
Abstract
Few studies have assessed whether patterns of resource partitioning among sympatric marine predators vary between different habitats. This type of data is important for understanding food web functioning as well as for supporting an ecosystem-based fisheries management plan. In this study, we used δ13C and δ15N stable isotope analysis to assess the relative isotopic niche area (i.e., which species has the largest and smallest isotopic niche area within a study site) and core isotopic niche overlap between 299 sharks of 11 co-occurring shark species that vary in size, diet and behavior in three coastal study areas in South Florida. Overall, results show that the relative isotopic niche area and patterns of core isotopic niche overlap between some sympatric shark species varied between sites, suggesting plasticity in resource use and competitive interactions between shark species (e.g., 85% of blacktip shark Carcharhinus limbatus standard ellipse area overlapped with the blacknose shark C. acronotus ellipse in Biscayne Bay, but there was no overlap between these species’ ellipses in nearby Florida Bay). Therefore, patterns of resource use and trophic interactions documented among species from one site may not be applicable to a neighboring location. These findings may have implications for food web models that incorporate trophic data from different areas for a species when local data are unavailable.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The study of predator–prey interactions is complicated by interspecific and intraspecific variation in the resource use of predators and in their competitive interactions with other species (Polis and Holt 1992). In some cases, co-occurring predator species may compete with one another for the same prey resources, but in other cases they may minimize competition by partitioning available resources (Schoener 1974). Determination of the diet and associated trophic relationships of sharks has been identified as research priorities for the conservation and management of shark species (Simpfendorfer et al. 2011), many of which are threatened (Dulvy et al. 2014).
The assessment of shark diets and trophic interactions is complex, because sharks are an ecologically diverse group with considerable variation in size, diet and trophic level both within and between species (Cortés 1999; Hussey et al. 2015; Frisch et al. 2016). In general, relatively few studies to date have compared the spatiotemporal variation in diet breadth and overlap of multiple co-occurring predators (Sanford et al. 2003; Wieters et al. 2008). Both resource partitioning (e.g., Papastamatiou et al. 2006) and competition for resources (e.g., Bethea et al. 2004) have been documented in communities of co-occurring elasmobranch fishes. Spatiotemporal variation in diets is common, but it is important to understand the degree of variation in diet and dietary overlap within and among shark species, and across habitats in order to create ecosystem-based fisheries management plans (Pikitch et al. 2004) and/or for generating ecological risk assessments of threatened species (Gallagher et al. 2012).
Stable isotope analysis provides a non-lethal means of studying aspects of elasmobranch resource use and trophic dynamics (see Shiffman et al. 2012 for a review). While there have been numerous diet and stable isotope studies involving marine predators, many focus on a single species or a single location, and there has been a relative lack of research which has considered how diet and trophic relationships among an assemblage of sympatric marine predators could vary intraspecifically across the range of those species (Sanford et al. 2003; Wieters et al. 2008). Stable isotopic signatures of consumers may vary spatially, temporally or by consumer body size, but few studies have examined all of these variables simultaneously (but see Davenport and Bax 2002; Heithaus et al. 2013; Matich and Heithaus 2014).
It is important to consider the possible effects of temporal variation in baseline isotopic signature (i.e., not distinct patterns of resource consumption among consumers) on the interpretation of consumer isotopic signatures (e.g., Goering et al. 1990; Harvey et al. 2002). Baseline isotopic signature may vary seasonally because of changing environmental conditions that influence the growth of primary producers (e.g., Goering et al. 1990), or there may be interannual variation (e.g., Anderson and Fourqurean 2003). Additionally, the size of an individual consumer may also influence its isotopic signature (more so than just variation in diet between different-sized consumers), and it is important to test for the effects of size (reviewed in Martínez del Rio et al. 2009).
The coastal waters of South Florida provide an opportunity to evaluate potential intraspecific (defined here as between populations of a species located in different habitats) differences in the trophic ecology of sympatric sharks across distinct nearby habitats. South Florida is home to a diverse community of sharks in several distinct habitats which differ in depth, bottom type, biodiversity and fishing pressure (Trent et al. 1997; Torres et al. 2006; Heithaus et al. 2007; Graham et al. 2016). While these habitats are geographically proximate and isotopically similar (described in the study sites section of methods) and some shark species regularly migrate much greater distances than the distance between study sites used here, these study sites are ecologically (in terms of relative biodiversity and biomass of prey and possible competitive interactions) and environmentally (in terms of bottom type and depth) distinct enough to be considered separate habitats. The present study used δ13C and δ15N stable isotope analysis to investigate patterns of relative isotopic niche area and core isotopic niche overlap among 11 species of co-occurring sharks within three distinct coastal areas of South Florida. Specifically, this study investigated the following research question: Do patterns of relative isotopic niche area, core isotopic niche overlap and other standard isotopic metrics between sympatric shark species vary across different habitats? We hypothesized that since prey availability is generally similar throughout South Florida and study species’ ranges overlap, co-occurring species patterns of relative isotopic niche area, core isotopic niche overlap and other trophic interactions would remain constant across different habitats.
Methods
Study sites
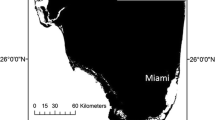
Sampling occurred year round (see supplementary materials for N by month and year) in three geographically proximate, isotopically similar, but ecologically and environmentally distinct coastal habitat in South Florida: Biscayne Bay, Florida Bay and the Florida Keys Reef, locations that vary in depth, bottom type, prey biodiversity and biomass, and fishing pressure (Fig. 1). These sites also vary slightly (but are generally similar) in isotopic signature of primary producers and prey species (see Kieckbusch et al. 2004 for Biscayne Bay; Chasar et al. 2005 for Florida Bay; and both Swart et al. 2005; Behringer and Butler 2006 for the Reef, with differences described briefly below).

GPS locations of South Florida sampling sites from this study. Larger stars indicate the general location of each study area, while smaller pluses indicate specific sampling sites within each study area
In the Biscayne Bay study area, sampling occurred year round from 2011 to 2013 on the sand, sponge flat and sea grass habitats within Biscayne National Park. Depth ranged from 5 to 20 m. The mean temperature in this area is 26 °C, with a range from 19 to 33 °C, while salinity is typically 30–40‰ but can occasionally decrease rapidly with freshwater pulses (Lirman and Cropper 2003). Twenty-three families of fishes reside within Biscayne Bay, and the most common are planktivores like Clupeidae, low-level predators like Gerridae and Cyprinodontidae, and small mesopredators like Lutjanidae and Haemulidae (Serafy et al. 2003). Although it is administered by the Federal National Parks Service, fishing within Biscayne National Park is governed by Florida’s state fishing regulations, with commercial and recreational fishing generally permitted except in seasonally closed areas (Graham et al. 2016). The average δ13C values of primary producers in this habitat have been found to range from − 27 (mangroves) to − 6.8 (epiphytic algae), though Kieckbusch et al. (2004) noted that 79% of all values were in the narrower range of − 21.9 to − 11.9 and that mangroves were not a major carbon source for the fish community. Low-level predatory teleosts such as pinfish (Lagodon rhomboides) have an average signature of (− 12.5, 9.5), and mesopredatory jacks (Caranx ruber) have an average signature of (− 10.5, 10.2), with a total isotopic range of the sample food web of approximately − 32 to − 7 δ13C and approximately 1–12 δ15N.
In the Florida Bay study area, sampling primarily occured in the shallow (3–5 m) and turbid waters of Everglades National Park, year round from 2009 to 2012. Due to pulses of freshwater input from the nearby Everglades, salinity in this location can vary from ~ 9 to ~ 35‰ (Fourqurean and Schrlau 2003), though measurements in this location taken by our laboratory were almost all over 28‰ (Hammerschlag, unpublished data). Mean temperature is 24.5 °C (range 20–28 °C) (Fourqurean and Robblee 1999). The benthos is primarily carbonate mud with some sea grass beds (Chasar et al. 2005). This region has high biomass of prey fishes (including planktivorous and low-level predatory fishes like Engraulidae, Blenniodei, Gobiidae and Clupeidae, and mesopredatory fishes from the family Sciaenidae, Thayer and Powell 1999), but these prey fishes are mostly relatively small, and there is relatively low fish biodiversity compared to coral reefs. Commercial fishing is prohibited within the waters of Everglades National Park (Graham et al. 2016), but recreational fishing is permitted with some restrictions on gear and bag limit. This habitat has been found to have a distinct isotopic range from Biscayne Bay, approximately − 18 to − 10 δ13C and approximately 1–14 δ15N, with pinfish having an average signature of (− 14.07, 9.77) and mesopredatory crevalle jacks (Caranx hippos) measured at (− 13.65, 11.45) (Chasar et al. 2005).
Lastly, for the Reef study area, sampling occurred on and around a 30-m-deep coral reef near Islamorada, Florida, year round from 2010 to 2013. Salinity in this region is a nearly constant 35‰ (Vaslet et al. 2012), and water temperature ranges from 21 to 29 °C (NOAA National Centers for Environmental Information). The Florida Keys coral reef system has hundreds of species of fish of all trophic levels (Ault and Johnson 1998), including non-elasmobranch mesopredators like grouper and barracuda in higher abundance than found in the other two study areas. Our sampling area is not part of one of the Florida Keys National Marine Sanctuary’s no-take zones (ecological reserves, sanctuary preservation areas or research natural areas), and therefore, general National Marine Fisheries Service fisheries regulations for US Federal waters apply (Graham et al. 2016). The average δ13C of coral zooxanthellae in the region has been found to be approximately (− 12, 5) (Swart et al. 2005, measured in 1996), while sea grass primary producers here have an average signature of (− 6.9, 2.9) while encrusting algae have an average signature of (− 14.9, 4.3) (Behringer and Butler 2006). The total isotopic range of the sampled food web was approximately − 18 to − 5 δ13C and approximately 0–8 δ15N (Behringer and Butler 2006), but this study did not include any top predators (though secondary consumer spiny lobster Panulirus argus had an average signature of (− 11.8, 7.3).
Sample collection and processing
Sharks of 11 species (Table 1) were captured by drum line as part of the University of Miami Shark Research and Conservation Program’s ongoing coastal shark survey (following Gallagher et al. 2014). For every shark captured, 5 mL of blood was drawn from the caudal vein. Sharks were measured (stretch total length, in centimeters), sexed and released. Whole blood (including plasma and red blood cells) was used for this analysis, and this tissue has a turnover time of approximately 8 months in chondrichthyans (Macneil et al. 2006).
Samples were stored in vacutainers (not containing anticoagulant) on ice in the field before being stored in a − 30 °C degree freezer in the laboratory. Stable isotope analysis was performed in the stable isotope laboratory at Boston University using glycine (− 34, 10.73) and peptone (− 14.73, 7.4) standards obtained from the National Bureau of Standards. Between 2 and 3 mg of wet weight whole blood was added to tins, which were dried overnight at 60 °C. Samples were combusted in a EuroVector Euro EA elemental analyzer and then passed through a GV Instruments diluter before being introduced to a GV Instruments IsoPrime isotope ratio mass spectrometer. One replicate per ten samples was run, any outliers were rerun, and laboratory standards of glycine and peptone were run once per 15 samples. Urea was not removed from blood samples. (See Kim and Koch 2012 for a description of this technique and how it impacts isotopic values, and see the limitations section of this manuscript for a discussion of how it could bias our results.) Ratios of stable isotopes were expressed as parts per thousand using standard δ notation.
Stable isotope analysis metrics
Isotopic ratios of carbon (C13/C12) and nitrogen (N15/N14) are enriched relative to a food source in a stepwise and relatively predictable manner (Peterson and Fry 1987), and are reported ‰. Carbon ratios primarily indicate the carbon source (e.g., pelagic vs. benthic, coastal vs. offshore, Minagawa and Wada 1984) and nitrogen ratios suggesting trophic level (i.e., those with higher N15/N14 ratios are feeding relatively higher in the food web, Post 2002). Analysis of these isotopic ratios can provide measurements of a species’ isotopic niche, as well as whether the isotopic niches of co-occurring species overlap. Although an isotopic niche provides insights into the combination of basal resources supporting a consumer’s diet and relative trophic level, isotopic niches are distinct from ecological or dietary niches because they do not provide direct insights into particular food sources (reviewed in Layman et al. 2012). While caution is needed in interpreting isotopic data relative to spatiotemporal variation in isotopic values at the base of food webs and dynamics of isotopic discrimination among species, isotopic niche overlap can provide insights into whether dietary overlap might occur.
Five metrics of relative isotopic niche breadth (i.e., comparisons between co-occurring species within the same study site) and area were calculated for each species in each study site with a sample size of greater than five individuals. This included calculations of (1) δ13C range, (2) δ15N range, (3) total occupied niche area, (4) standard ellipse area (SEAc) and (5) the mean distance to the centroid (CD), which are defined below. Both δ13C range and δ15N range are the differences between the smallest and largest δ13C and δ15N values within a sample, respectively (Layman et al. 2007). The total occupied niche area is the area of the convex hull surrounding the outermost points in a sample when those points are plotted on a δ13C vs. δ15N biplot, and this measures the total isotopic niche area of sampled individuals (Jackson et al. 2011). The small sample size-corrected standard ellipse area (SEAc) represents the core isotopic niche area of sampled individuals after factoring in maximum likelihood and is similar to a two-dimensional measurement of standard deviation (Jackson et al. 2011). Mean distance to the centroid (CD) measures trophic diversity within a sample (Layman et al. 2007). SEAc is used for overlap calculations and discussed in more detail because it is more suitable for the smaller sample sizes found for some species in this study than total occupied niche area (Jackson et al. 2011; Syväranta et al. 2013).
For δ13C range, δ15N range, total occupied niche area and standard ellipse area, larger values indicate relatively broader isotopic niches, likely suggesting a relatively generalist diet. A larger mean distance to the centroid indicates more isotopic dispersion (two-dimensional spread from the mean) within a group of individuals, likely suggesting more individual variation in diet among a population. Non-overlapping ellipses between sympatric species suggest the presence of resource partitioning between those species either in basal resources supporting diets, foraging locations or specific prey (e.g., Heithaus et al. 2013). However, the presence of overlapping ellipses does not necessarily suggest shared patterns of resource use between species. In the case of complex habitats with multiple carbon sources and many available prey resources, multiple distinct patterns of resource consumption can result in similar stable isotope signatures (reviewed in Layman et al. 2012).
Data analysis
All statistical analysis was performed using R version 3.1.2 (R Core Team 2013), including the SIAR and SIBER packages. For each habitat, the mean δ13C and δ15N values ± one standard deviation for each species were plotted (following Davenport and Bax 2002). SEAc and convex hull area was only calculated for species with an N greater than or equal to five within a study site.
Model selection
Following Tamburello et al. (2015), we used a model selection approach assessing the support of multiple GLMs to explore the influence of capture month, capture year and individual size (total length) on mean isotopic signature for species which had sufficient sample size across these variables. A species was considered to have sufficient sample size for the capture month variable if at least three sharks were sampled in at least three non-consecutive months, for the year variable if at least five sharks were captured in at least two different years (which sometimes meant comparing species caught in different months between years) and for the length variable if the range of sampled shark length was greater than 1 m. The relative support for each model was assessed using the Akaike information criterion (AIC), with lower AIC scores indicating a better-supported model. The log likelihood of each model was also calculated and reported, with higher values indicating a better-fitting model coefficient. The number of estimated model parameters, K, was also reported.
Results
Blood samples from 299 sharks of 11 species were analyzed, ranging in size from a 70 cm Atlantic sharpnose shark (Rhizoprionodon terranovae) to a 297 cm scalloped hammerhead shark (Sphyrna lewini) (Table 1). Species sampled also included blacknose (Carcharhinus acronotus), blacktip (C. limbatus), bull (C. leucas), Caribbean reef (C. perezi), great hammerhead (S. mokarran), lemon (Negaprion brevirostris), nurse (Ginglymostoma cirratum), sandbar (C. plumbeus) and tiger (Galeocerdo cuvier) sharks. Ten species were captured at both Biscayne Bay and the Reef, and seven species were captured at Florida Bay (Table 1). See supplementary materials for N by month and year for each species.
Biscayne Bay
Nurse sharks had the highest values for standard ellipse area (3.36), convex hull area (9.82) and δ15N range (4.63, Table 2) of any shark species captured at Biscayne Bay. Lemon, bull and sandbar sharks also had relatively high isotopic niche areas. Atlantic sharpnose sharks had the smallest isotopic niche, indicated by their standard ellipse area (1.05) and convex hull area (1.06). Lemon, great hammerhead, bull and sandbar sharks had the highest mean δ15N values of any species captured at Biscayne Bay, followed by blacktip and blacknose sharks (Fig. 2a, Table 2). The highest overlap between ellipses of sharks captured in Biscayne Bay was observed between blacknose and blacktip sharks, between sharpnose and tiger sharks, and between tiger sharks and nurse sharks (Fig. 3a, Table 3) The ellipses of the three species with the highest δ15N values (bull, lemon and tiger sharks) did not overlap at all (Fig. 3a). Nurse sharks had the largest CD (1.42, Table 2), and Atlantic sharpnose sharks had the smallest CD (0.69). Mean δ13C isotopic values ranged from − 14.9 ‰ (sandbar shark) to − 12.6 (lemon shark), and mean δ15N values ranged from 11.23 ‰ (Atlantic sharpnose shark) to 13.24 ‰ (sandbar shark).

a Average stable isotope ratios of shark species sampled in the Biscayne Bay study area ± 1 standard deviation. b Average stable isotope ratios of sharks species sampled in the Florida Bay study area ± 1 standard deviation. c Average stable isotope ratios of sharks species sampled in the Reef study area ± 1 standard deviation

a SIBER ellipses of shark species sampled in the Biscayne Bay study area. b SIBER ellipses of shark species sampled in the Florida Bay study area. c SIBER ellipses of shark species sampled in the Reef study area
Florida Bay
Bull and lemon sharks had the highest convex hull area (12.89 and 13.98, respectively) and standard ellipse area (5.51 and 4.96, respectively), demonstrating that these species have the largest isotopic niche area of any species captured in Florida Bay. Bull sharks had the highest δ15N values (Fig. 2b, Table 2). The most overlap between standard ellipses of sharks captured at the Florida Bay study area occurred between ellipses of lemon and blacktip sharks, lemon and bull sharks, blacktip and nurse sharks, and lemon and nurse sharks (Fig. 3b, Table 3). In Florida Bay, the ellipses of shark species with the highest δ15N values (bull shark, blacktip shark and lemon sharks) overlapped extensively with each other (Fig. 3b). Bull sharks had the largest CD (1.99, Table 2), and blacknose sharks had the smallest (0.66). Mean δ13C isotopic values ranged from − 17.38 ‰ (great hammerhead shark) to − 13.41 ‰ (nurse shark), and mean δ15N values ranged from 10.51 ‰ (tiger shark) to 12.44 ‰ (bull shark).
Reef
Bull and nurse sharks had the largest isotopic niche areas of any species captured in the Reef study area. Bull sharks had the highest standard ellipse area (6.1), δ13C range (6.1) and δ15N range (Table 2). Nurse sharks had the highest convex hull area (6.68), and the second-highest standard ellipse area (2.62), δ13C range (4.25) and δ15N range (4.71). Sandbar sharks had the smallest isotopic niche area of any species captured in the Reef study area, demonstrated by the comparatively low standard ellipse area (1.02) and convex hull area (2.76) of this species. Sandbar, scalloped hammerhead and bull sharks had the highest δ15N values of any species captured in the Reef study area (Fig. 2c). The highest overlap between standard ellipses of species captured in the Reef study area occurred between nurse and bull sharks, and between two of the species with the highest δ15N values (sandbar and bull sharks, Fig. 3c, Table 3). There was no overlap between the ellipses of sandbar sharks and tiger sharks, or between the ellipses of sandbar and nurse sharks (Fig. 3c, Table 3). Bull sharks had the largest CD (2.98, Table 2), and tiger and sandbar sharks had the smallest CD of 0.96. Mean δ13C isotopic values ranged from − 15.82 ‰ (scalloped hammerhead sharks) to − 10.41 ‰ (lemon shark), and mean δ15N values ranged from 9.7 ‰ (Atlantic sharpnose shark) to 14.43 ‰ (sandbar shark).
Comparisons of isotopic niche patterns between study areas
Bull, sandbar and hammerhead sharks had among the highest δ15N levels in each of the three study areas where they were caught (Fig. 2). Atlantic sharpnose sharks, the smallest species caught during this study, consistently had among the lowest δ15N levels in both habitats where they were found (Fig. 2a, c). Nurse sharks and blacknose sharks had consistently intermediate δ15N signatures relative to co-occurring species, and the δ15N of blacktip sharks varied from relatively high to intermediate (Fig. 2). The δ15N levels of lemon and tiger sharks were the fourth highest of co-occurring species in some study areas, but the eighth highest in others, demonstrating the most variability in relative δ15N of any species. (Although raw tiger shark δ15N value was similar between sample sites, how it compared with the δ15N values of co-occurring species varied).
Some pairs of species had consistent patterns of core isotopic niche overlap across different study areas. The ellipses of sandbar sharks and nurse sharks captured in the Reef study area did not overlap at all, and there was only a small area of overlap between the ellipses of these species captured in the Biscayne Bay study area. There was no standard ellipse overlap between blacknose and bull sharks captured in the Florida Bay study area, and there was only a very small area of overlap between the ellipses of these species captured in the Biscayne Bay study area. Tiger shark and bull shark ellipses did not overlap at all in either the Biscayne Bay or Reef study areas. In Biscayne Bay, only species with high δ15N values such as sandbar, bull and lemon sharks had notable unique isotopic niche space, the niches for the other species overlapped almost entirely with those of another species. In the Florida Bay area and at the Reef, only bull sharks had a notable amount of unique isotopic niche space.
Other pairs of species had widely variable patterns of core isotopic niche overlap between different study areas. Eighty-six percent of the standard ellipse of tiger sharks captured in Biscayne Bay overlapped with that of nurse sharks captured in Biscayne Bay. In contrast, among sharks captured in the Reef study area, only 17% of the tiger shark ellipse overlapped with the nurse shark ellipse. Eighty-five percent of the ellipse of blacktip sharks captured in Biscayne Bay overlapped with that of blacknose sharks, but in the Florida Bay study area there was no overlap between the ellipses of these species (Fig. 4). Ninety-two percent of the blacktip shark ellipse overlapped with that of lemon sharks, compared with 44% overlap between the ellipses of these species captured in the Biscayne Bay study area. Eighty-nine percent of the ellipse of nurse sharks captured in Florida Bay overlapped with that of lemon sharks, compared with 0% overlap between the standard ellipses of these species in Biscayne Bay (Fig. 4). Patterns of overlap were similar when using relatively larger convex hulls instead of small sample size-corrected SEAc values (Supplementary material); blacktip and blacknose sharks still showed notably higher isotopic niche overlap in Biscayne Bay than in Florida Bay, and nurse and lemon sharks still showed notably higher isotopic niche overlap in Florida bay than in Biscayne Bay.

Variable patterns in SIBER ellipse overlap for two selected species pairs between two habitats, highlighting the most significant shifts in patterns of isotopic niche overlap discovered in this study. a Blacknose (black) and blacktip shark (red) SIBER ellipse overlap in Biscayne Bay compared with b Blacknose and blacktip shark SIBER ellipse overlap in Florida Bay. c Lemon (black) and nurse (red) shark SIBER ellipse overlap in Biscayne Bay compared with d Lemon and nurse shark SIBER ellipse overlap in Florida Bay
Some shark species had consistent patterns of relative isotopic niche area compared with co-occurring species. Lemon and nurse sharks consistently had high ellipse areas relative to other species captured in the same study area. Bull sharks had the highest ellipse areas of any species captured in the Florida Bay (Fig. 3b) and in the Reef (Fig. 3c) study areas, but an intermediate ellipse area relative to species captured at Biscayne Bay (which was driven by a much smaller bull shark ellipse area in Biscayne Bay, rather than notable increases in the ellipse area of the other species, Fig. 3a). Blacknose, blacktip and tiger sharks had intermediate to relatively low ellipse areas in every study area where they were captured.
Model selection
In Biscayne Bay, there was sufficient sample size to test for the effects of all three variables on the isotopic signatures of lemon and nurse sharks and sufficient sample size to assess the effects of capture year and length on bull shark isotopic signature. In Florida Bay, the sample size was adequate to test for the effects of all three variables on lemon and nurse shark isotopic signatures. There was sufficient sample size to assess the influence of length and capture year on bull shark isotopic signature and to assess the influence of length and capture month on the isotopic signature of blacktip sharks. At the Reef, sample size was adequate to investigate the effects of all three variables on nurse sharks. These effects could not be evaluated for other shark species because of little variation in shark total length (most were large juvenile or adult sharks), capture month or capture year.
The best-supported model never included a month × year interaction effect, suggesting that month and year can be safely analyzed as separate factors. When all three sample areas were analyzed together, the best-supported model did not incorporate study area, but all three study areas were analyzed separately for easier comparison with the rest of this study’s results.
The best-supported model for nitrogen signature of lemon sharks captured in Biscayne Bay incorporated length (with larger sharks generally having a lower δ13C and a higher δ15N value) and capture year (with later years showing a generally higher δ13C value and lower δ15N value, Supplementary Material). For every other species captured in Biscayne Bay, the best-supported models incorporated only length (with larger sharks generally having a lower δ13C and a higher δ15N value). The best-supported models for the carbon and nitrogen signatures of species sampled in Florida Bay all incorporated a temporal factor, either month, year or both, with inconsistent patterns between species (Supplementary material). The best-supported models for the nitrogen signatures of bull, lemon and nurse sharks captured in Florida Bay also incorporated length, with larger individuals having a generally higher δ15N value. The best-supported model for the isotopic signatures of nurse sharks captured at the Reef incorporated capture month only (Supplementary material).
Discussion
Though studies of the diet of a single predator species or studies of dietary overlap between predators in a single location are relatively common, comparatively few prior studies have compared how patterns of resource use between multiple co-occurring marine top predators may vary among different locations within their range (Sanford et al. 2003; Wieters et al. 2008). The results of this study demonstrate that for some shark species in coastal South Florida, there appears to exist spatial variation in the relative size of core isotopic niches between sympatric species, as well as variation in patterns of core isotopic niche overlap between sympatric species. This leads us to reject our null hypothesis that these isotopic relationships would remain consistent across neighboring study areas where these species co-occur, and given that the isotopic signatures of food webs are similar between these study sites, it suggests that some patterns of resource use and competition among co-occurring shark species may differ between nearby habitats. Accordingly, ecological interactions between sympatric shark species may vary across their ranges, which should be considered when applying ecological data from one location to another location.
Similar patterns of spatial variation in isotopic and trophic interactions between sympatric predators have occasionally been noted in both freshwater and marine systems. A study of trophic interactions between communities of co-occurring freshwater teleost predators noted that the species with the relatively largest isotopic niche varied between nearby lakes (Hayden et al. 2013). For example, in Lake Roppi, SEAc values were 2.4 for whitefish (Coregonus lavaretus) and 4.7 for ruffe (Gymnocephalus cernuus), but in lake Oiko, whitefish had a larger standard ellipse area than ruffe (2.1 vs. 1.7, Hayden et al. 2013), which was attributed to different resource use patterns of the same species between these lakes despite similar prey availability and total system isotopic range in both lakes. A similar pattern was discovered in a marine system, where stomach content analysis showed that gray reef sharks (Carcharhinus amblyrhynchos) and sandbar sharks (Carcharhinus plumbeus) both primarily consume teleosts (found in 85.2% of gray reef shark stomachs and 70.5% of sandbar shark stomachs) and cephalopods (found in 29.5% of gray reef shark stomachs and 26.5% of sandbar shark stomachs) when either is the only shark species present in a location (Papastamatiou et al. 2006). However, at study sites with large numbers of both gray reef sharks and sandbar sharks, dietary overlap between these species was very low, suggesting resource partitioning (Papastamatiou et al. 2006).
Studies focusing on the diet of a single elasmobranch species across their range have shown that several species have geographic plasticity in their diet. It was found that the diet of the bonnethead sharks (Sphyrna tiburo) is approximately 50% crabs and 5% other crustaceans in Tampa Bay, Florida, but approximately 10% crabs and 30% other crustaceans in Florida Bay, despite similar prey availability in both locations (Bethea et al. 2007). Isotopic mixing model results suggest that the diet of Australian sharpnose sharks was split evenly between benthic and pelagic sources in one study site, but was almost entirely composed of benthic sources in another site (Munroe et al. 2015), which was attributed primarily to these dietary generalists taking advantage of distinct prey availability in different study sites. Our results are consistent with these past studies and build upon them by incorporating multiple co-occurring predators.
By demonstrating that some of these shark species can vary their trophic interactions relative to co-occurring species, our results suggest the potential for trophic plasticity in these animals. Trophic plasticity is believed to benefit predators by making them more resilient to changing environmental conditions, such as declines in the availability of certain prey species (Munroe et al. 2014), while also further complicating any analysis of that species’ ecological role (since that role is variable). A change in dietary preference can also occur as a result of increased competition for resources due to the presence of another predator. For example, African wild dogs (Lycaon pictus) have similar diets as hyenas (Crocuta crocuta), but significantly alter their diet when hyenas are present (Creel and Creel 1996, 1998), resulting in resource partitioning and associated reduced competition for available prey. This shift from possible competition for resources to resource partitioning is similar to what was observed for some species pairs between habitats, possibly as a result of different prey availability between habitats. When voles declined in population, a predatory bird species (the pallid harrier Circus macrourus) capable of trophic plasticity was able to adapt to consume 40% more alternative prey biomass than a co-occurring species that specialized in feeding on voles (the Montagu’s harrier C. pygargus, Terraube et al. 2011). Following a coral mortality event, a species of corallivorous butterflyfish that specialized in consuming one species of coral (Chaetodon trifascialis) suffered 98% population declines, while a co-occurring butterflyfish species capable of changing its diet (C. trifasciatus) suffered still problematic but comparatively lesser 73% population declines (Graham 2007). Co-occurring elasmobranch, teleost and mammal top predators within a single study site have been shown to vary their degree of resource partitioning seasonally (Matich et al. 2017). The shifts in patterns of isotopic niche overlap and relative isotopic signature between sympatric shark species in different habitats may demonstrate trophic plasticity not only in diet, but in competitive interactions for prey.
Patterns of relative isotopic niche area and core isotopic niche overlap
Bull, sandbar and hammerhead sharks had among the highest mean δ15N values in each study area where they were sampled. This suggests that these species may regularly occupy higher trophic positions than the other sampled species in these systems, consistent with a multi-species analysis of shark trophic positions focusing on stomach content analysis results (Cortés 1999). Atlantic sharpnose sharks had among the lowest mean δ15N of sampled species, suggesting that they may feed lower in the food web than the other sampled species from this study. This is consistent with stomach content studies of Atlantic sharpnose diet, which show that this species primarily consumes small teleost fishes (ranging from 93.2% IRI to 100% IRI, Drymon et al. 2012). On the other hand, tiger sharks, which are large animals often considered to be apex predators, had relatively low mean δ15N values in each study area where they were sampled. The generalist diet of tiger sharks includes large but low trophic-level prey like herbivorous turtles (Lowe et al. 1996). This may be responsible for the relatively low mean δ15N values of this species, a pattern also found in Dicken et al. (2017). This finding furthers the assertion (Frisch et al. 2016) that the trophic level of a shark should not be inferred just from a relatively large size.
In this study, bull, sandbar and nurse sharks consistently had large isotopic niche area compared with co-occurring sampled species. Bull sharks had the highest standard ellipse area of any sampled shark species in two of the three habitats included in this study. Similarly, it was found that bull sharks had a much larger isotopic niche area co-occurring predatory than teleost fishes (including wahoo Acanthocybium solandri, dorado Coryphaena hippurus, sailfish Istiophorus platypterus and kawakawa Euthynnus affinis) (Daly et al. 2013). Shiffman et al. (2014) also noted a comparatively large isotopic niche area for sandbar sharks in South Carolina (true here for sandbar sharks in Biscayne Bay but not in the Reef study area), and another study noted a relatively large standard ellipse area for nurse sharks in Belize (Tilley et al. 2013).
Several combinations of shark species studied here that had high core isotopic niche overlap in one location had low or no core isotopic niche overlap in other locations, suggesting that whether two sympatric species partition or compete for available prey can vary dramatically even across a short geographic distance if the habitats are different enough. This variability in resource partitioning between co-occurring predators between locations has been noted in some freshwater systems. For example, it was found that the overlap between white (Morone americana) and yellow perch (Perca flavescens) ellipses was 0% in one study site and 88% in another (Guzzo et al. 2013).
These observed changes in patterns of resource partitioning versus competition for prey may be explained by available prey resources, which varied between the geographically proximate study sites. When a variety of prey resources suitable for both species is available, the predators may partition resources, but when fewer prey types are available, competition may occur. In this study, blacknose and blacktip sharks had 85% core isotopic niche overlap in Biscayne Bay, but 0% overlap in Florida Bay, suggesting a possible shift from consuming similar resources to resource partitioning. Stomach content analysis of these species has revealed that blacktip sharks in the Northwestern Atlantic primarily eat filter-feeding menhaden (Brevoortia tyrannus, Castro 2000), while blacknose sharks eat primarily small piscivorous sciaenids (Ford 2012). However, some teleost, mollusk and crustacean species are consumed by both shark species, and if these prey species are the primary resources available in a location, it may lead to these shark species competing for prey when they would otherwise partition available resources. Similarly, lemon and nurse sharks in this study exhibited high core isotopic niche overlap in one study area, but no overlap in another. Stomach content analysis has shown that both lemon sharks (Cortés and Gruber 1990; Newman et al. 2010) and nurse sharks (Castro 2000) are primarily piscivorous, but also eat benthic invertebrates. When both small invertebrates and fishes are abundant, lemon and nurse sharks may partition available resources between them, but competition may result if a more limited selection of prey is available.
The total isotopic range of the shark community from the Reef study area was highest, likely because of high diversity of prey species in that habitat resulting in complex trophodynamics. With the exception of the lone highly migratory great hammerhead shark in Florida Bay, the total isotopic range of the shark community in Florida Bay was the narrowest, possibly reflecting the comparatively low prey biodiversity in this habitat. These results largely fit into past trophic analyses of these habitats that used stable isotope analysis. The isotopic range of sharks sampled in this study in Biscayne Bay is notably narrower than that reported for the entire food web by Kieckbusch et al. (2004), but comparable to what that study found for the teleost predator community in Biscayne Bay. The relatively enriched δ13C values of sharks in this study further the findings of Kieckbusch et al. (2004) that mangroves are unlikely to be a major carbon source of predators in this habitat. The average isotopic signature of nurse sharks in Florida Bay was very similar to those reported for mesopredatory teleosts in Chasar et al. (2005), with the average N15 signature of the apex predatory bull shark in this study being higher than anything found in Chasar et al. (2005). The lemon sharks analyzed in this study, however, had a slightly lower δ15N signature than those analyzed in Chasar et al. (2005), but those were from a museum collection originally sampled in the 1950s. At the Reef study area sharks appear to be feeding in a food web supported more by algae and coral than the more enriched sea grass-supported food web, and this study’s nurse sharks, a known predator of spiny lobster, have an average isotopic signature approximately one trophic level higher than what Behringer and Butler (2006) found for local spiny lobster.
Model selection
While it is still important to test for the effects of seasonal variation on the isotopic signatures of elasmobranch predators, those effects were expected to be comparatively small. This is because the effects of temporal variation on baseline isotopic signatures on upper-level consumers are comparatively low (Harvey et al. 2002), and few past studies have documented significant seasonal or interannual variation in elasmobranch isotopic signatures (but see Matich and Heithaus 2014 for an analysis of seasonal prey pulses in a similar system). Additionally, Florida waters have less dramatic seasonal changes in environmental conditions than those of Alaska (Goering et al. 1990) or the Mediterranean (Vizzini and Mazzola 2003), and therefore, a less dramatic seasonal change in baseline isotopic signature would be expected for this study than in those previous studies. Additionally, while it is important to test for the effects of size-based variation in the isotopic signature of consumers, this study was not expected to detect ontogenetic shifts in diet or length-based variation in isotopic routing. This is because most of the individuals captured in this study are at or near adult sizes.
Assumptions and limitations
Interpretation of stable isotope analysis results requires making several assumptions that could affect our conclusions if not met (Hussey et al. 2012; Layman et al. 2012). It is possible that two species can have distinct diets but very similar isotopic signatures (for example, if one consumer species’ isotopic signature represents the average of two isotopically extreme resource bases), which would influence our interpretation of overlapping ellipses between species (reviewed in Layman et al. 2012).
Similarly, we are assuming that sharks are consuming resources primarily in the food web associated with the study area where they were captured, but the distances between study sites are below known migration distances of many species included in this study. These annual migrations, when combined with the long isotopic turnover time of elasmobranch blood (see MacNeil et al. 2006; Matich and Heithaus 2014), may mean that few of these sharks ever came into total isotopic equilibrium with the food web where they were captured. This likely means that the isotopic signatures of many or all sharks in this study represent a mix of feeding in their current capture location and elsewhere, an important consideration when interpreting results. However, the fact that we noted such distinct differences in isotopic niche area and overlap between the same species in different study areas suggests that this is not a major concern; if all sampled sharks were all feeding on a shared pool of resources regardless of capture location, these different patterns would not have been observed.
The relatively low sample size of some species included in this study may have underestimated the true isotopic niche variability of these species (e.g., an N of 25–40 captures a population’s isotopic variability well, Vaudo and Heithaus 2011), and all sample sizes were below 25 except for Biscayne Bay nurse sharks, Florida Bay lemon sharks and Reef sandbar sharks (though several other species were close to an N of 25). Similarly, the low sample size of some species in some habitats may have contributed to observed patterns in SEAc overlap—while SEAc is better suited than total occupied niche area for comparing isotopic niche width between small populations (Syväranta et al. 2013), it is certainly possible that a higher sample size would have resulted in more consistent patterns of isotopic niche overlap between study sites as a result of capturing more of the total range in resource use among populations (i.e., our small sample size may have captured outliers in both populations).
While several species of sharks have been known to exhibit ontogenetic shifts in diet (reviewed in Grubbs 2010), this study focused mostly on large juvenile or adult individuals who had likely already undergone their dietary shift. [In many but not all species this shift occurs between small and large juveniles during a transition out of a nursery area, not between large juveniles and adults (Grubbs 2010)]. However, some species do shift habitats and associated diets later in their life history which could cause issues in our interpretation of the data, though our statistical analysis shows that it was likely not a major factor.
In this study, we used only a single tissue type for stable isotope analysis, and this tissue has a relatively long turnover time. Comparing isotopic signatures between tissue types with different turnover rates can reveal whether or not observed patterns in isotopic niche overlap are temporally consistent (e.g., Gallagher et al. 2017, which found that for one of the study sites included in this study, patterns of overlap between species are consistent temporally by comparing isotopic signatures from two distinct tissue types). Also, most of the study species used here have never had their isotopic discrimination factors explicitly measured in laboratory conditions (few elasmobranchs have to date), and different discrimination between species may cause bias in our results (i.e., showing different isotopic signatures even for species with very similar diets because these species may have distinct patterns of isotopic incorporation from their diets).
Not removing urea from samples may result in artificially depleted δ15N values due to nitrogen (Hussey et al. 2012; Carlisle et al. 2016), but this is expected to be more of an issue when comparing elasmobranch isotopic signatures to co-occurring teleosts and invertebrates, and less of an issue when only comparing results between elasmobranchs (Kim and Koch 2012). While this study showed that there were intraspecific differences between animals in different habitats, those habitats were too varied (in terms of bottom type, depth and prey availability) to reliably assess a single cause for observed differences in isotopic niche area between study areas.
Conclusions
This study provides evidence of intraspecific variability in the relative isotopic niche area and patterns of core isotopic niche overlap in several species of co-occurring marine predators, showing that within different habitats the resource usage and trophic interactions of sympatric species may vary. Overall, this suggests that the ecological roles of shark species are quite complex and varied. Therefore, results from a dietary overlap study in one habitat cannot necessarily be extrapolated to a separate habitat, even if that separate habitat is nearby. Our results suggest the need to generate empirical assessments of resource use and trophic interactions among sympatric predators specific to a particular area when trying to understand and model trophodynamics for that area.
References
Anderson WT, Fourqurean JW (2003) Intra-and interannual variability in seagrass carbon and nitrogen stable isotopes from south Florida, a preliminary study. Org Geochem 34(2):185–194
Ault TR, Johnson CR (1998) Spatially and temporally predictable fish communities on coral reefs. Ecol Monogr 68(1):25–50
Behringer DC, Butler MJ (2006) Stable isotope analysis of production and trophic relationships in a tropical marine hard-bottom community. Oecologia 148(2):334–341
Bethea DM, Buckel JA, Carlson JK (2004) Foraging ecology of the early life stages of four sympatric shark species. Mar Ecol Prog Ser 268:245–264
Bethea DM, Hale L, Carlson JK, Cortés E, Manire CA, Gelsleichter J (2007) Geographic and ontogenetic variation in the diet and daily ration of the bonnethead shark, Sphyrna tiburo, from the eastern Gulf of Mexico. Mar Biol 152(5):1009–1020
Carlisle AB, Litvin SY, Madigan DJ, Lyons K, Bigman JS, Ibarra M, Bizzarro J (2016) Interactive effects of urea and lipid content confound stable isotope analysis in elasmobranch fishes. Can J Fish Aquat Sci 74(3):419–428
Castro JI (2000) The biology of the nurse shark, Ginglymostoma cirratum, off the Florida east coast and the Bahama Islands. Environ Biol Fishes 58(1):1–22
Chasar LC, Chanton JP, Koenig CC, Coleman F (2005) Evaluating the effect of environmental disturbance on the trophic structure of Florid Bay using multiple stable isotope analyses of contemporary and historical specimens. Limnol Ocean 50(4):1059–1072
Cortés E (1999) Standardized diet compositions and trophic levels of sharks. ICES J Mar Sci 56(5):707–717
Cortés E, Gruber SH (1990) Diet, feeding habits and estimates of daily ration of young lemon sharks, Negaprion brevirostris (Poey). Copeia 1990:204–218
Creel S, Creel NM (1996) Limitation of African wild dogs by competition with larger carnivores. Conserv Biol 10(2):526–538
Creel S, Creel NM (1998) Six ecological factors that may limit African wild dogs, Lycaon pictus. Anim Conserv 1:1–9
Daly R, Froneman PW, Smale MJ (2013) Comparative feeding ecology of bull sharks (Carcharhinus leucas) in the coastal waters of the southwest Indian Ocean inferred from stable isotope analysis. PLoS ONE 8(10):e78229
Davenport SR, Bax NJ (2002) A trophic study of a marine ecosystem off southeastern Australia using stable isotopes of carbon and nitrogen. Can J Fish Aquat Sci 59(3):514–530
Dicken ML, Hussey NE, Christiansen HM, Smale MJ, Nkabi N, Cliff G, Winter SP (2017) Diet and trophic ecology of the tiger shark from South African waters. PLoS ONE 12(6):e0177897
Drymon JM, Powers SP, Carmichael RH (2012) Trophic plasticity in the Atlantic sharpnose shark (Rhizoprionodon terraenovae) from the north central Gulf of Mexico. Environ Biol Fishes 95(1):21–35
Dulvy NK, Fowler SL, Musick JA, Cavanaugh RD, Kyne PM, Harrison LR, Carlson JK, Davidson LNK, Fordham SV, Francis MP, Pollock CM, Simpfendorfer CA, Burgess GH, Carpenter KE, Compagno LJV, Ebert DA, Gibson C, Heupel MR, Livingstone SR, Sanciangco JC, Stevens JD, Valenti S, White WT (2014) Extinction risk and conservation of the world’s sharks and rays. eLife 3:ee590
Ford RM (2012) Diet and Reproductive Biology of the Blacknose Shark (Carcharhinus acronotus) from the Southwestern Atlantic Ocean. Thesis, University of North Florida
Fourqurean JW, Robblee MB (1999) Florida Bay: a history of recent ecological changes. Estuaries 22(2):345–357
Fourqurean JW, Schrlau JE (2003) Changes in nutrient content and stable isotope ratios of C and N during decomposition of seagrasses and mangrove leaves along a nutrient availability gradient in Florida Bay, USA. Chem Ecol 19(5):373–390
Frisch AJ, Ireland M, Rizzari JR, Lönnstedt OM, Magnenat KA, Mirbach CE, Hobbs JPA (2016) Reassessing the trophic role of reef sharks as apex predators on coral reefs. Coral Reefs 35:1–14
Gallagher AJ, Kyne PM, Hammerschlag N (2012) Ecological risk assessment and its application to elasmobranch conservation and management. J Fish Biol 85(5):1727–1748
Gallagher AJ, Serafy JE, Cooke SJ, Hammerschlag N (2014) Physiological stress response, reflex impairment, and survival of five sympatric shark species following experimental capture and release. Mar Ecol Prog Ser 496:207–218
Gallagher AJ, Shiffman DS, Byrnes EE, Hammerschlag-Peyer CM, Hammerschlag N (2017) Patterns of resource use and isotopic niche overlap among three species of sharks occurring within a protected subtropical estuary. Aquat Ecol 51(3):435–448
Goering J, Alexander V, Haubenstock N (1990) Seasonal variability of stable carbon and nitrogen isotope ratios of organisms in a North Pacific bay. Estuar Coast Shelf Sci 30(3):239–260
Graham NA (2007) Ecological versatility and the decline of coral feeding fishes following climate driven coral mortality. Mar Biol 153(2):119–127
Graham F, Rynne P, Estevanez M, Luo J, Ault JS, Hammerschlag N (2016) Use of marine protected areas and exclusive economic zones in the subtropical western North Atlantic Ocean by large highly mobile sharks. Divers Distrib 22(5):534–546
Grubbs RD (2010) Ontogenetic shifts in movements and habitat use. Sharks and their relatives II: Biodiversity, adaptive physiology, and conservation. CRC Press, Boca Raton, pp 319–350
Guzzo MM, Haffner GD, Legler ND, Rush SA, Fisk AT (2013) Fifty years later: trophic ecology and niche overlap of a native and non-indigenous fish species in the western basin of Lake Erie. Biol Invasions 15(8):1695–1711
Harvey CJ, Hanson PC, Essington TE, Brown PB, Kitchell JF (2002) Using bioenergetics models to predict stable isotope ratios in fishes. Can J Fish Aquat Sci 59(1):115–124
Hayden B, Holopainen T, Amundsen PA, Eloranta AP, Knudsen R, Praebel K, Kahilainen K (2013) Interactions between invading benthivorous fish and native whitefish in subarctic lakes. Freshw Biol 58(6):1234–1250
Heithaus MR, Burkholder D, Hueter R, Heithaus L, Pratt W, Carrier JC (2007) Spatial and temporal variation in shark communities of the lower Florida Keys and evidence for historical population declines. Can J Fish Aquat Sci 64:1302–1313
Heithaus MR, Vaudo JJ, Kreicker S, Layman CA, Krützen M, Burkholder DA, Gastrich K, Bessey C, Sarabia R, Cameron K, Wirsing A, Thomson JA, Dunphy-Daly MM (2013) Apparent resource partitioning and trophic structure of large-bodied marine predators in a relatively pristine seagrass ecosystem. Mar Ecol Prog Ser 481:225–237
Hussey NE, MacNeil MA, Olin JA, McMeans BC, Kinney MJ, Chapman DD, Fisk AT (2012) Stable isotopes and elasmobranchs: tissue types, methods, applications and assumptions. J Fish Biol 80(5):1449–1484
Hussey NE, MacNeil MA, Siple MC, Popp BN, Dudley SF, Fisk AT (2015) Expanded trophic complexity among large sharks. Food Webs 4:1–7
Jackson AL, Inger R, Parnell AC, Bearhop S (2011) Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J Anim Ecol 80(3):595–602
Kieckbusch DK, Koch MS, Serafy JE, Anderson WT (2004) Trophic linkages among primary producers and consumers in fringing mangroves of subtropical lagoons. Bull Mar Sci 74(2):271–285
Kim SL, Koch PL (2012) Methods to collect, preserve, and prepare elasmobranch tissues for stable isotope analysis. Environ Biol Fish 95(1):53–63
Layman CA, Arrington DA, Montaña CG, Post DM (2007) Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology 88(1):42–48
Layman CA, Araujo MS, Boucek R, Hammerschlag-Peyer CM, Harrison E, Jud ZR, Matich PM, Rosenblatt AE, Vaudo JJ, Yeager LA, Post DM, Bearhop S (2012) Applying stable isotopes to examine food-web structure: an overview of analytical tools. Biol Rev 87(3):545–562
Lirman D, Cropper WP (2003) The influence of salinity on seagrass growth, survivorship, and distribution within Biscayne Bay, Florida: field, experimental, and modeling studies. Estuaries 26(1):131–141
Lowe CG, Wetherbee BM, Crow GL, Tester AL (1996) Ontogenetic dietary shifts and feeding behavior of the tiger shark, Galeocerdo cuvier, Hawaiian waters. Environ Biol Fishes 47(2):203–211
MacNeil MA, Drouillard KG, Fisk AT (2006) Variable uptake and elimination of stable nitrogen isotopes between tissues in fish. Can J Fish Aquat Sci 63(2):345–353
Martínez del Rio C, Wolf N, Carleton SA, Gannes L (2009) Isotopic ecology ten years after a call for more laboratory experiments. Biol Rev 84(1):91–111
Matich P, Heithaus MR (2014) Multi-tissue stable isotope analysis and acoustic telemetry reveal seasonal variability in the trophic interactions of juvenile bull sharks in a coastal estuary. J Anim Ecol 83(1):199–213
Matich P, Ault JS, Boucek RE, Bryan DR, Gastrich KR, Harvey CL, Heithaus M, Rosenblatt A (2017) Ecological niche partitioning within a large predator guild in a nutrient-limited estuary. Limnol Ocean 62(3):934–953
Minagawa M, Wada E (1984) Stepwise enrichment of 15N along food chains: further evidence and the relation between δ15N and animal age. Geochim Cosmochim Acta 48(5):1135–1140
Munroe SEM, Simpfendorfer CA, Heupel MR (2014) Defining shark ecological specialisation: concepts, context, and examples. Rev Fish Biol Fish 24(1):317–331
Munroe SEM, Heupel MR, Fisk A, Logan M, Simpfendorfer CA (2015) Regional movement patterns of a small-bodied shark revealed by stable-isotope analysis. J Fish Biol 86(5):1567–1586
Newman SP, Handy RD, Gruber SH (2010) Diet and prey preference of juvenile lemon sharks Negaprion brevirostris. Mar Ecol Prog Ser 398:221–234
Papastamatiou YP, Wetherbee BM, Lowe CG, Crow GL (2006) Distribution and diet of four species of carcharhinid shark in the Hawaiian Islands: evidence for resource partitioning and competitive exclusion. Mar Ecol Prog Ser 320:239–251
Peterson BJ, Fry B (1987) Stable isotopes in ecosystem studies. Annu Rev Ecol Syst 18:293–320
Pikitch E, Santora C, Babcock EA, Bakun A, Bonfil R, Conover DO, Dayton P, Doukakis P, Fluharty D, Heneman B, Houde ED, Link J, Livingston PA, Mangel M, McAllister M, Pope J, Sainsbury KJ (2004) Ecosystem-based fishery management. Science 305(5682):346–347
Polis GA, Holt RD (1992) Intraguild predation: the dynamics of complex trophic interactions. Trends Ecol Evol 7(5):151–154
Post DM (2002) Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology 83(3):703–718
R Core Team (2013) R: a language and environment for statistical computing. R Foundation for Statistical Computing. Vienna, Austria. http://www.R-project.org/
Sanford E, Roth MS, Johns GC, Wares JP, Somero GN (2003) Local selection and latitudinal variation in a marine predator-prey interaction. Science 300(5622):1135–1137
Schoener TW (1974) Resource partitioning in ecological communities. Science 185(4145):27–39
Serafy JE, Faunce CH, Lorenz JJ (2003) Mangrove shoreline fishes of Biscayne Bay, Florida. Bull Mar Sci 72(1):161–180
Shiffman DS, Gallagher AJ, Boyle MD, Hammerschlag-Peyer CM, Hammerschlag N (2012) Stable isotope analysis as a tool for elasmobranch conservation research: a primer for non-specialists. Mar Fresh Res 63(7):635–643
Shiffman DS, Frazier BS, Kucklick JR, Abel D, Brandes J, Sancho G (2014) Feeding ecology of the sandbar shark in South Carolina estuaries revealed through δ13C and δ15 N stable isotope analysis. Mar Coast Fish 6(1):156–169
Simpfendorfer CA, Heupel MR, White W, Dulvy NK (2011) The importance of research and public opinion to conservation management of sharks and rays: a synthesis. Mar Fresh Res 62:518–527
Swart PK, Saier A, Lamb K (2005) Temporal and spatial variation in the δ15N and δ13C of coral tissue and zooxanthellae in Montastraea faveolata collected from the Florida reef tract. Limnol Ocean 50(4):1049–1058
Syväranta J, Lensu A, Marjomäki TJ, Oksanen S, Jones RI (2013) An empirical evaluation of the utility of convex hull and standard ellipse areas for assessing population niche widths from stable isotope data. PLoS ONE 8(2):e56094
Tamburello N, Côté IM, Dulvy NK (2015) Energy and the scaling of animal space use. Am Nat 186(2):196–211
Terraube J, Arroyo B, Madders M, Mougeot F (2011) Diet specialisation and foraging efficiency underfluctuating vole abundance: a comparison between generalist and specialist avian predators. Oikos 120(2):234–244
Thayer GW, Powell AB, Hoss DE (1999) Composition of larval, juvenile, and small adult fishes relative to changes in environmental conditions in Florida Bay. Estuaries 22(2):518–533
Tilley A, López-Angarita J, Turner JR (2013) Diet reconstruction and resource partitioning of a Caribbean marine mesopredator using stable isotope Bayesian modelling. PLoS ONE 8(11):e79560
Torres LG, Heithaus MR, Delius B (2006) Influence of teleost abundance on the distribution and abundance of sharks in Florida Bay, USA. Hydrobiologia 569:449–455
Trent L, Parshley DE, Carlson JK (1997) Catch and bycatch in the shark drift gillnet fishery off Georgia and east Florida. Mar Fish Rev 59(1):19–28
Vaslet A, Phillips DL, France C, Feller IC, Baldwin CC (2012) The relative importance of mangroves and seagrass beds as feeding areas for resident and transient fishes among different mangrove habitats in Florida and Belize: evidence from dietary and stable-isotope analyses. J Exp Mar Biol Ecol 434:81–93
Vaudo JJ, Heithaus MR (2011) Dietary niche overlap in a nearshore elasmobranch mesopredator community. Mar Ecol Prog Ser 425:247–260
Vizzini S, Mazzola A (2003) Seasonal variations in the stable carbon and nitrogen isotope ratios (13C/12C and 15 N/14 N) of primary producers and consumers in a western Mediterranean coasal lagoon. Mar Biol 142(5):1009–1018
Wieters EA, Gaines SD, Navarrete SA, Blanchette CA, Menge BA (2008) Scales of dispersal and the biogeography of marine predator–prey interactions. Am Nat 171(3):405–417
Acknowledgments
The authors would like to thank the other members of author D.S.’s dissertation committee: Dr. Nicholas K Dulvy, Dr. Andrew R Binder and Gina Maranto. We would like to thank Robert Michener, the manager of author L. K.’s stable isotope laboratory, for his assistance with sample preparation and analysis and his detailed description of laboratory methods. We would like to thank Dr. Catherine Macdonald, Christian Pankow, Rachel Skubel, Emily Nelson and Jacob Jerome for their assistance with sample management and retrieval, and Lindsay Jennings for her assistance with the UM Shark Research and Conservation Program’s sample database. Sample processing fees were provided by more than 100 unique crowdfunding donors through the SciFund Challenge. The American Elasmobranch Society provided a student travel grant for author D.S. to present this research at the 2016 AES conference, and we thank the scientists who attended that presentation for their helpful feedback. Figure 1 was created by author NH’s masters student Abby Tinari, and we thank her for her assistance. This research was carried out under the following permits: University of Miami Institutional Animal Care and Use Committee 15-238, Florida Fish and Wildlife Conservation Commission SAL 15-0957-SR, Florida Keys National Marine Sanctuary 2015-168, National Oceanographic and Atmospheric Administration Highly Migratory Species SHK-EFP-16-02, Biscayne National Park 2015-SCI-0012 and Everglades National Park 2015-SCI-0011. The authors have no conflict of interest to declare.
Author information
Authors and Affiliations
Contributions
This work represents Ph.D. dissertation research by author D.S., who performed data analysis and writing. Authors M.H. and N.H. served on D.S.’s dissertation committee and provided guidance on the planning of this project as well as provided guidance on analysis and write-up. Author L.K. assisted with data analysis and write-up.
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Handling Editor: Télesphore Sime-Ngando.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Shiffman, D.S., Kaufman, L., Heithaus, M. et al. Intraspecific differences in relative isotopic niche area and overlap of co-occurring sharks. Aquat Ecol 53, 233–250 (2019). https://doi.org/10.1007/s10452-019-09685-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10452-019-09685-5