. PhD thesis. University of Abomey-Calavi, Republic of Benin, 184 p
Baddeley A, Turner R (2005) Spatstat: an R package for analyzing spatial point patterns. J Stat Softw 12:1–42
Barot S, Gignoux J (2003) Neighbourhood analysis in the savanna palm Borassus aethiopum: interplay of intraspecific competition and soil patchiness. J Veg Sci 14:79–88
Barot S, Gignoux J, Menaut JC (1999) Demography of a savanna adult palm: predictions from comprehensive spatial pattern analyses. Ecology 80:1987–2005
Barot S, Gignoux J, Vuattoux R, Legendre S (2000) Demography of a savanna adult palm in Ivory Coast (Lamto): population persistence and life-history. J Trop Ecol 16:637–655
Bhagwat SA, Willis KJ, Birks HJB, Whittaker RJ (2008) Agroforestry: a refuge for tropical biodiversity? Trends Ecol Evol 23:261–267
Boll T, Svenning JC, Vormisto J, Normand S, Grandez C, Balslev H (2005) Spatial distribution and environmental preferences of the piassaba palm Aphandra natalia (Arecaceae) along the Pastaza and Urituyacu rivers in Peru. For Ecol Manag 213:175–183
Condit R, Sukumar R, Hubbell SP, Foster RB (1998) Predicting population trends from size distributions: a direct test in a tropical tree community. Am Nat 152:495–509
Condit R, Watts K, Bohlman SA, Perez R, Foster RB, Hubbell SP (2000) Quantifying the deciduousness of tropical forest canopies under varying climates. J Veg Sci 11:649–658
Dawson W, Burslem DF, Hulme PE (2009) The suitability of weed risk assessment as a conservation tool to identify invasive plant threats in East African rainforests. Biol Conserv 142:1018–1024
Diggle P, Ribeiro PJ (2007) Model-based geostatistics. Lancaster University, Lancaster
Djossa BA, Fahr J, Wiegand T, Ayihouénou B, Kalko E, Sinsin B (2008) Land use impact on Vitellaria paradoxa CF Gaerten. stand structure and distribution patterns: a comparison of Biosphere Reserve of Pendjari in Atacora district in Benin. Agrofor Syst 72:205–220
Fanshawe D (1966) The Doum palm-Hyphaene thebaica (Del.) Mart East African. Agric For J 32:108–116
Gaoue OG, Ticktin T (2008) Impacts of bark and foliage harvest on Khaya senegalensis (Meliaceae) reproductive performance in Benin. J App Ecol 45:34–40
Gaoue OG, Ticktin T (2009) Fulani Knowledge of the Ecological Impacts of Khaya senegalensis (Meliaceae) Foliage Harvest in Benin and its Implications for Sustainable Harvest. Econ Bot 63:256–270
Gelfand AE, Diggle P, Guttorp P, Fuentes M (2010) Handbook of spatial statistics. CRC Press/Taylor and Francis Group, Boca Raton
Gibson DJ, Menges ES (1994) Population structure and spatial pattern in the dioecious shrub Ceratiola ericoides. J Veg Sci 5:337–346
Giffard PL (1966) Le palmier doum (Hyphaene thebaica Mart.). Bois For Trop 106:3–11
Glèlè Kakaï R, Akpona T, Assogbadjo AE, Gaoué OG, Chakeredza S, Gnanglè PC, Mensah GA, Sinsin B (2011) Ecological adaptation of the shea butter tree (Vitellaria paradoxa CF Gaertn.) along climatic gradient in Bénin. West Africa. Afr J Ecol 49:440–449
Goreaud F, Pélissier R (2003) Avoiding misinterpretation of biotic interactions with the intertype K12-function: population independence vs. random labelling hypotheses. J Veg Sci 14:681–692
Haarmeyer DH, Schumann K, Bernhardt-Römermann M, Wittig R, Thiombiano A, Hahn K (2013) Human impact on population structure and fruit production of the socio-economically important tree Lannea microcarpa in Burkina Faso. Agrofor Syst 87:1363–1375
Hansen AJ, DeFries R (2007) Ecological mechanisms linking protected areas to surrounding lands. Ecol Appl 17:974–988
Harms KE, Condit R, Hubbell SP, Foster RB (2001) Habitat associations of trees and shrubs in a 50-ha Neotropical forest plot. J Ecol 89:947–959
Haugo RD, Hall SA, Gray EM, Gonzalez P, Bakker JD (2010) Influences of climate, fire, grazing, and logging on woody species composition along an elevation gradient in the eastern Cascades, Washington. For Ecol Manag 260:2204–2213
Hsu B, Coupar IM, Ng K (2006) Antioxidant activity of hot water extract from the fruit of the Doum palm, Hyphaene thebaica. Food Chem 98:317–328
Hubbell SP (2001) The unified neutral theory of biodiversity and biogeography (MPB-32), vol 32. Princeton University Press, Princeton
INSAE (2015) RGPH4: que retenir des effectifs de population en 2013? Direction des études démographiques. Ministère du Développement de l’Analyse Economique et de La Prospective. Cotonou, République du Bénin
Isaac ME, Dawoe E, Sieciechowicz K (2009) Assessing local knowledge use in agroforestry management with cognitive maps. Environ Manag 43:1321–1329
Jahiel M (1993) Le palmier doum: Hyphaene thebaica (L.) (Mart.). Le Flamboyant 28:4–10
Jorgensen SE, Fath B (2008) Encyclopedia of ecology, five-volume set: online version, vol 1. Newnes, Oxford
Kahn F, Luxereau A (2008) Doum palm habit and leaf collecting practices in Niger. Palms 52:23–29
Kelly BA, Bouvet J-M, Picard N (2004) Size class distribution and spatial pattern of Vitellaria paradoxa in relation to farmers’ practices in Mali. Agrofor Syst 60:3–11
Kiringe JW, Okello MM, Ekajul SW (2007) Managers’ perceptions of threats to the protected areas of Kenya: prioritization for effective management. Oryx 41:314–321
Krebs CJ (1999) Ecological methodology, vol 620. Benjamin Cummings, Menlo Park
Laris P, Wardell DA (2006) Good, bad or ‘necessary evil’? Reinterpreting the colonial burning experiments in the savanna landscapes of West Africa. Geogr J 172:271–290
Law R, Illian J, Burslem DF, Gratzer G, Gunatilleke C, Gunatilleke I (2009) Ecological information from spatial patterns of plants: insights from point process theory. J Ecol 97:616–628
Leakey R, Pate K, Lombard C (2005) Domestication potential of Marula (Sclerocarya birrea subsp caffra) in South Africa and Namibia: 2. Phenotypic variation in nut and kernel traits. Agrofor Syst 64:37–49
Lejeune P (2001) Arpent 2.0, logiciel de saisie de données d’arpentage – Guide d’utilisation. FUSAGx. Note Tech For 5:1–12
Lykke AM (1998) Assessment of species composition change in savanna vegetation by means of woody plants’ size class distributions and local information. Biodivers Conserv 7:1261–1275
Montesinos D, Verdú M, García-Fayos P (2007) Moms are better nurses than dads: gender biased self-facilitation in a dioecious Juniperus tree. J Veg Sci 18:271–280
Moussa H, Margolis HA, Dubé P-A, Odongo J (1998) Factors affecting the germination of doum palm (Hyphaene thebaica Mart.) seeds from the semi-arid zone of Niger, West Africa. For Ecol Manag 104:27–41
Oksanen J, Kindt R, Legendre P, O’Hara B, Stevens MHH, Oksanen MJ, Suggests M (2007) The vegan package Community ecology package, pp 631-637
Ouédraogo A, Glèlè Kakaï R, Thiombiano A (2013) Population structure of the widespread species, Anogeissus leiocarpa (DC.) Guill. & Perr. across the climatic gradient in West Africa semi-arid area. S Afr J Bot 88:286–295
Pannell JR (2002) The evolution and maintenance of androdioecy. Annu Rev Ecol Syst 33:397–425
R Core Team (2013) R: a language and environment for statistical computing. R. foundation for statistical computing, Vienna
Ripley BD (1981) Spatial statistics. Wiley, New York
Sapir Y, Mazer S, Holzapfel C, Erik J, Brian F (2008) Sex ratio. Encyclopedia of ecology I. Elsevier, Amsterdam, pp 3243–3248
Schumann K, Wittig R, Thiombiano A, Becker U, Hahn K (2011) Impact of land-use type and harvesting on population structure of a non-timber forest product-providing tree in a semi-arid savanna, West Africa. Biol Conserv 144:2369–2376
Sinsin B, Matig OE, Assogbadjo AE, Gaoue OG, Sinadouwirou T (2004) Dendrometric characteristics as indicators of pressure of Afzelia africana Sm. dynamic changes in trees found in different climatic zones of Benin. Biodivers Conserv 13:1555–1570
Sterner RW, Ribic CA, Schatz GE (1986) Testing for life historical changes in spatial patterns of four tropical tree species. J Ecol 74:621–633
Stoyan D, Stoyan H (1996) Estimating pair correlation functions of planar cluster processes. Biometrical J 38:259–271
Teklehaimanot Z (2003) Improved management of agroforestry parkland systems in Sub-Saharan Africa. School of Agricultural and Forest Sciences, University of Wales, Bangor
Turner MG (1989) Landscape ecology: the effect of pattern on process. Annu Rev Ecol Syst 20:171–197
Vormisto J, Svenning JC, Hall P, Balslev H (2004) Diversity and dominance in palm (Arecaceae) communities in terra firme forests in the western Amazon basin. J Ecol 92:577–588
Zar JH (1999) Biostatistical analysis. Pearson Education, India
Acknowledgments
This work was funded by the University of Abomey-Calavi through WILD-PALM Project. Additional funding was provided by International Foundation for Science through a grant (Grant no.: D/5612-1, Stockholm, Sweden) and the Rufford Small grants (Grant no. 17952-1) provided to Rodrigue Idohou. We are also grateful to Prof Arturo Ariño (University of Navarra, Spain), Prof Andrew Townsend Peterson and Lindsay Campbell (University of Kansas, USA) for thorough language and style corrections. We also acknowledge high contribution of the editor and anonymous referees.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Idohou, R., Assogbadjo, A.E., Azihou, F. et al. Influence of the landscape context on stand structure and spatial patterns of the doum palm (Hyphaene thebaica Mart.) in the Republic of Benin (West Africa). Agroforest Syst 90, 591–605 (2016). https://doi.org/10.1007/s10457-016-9920-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-016-9920-4