
Abstract
During the spring of 2006, three willow varieties (SV1, SX67 and 9882-41) were established on marginal land in an agroforestry tree-intercropping arrangement where plots of short rotation willows were planted between rows (spaced 15 m apart) of 21-year-old mixed tree species. As a control, the same varieties were established on an adjacent piece of land without established trees (conventional willow system). This study investigated the magnitude of carbon pools, fine root and leaf biomass inputs and clone yields in both the tree-based intercropping (agroforestry) and conventional monocropping systems. Willow biomass yield was significantly higher in the agroforestry field (4.86 odt ha−1 y−1) compared to the conventional field (3.02 odt ha−1 y−1). In both fields, varieties SV1 and SX67 produced higher yields than the variety 9882-41. Willow fine root biomass in the top 20 cm of soil was significantly higher in the intercropping system (3,062 kg ha−1) than in the conventional system (2,536 kg ha−1). Differences in fine root biomass between clones were similar to that observed for differences in biomass yield: SV1 > SX67 > 9882-41. Leaf input was higher in the intercropping system (1,961 kg ha−1) than in the conventional system (1,673 kg ha−1). Clonal differences in leaf inputs followed the same trends as those for root biomass and yield: SV1 > SX67 > 9882-41. Soil organic carbon was significantly higher in the agroforestry field (1.94 %) than in the conventional field (1.82 %). A significant difference in soil organic carbon was found between the three clones: soils under clone 9882-41 had the lowest soil organic carbon at 1.80 %.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
A general principle of tree-crop complementarities is to grow trees with crops such that each takes advantage of spatially- or temporally-separated resources (Ong and Huxley 1996; Gordon and Newman 1997). Given the potential for competition between trees and crop plants, it is important to maximize complementary interactions and minimize any competitive interactions (Thevathasan and Gordon 2004). One way to achieve this is by understanding parameters such as the minimal distance required between intercropped tree rows and crop plants to avoid significant competition for light and nutrients.
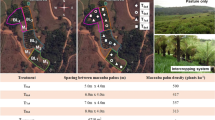
Results from experiments conducted at the University of Guelph Agroforestry Research Station (GARS) over the last 24 years suggest that two distinct zones exist across a 15 m wide tree-intercropping alley with temperate mixed species (Thevathasan and Gordon 2004; Clinch, 2008). The first zone—the competitive zone is the area within 2 m of tree rows. The second zone—the complementary zone is the remaining area in the centre of the alley, which is ~11 m wide. The competitive zone is characterized by direct competition for nutrients, moisture and light. The complementary zone is characterized by favourable growing conditions, where the following processes are enhanced: nutrient cycling, nitrogen mineralization, soil organic carbon addition and earthworm activity. In addition, the complementary zone has lower soil temperature and higher moisture availability, the latter as a result of less evapotranspiration and carbon assimilation (Reynolds et al. 2007; Thevathasan and Gordon 2004; Clinch et al. 2009). The two zones are depicted in Fig. 1.

Schematic showing the ‘competitive’ and the ‘complementary’ zones in the tree-based intercropping field at the GARS, Guelph, Ontario, Canada
The Ontario greenhouse industry, worth about US$ 2.3 billion per year, is seeking alternative energy sources to oil and natural gas since about 25 % of greenhouse operating costs are incurred for heating (Picchi et al. 2006). Emissions from greenhouses are currently unregulated, but if emission standards and/or carbon trading strategies develop in the future, a move towards cleaner bioenergy from short rotation woody crops (SRWC) and other wood residues would become economically attractive. This study was designed to investigate the potentials and possibilities of growing hybrid willow as a crop in the complementary zone of a tree-based intercropping system in southern Ontario, Canada in order to maximize woody biomass production for bioenergy. Other concurrent studies are investigating the economic and life-cycle feasibility of using this bioenergy source for the greenhouse industry. The results from the current study could serve as a “decision support tool” for landowners practicing tree-based intercropping systems in temperate North America.
Materials and methods
Field location and design
The experimental field sites are located at the University of Guelph’s Agroforestry Research Station in Guelph, Ontario (latitude 43°32′28″ N, longitude 80°12′32″ W). The soils are classified as Gray Brown Luvisols with a fine sandy loam texture, and a summary of soil characteristics measured in both experimental fields at the onset of the experiment is given in Table 1. Willow plots were located in two fields, one with tree-based intercropping (agroforestry field), and one without trees (the control/monocrop field). The area of the agroforestry field used for this study consisted of 21-year-old mixed tree species (predominantly Juglans nigra L. with some Quercus rubra L., Fraxinus Americana L. and Robinia pseudoacacia L.) planted in 370 m long rows with 6 m spacing between trees in a row, and 15 m width between tree rows (the crop alley or alleyway). Tree rows were oriented northwest to southeast. Willow plots were located within the alleyways between the large tree rows with a 2 m buffer on either side, placing them in the central 11 m (complementary zone) of the alleyways. The control field had no established trees, and plots in both fields were separated by at least a 2 m buffer.
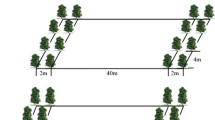
Plots of three willow varieties (Salix dasyclados—SV1, S. miyabeana—SX67 and S. purpurea—9882-41), obtained from State University of New York-Environmental Science and Forestry (SUNY-ESF), Syracuse, New York, were randomly arranged with four replications within each field, for a total of 12 plots in each field in 2006. Plot dimensions were 10 × 50 m with five double rows oriented along the long axis. Each double row consisted of two single rows of willow plants; the single rows were separated by a space 0.75 m in length (smaller inter-row), and each double row was separated from the next double row by 1.5 m (larger inter-row). The spacing between cuttings in each row was 0.55 m, resulting in a planting density of approximately 20,000 stems ha−1 (Fig. 2).

Willow row configuration within the ‘complementary’ zone
Willow biomass yield
In late fall, 2009, two random samples (each consisting of four trees) per plot were harvested at about 10 cm above the soil surface resulting in a total of eight samples per clone (variety) (2 samples × 4 replications = 8 samples). The sample area was 3.105 m2. Willow biomass samples were oven dried at 65 °C for 5 days in order to obtain dry biomass yields per unit land area.
Willow root biomass
Four randomized core samples per plot were taken in June, 2009: two in the large inter-row (1.5 m) and two in the smaller inter-row (0.75 m) (Fig. 2). Each sample was divided into four subsamples, in order to study the root distribution in the 0–5, 5–10, 10–15 and 15–20 cm soil layers. Soil samples were stored at 4 °C until washing, in order to prevent root decomposition. Roots were separated from the soil by manually washing and using sieves and tweezers, and then dried in an oven at 65 °C for 2 days.
Willow leaf input
Forty-eight leaf litter traps were manufactured and established in the two fields. Two random traps per plot were used, one in the larger inter-row and one in the smaller inter-row. The traps were 30 cm in diameter and 50 cm high. Leaves were collected every 2 weeks, from June to November, 2009. Leaves collected from June 5th to September 11th corresponded to leaf turnover during the growing season, whereas leaves collected from September 11th to November 20th corresponded to leaf drop during the fall season.
Soil C determination
Soil samples were collected in 2006, prior to the commencement of the experiment in order to obtain baseline values and in 2009 just before harvest, from the 0 to 20 cm depth using a soil auger and were analyzed for total C, inorganic and organic C using a Leco CR-12 carbon analyzer. Sample preparation was done using standard procedures as outlined in Carter and Gregorich (2008).
System carbon pools
Carbon pools in both systems were calculated from the data collected from this study. The aboveground C pool was calculated from the standing willow biomass just before harvest; three years of growth in 2009. The belowground C pool was calculated from the root biomass. The soil organic C pool was quantified for a soil weight of 2,000,000 kg as being the weight of one hectare furrow soil (20 cm depth).
Statistical analysis
All variance analyses were conducted using PROC MIXED in SAS v.9.1 (SAS Institute, Cary NC, USA) with a Type I Error rate of α = 0.05. The ‘repeated’ statement (corrected for autocorrelation using the TYPE = AR(1) statement) was used for repeated sampling (leaf biomass collection over time). Assumptions of homoscedasticity were tested by examining plots of residuals, and PROC UNIVARIATE was used to ensure residuals had a mean of zero. A Shapiro–Wilk test (α = 0.05) was used to test whether residuals followed a normal distribution, and log or square-root transformations were used to normalize residuals where necessary. Lund’s test for outliers was used to detect outliers in all datasets (Lund 1975). For data sets with n < 30 (foliar carbon) significant differences between means were determined using PROC TTEST. Correlation and regression analyses were done using PROC CORR and PROC REG in SAS. Plot average data for organic carbon was correlated with willow leaf input.
Results
In order to validate that both fields consisted of similar soil characteristics, complete soil analysis was done prior to the establishment of the test plots in 2006 (Baseline data). The results are presented in Table 1.
Willow biomass yield
Willow biomass yields are presented in Table 2.
Significant yield differences were observed between the two fields (agroforestry > control), and also between the three clones (SV1 = SX67 > 9882-41), when both fields were analyzed together. These differences in yields and possible causes, given that the baseline soil parameters were similar for both sites (Table 1), are discussed in the discussion section.
Willow root biomass
Root biomass (Fig. 3) was significantly different between the two fields (p < 0.05). In the top 20 cm of soil, root biomass was 3,062 kg ha−1 in the agroforestry field, and 2,536 kg ha−1 in the control field. The growth promoting parameters in the complementary zone, as indicated in Fig. 1, may have contributed towards the enhanced belowground root growth in the agroforestry field.

Willow root biomass distribution in the agroforestry field and monocropping field, three years after coppice in 2009, GARS, Ontario, Canada
Leaf biomass inputs to soil
Leaf litter inputs were found to be significantly different (p < 0.05) between the tested fields. In the agroforestry field, willow leaf input was 1,961 kg ha−1 y−1, and 1,673 kg ha−1 y−1 in the control field, from June to November, 2009. But in the agroforestry field, there was also leaf input from the larger intercropped trees, estimated at 450 kg ha−1 y−1 (this estimation does not represent the total leaf input from these trees, but represents leaf input collected within the sample plots, 2–6 m from the tree row).
Soil organic carbon
ANOVA on soil organic carbon data (Table 3) showed a significant variation between the two tested fields. The soil organic carbon in the agroforestry field was significantly higher than in the control or monocropped field. This is interesting to note as the baseline soil organic carbon, measured in 2006 at the onset of this experiment, did not show any significant differences between the tested sites. Annual addition of willow leaf biomass at the rate of almost 2 odt ha−1 y−1 coupled with leaf addition from matured trees in the rows could have contributed towards this increase in soil organic carbon.
Carbon pools
Quantified C pools, as described in the Materials and methods, are given in Table 4.
Carbon pools for both sites were calculated in order to study the trend of carbon sequestration over time under willow biomass production systems. Similar carbon pool values for the agroforestry site can be found in Peichl et al. 2006.
Willow mechanical harvest, 2009
In December 2009, both test sites were harvested with Anderson Biobaler harvester, the very first Canadian made willow bale harvester (Fig. 4).

Anderson biobaler harvester being operated by a 100 kW tractor, GARS, Guelph, Ontario, Canada
The average wet bale weight was around 350 kg with a moisture content of 52 % (data not presented). The bales were left in the field for winter drying during the winter of 2010 and were transported to a pelletizing plant in May 2010. At the time of transportation the bales contained an average moisture content of 11 % (data not presented). The bale harvest rate was 31 bales per hour. The biobaler should be operated with a tractor capable of delivering a minimum power of 100 kW.
Discussion
Tree-based intercropping is one among many temperate agroforestry land-use systems. Studies at the French National Institute for Agricultural Research (INRA) have identified three distinct tree-based intercropping systems based on tree densities per hectare (Dupraz and Liagre 2008). These intercropping systems are: a) stable agroforestry (20–50 trees ha−1), where crops can be grown until tree harvest. To have a stable agroforestry system, the distance between two tree rows should be at least twice the height of adult trees (30 to 40 m), b) changing agroforestry (50–200 trees ha−1), where at the end of tree growth, crop area may be reduced, or one will have to grow shade-tolerant crops, c) ephemeral agroforestry (more than 200 trees ha−1), where one can grow crops only during the first few years. This study was conducted in a “changing agroforestry” system with a tree density of 111 trees per hectare. During the early stages, annual crops were grown in the alleys. Due to the current interest in biomass for bioenergy, in this study, we utilized willow as an alternative crop that could be successfully grown in the alleys of a mature (21 year-old) tree-based intercropping system. This is a new temperate agroforestry concept—trees within trees—but willow is considered a crop due to the short harvest cycle of 3 years.
As discussed in the Introduction, past studies (Thevathasan and Gordon 2004; Reynolds et al. 2007; Clinch et al. 2009) have clearly demonstrated complementary growth promoting interactions in the middle of cropping alleys as influenced by the presence of mature trees along the tree rows (Fig. 1). It appears that these growth promoting interactions or processes have positively enhanced willow biomass yield in the agroforestry site when compared to the yield from the monocropping site (Table 2). The willow clonal varieties Salix dasyclados SV1 and S. miyabeana SX67 produced the highest yields overall (Table 2) and they may be recommended as promising biomass crops to be incorporated both in tree-based intercropping fields and in open fields. It is also important to note that such high biomass yields were obtained with only 15 kg N ha−1 (soil mineralization measured in both fields in June 2009—data not presented). This suggests that willow biomass crop can be grown without external inorganic fertilization at least during the initial years of establishment. As expected, root biomass followed a similar trend as observed in aboveground biomass yield. Root systems were more developed in the agroforestry field when compared with the control site (Fig. 3). It is believed that site-specific parameters not measured in this study but measured on these sites by other researchers, such as subsurface hydrology (Clinch et al. 2009), may have contributed to above and below ground biomass yield differences. In addition, the fine roots comprised a surprisingly large percentage of the total root biomass. Studies of hybrid poplar have shown that fine root biomass accounts for up to 60 % of the total root biomass in one-year-old trees (Dickman and Pregitzer 1992), about 40 % of the total root biomass in two-year-old trees (Friend et al. 1991), and 21–40 % of the total root weight in four-year-old hybrid poplar (Heilman et al. 1994). Based on these results, it was estimated that fine root biomass turnover was about 40 % of total root biomass, i.e. 1,200 kg ha−1 y−1 in the agroforestry field and 1,000 kg ha−1 y−1 in the control field.
This enhanced addition of below ground biomass in the agroforestry system significantly increased the soil organic carbon by 48 % in the agroforestry site but only by 27 % in the monocropping site, when compared with baseline soil organic carbon values for both sites (Tables 1, 3). Enhanced productivity in the complementary zone coupled with an associated soil C increase in the agroforestry field has resulted in a 12 % increase in the total C pool when compared with the total C pool of the conventional site over a period of three years (Table 4). It is expected that SRWC systems will last for seven harvest cycles or to a total period of 22 years from establishment (Abrahamson et al. 2010). Therefore, the total C pool in SRWC under agroforestry systems should potentially continue to increase in size as the system matures. This also emphasizes the need for further research on below ground C pools and the quantification of the belowground C sequestration process for short rotation willow biomass systems within the context of C credits and future trading scenarios.
Clinch et al. (2009) clearly demonstrated the availability of high soil moisture and low temperature in the complementary zone (Fig. 1). The belowground willow root distribution in Fig. 3 indicates that the majority of the roots in the agroforestry site were found to be in the top 10–12 cm, whereas in the control site, roots were evenly distributed with soil depth. This surface accumulation of roots in the agroforestry site may also contribute to the enhancement of C accumulation in the first 10–12 cm soil depth. In relation to marginal land reclamation, this potential surface C accumulation might prove to be extremely important.
Given the current political will and climate change mitigation strategies promoting bioenergy, SRWC production in mature tree-based intercropping systems in the temperate region could be a viable option. In addition, the development of commercial scale machinery such as, harvesters (Anderson biobaler) and planters (modified crop planters) in Canada in order to support such commercial scale biomass production units to produce bioenergy is also encouraging.
References
Abrahamson LP, Volk TA, Smart LB and Cameron KD (2010) Srub willow producer’s handbook, State University of New York http://www.esf.edu/willow/documents/ProducersHandbook.pdf Senior Research Associate. Accessed 04 Sep 2012
Carter MR, Gregorich EG (2008) Soil sampling and methods of analysis, 2nd edn. Canadian Society of Soil Science, CRC Press, Boca Raton, pp 228–229
Clinch RL (2008) Growing short-rotation woody crops in a tree-based intercropping system in southern Ontario. Dissertation, University of Guelph
Clinch RL, Thevathasan NV, Gordon AM, Volk TA, Sidders D (2009) Biophysical interactions in a short rotation willow intercropping system in southern Ontario, Canada. Agric Ecosyst Environ 131:61–69
Dickman DI, Pregitzer KS (1992) The structure and dynamics of woody plant systems. In: Mitchell CP, Ford JB, Hinckley T, Sennerby-Forsse L (eds) Ecophysiology of Short Rotation Forest Crops. Elsevier, London, pp 95–123
Dupraz C, Liagre F (2008) Agroforesterie - Des arbres et des cultures. Editions France Agricole, France
Friend AL, Scarascia-Mugnozza G, Isebrands JG, Heilman PE (1991) Quantification of two-year-old hybrid poplar root systems : morphology, biomass, and 14C distribution. Tree Physiol 8:109–119
Gordon AM, Newman SM (1997) Temperate agroforestry systems. CAB International Press, Wallingford
Heilman PE, Ekuan G, Fogle D (1994) Above- and below-ground biomass and fine roots of 4-year-old hybrids of Populus trichocarpa × Populus deltoides and parental species in short rotation culture. Can J For Res 24:1186–1192
Lund RE (1975) Tables for approximate test for outliers in linear models. Technometrics 17:473–476
Ong CK, Huxley P (1996) Tree-crop interactions: a physiological approach. CAB International, Wallingford
Peichl M, Thevathasan NV, Gordon AM, Huss J, Abohassan RA (2006) Carbon sequestration potentials in temperate tree-based intercropping systems, southern Ontario, Canada. Agrofor Syst 66:243–257
Picchi G, Gordon AM, Thevathasan NV (2006) Feedstock to furnace: bioenergy systems for the Ontario greenhouse industry. Misc. report, School of Environmental Sciences, University of Guelph
Reynolds PE, Simpson JA, Thevathasan NV, Gordon AM (2007) Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada. Ecol Eng 29:362–371
Thevathasan NV, Gordon AM (2004) Ecology of tree intercropping systems in the north temperate region: experiences from southern Ontario, Canada. Agrofor Syst 61:257–268
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Cardinael, R., Thevathasan, N., Gordon, A. et al. Growing woody biomass for bioenergy in a tree-based intercropping system in southern Ontario, Canada. Agroforest Syst 86, 279–286 (2012). https://doi.org/10.1007/s10457-012-9572-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-012-9572-y