
Abstract
To examine variation in diet and daily ration of the bonnethead shark, Sphyrna tiburo (Linnaeus 1758), animals were collected from three areas in the eastern Gulf of Mexico: northwest Florida (∼29°40′N, 85°13′W), Tampa Bay near Anclote Key (∼28°10′N, 82°42.5′W), and Florida Bay (∼24°50′N, 80°48′W) from March through September, 1998–2000. In each area, diet was assessed by life stage (young-of-the year, juveniles, and adults) and quantified using five indices: percent by number (%N), percent by weight (%W), frequency of occurrence (%O), index of relative importance expressed on a percent basis (%IRI), and %IRI based on diet category (%IRIDC). Diet could not be assessed for young-of-the-year in Tampa Bay or Florida Bay owing to low sample size. Diet analysis showed an ontogenetic shift in northwest Florida. Young-of-the-year stomachs from northwest Florida (n = 68, 1 empty) contained a mix of seagrass and crustaceans while juvenile stomachs (n = 82, 0 empty) contained a mix of crabs and seagrass and adult stomachs (n = 39, 1 empty) contained almost exclusively crabs. Crabs made up the majority of both juvenile and adult diet in Tampa Bay (n = 79, 2 empty, and n = 88, 1 empty, respectively). Juvenile stomachs from Florida Bay (n = 72, 0 empty) contained seagrass and a mix of crustaceans while adult stomachs contained more shrimp and cephalopods (n = 82, 3 empty). Diets in northwest Florida and Tampa Bay were similar. The diet in Florida Bay was different from those in the other two areas, consisting of fewer crabs and more cephalopods and lobsters. Plant material was found in large quantities in all stomachs examined from all locations (>15 %IRIDC in 6 of the 7 life stage-area combinations, >30 %IRIDC in 4 of the 7 combinations, and 62 %IRIDC in young-of-the-year diet in northwest Florida). Using species- and area-specific inputs, a bioenergetic model was constructed to estimate daily ration. Models were constructed under two scenarios: assuming plant material was and was not part of the diet. Overall, daily ration was significantly different by sex, life stage, and region. The bioenergetic model predicted increasing daily ration with decreasing latitude and decreasing daily ration with ontogeny regardless of the inclusion or exclusion of plant material. These results provide evidence that bonnetheads continuously exposed to warmer temperatures have elevated metabolism and require additional energy consumption to maintain growth and reproduction.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
It has long been postulated that many species of sharks are top predators and, as such, are hypothesized to play a major role in structuring marine communities through consumption (Cortés 1999; Stevens et al. 2000). The higher trophic levels occupied by sharks in conjunction with some recent declines in a number of their populations (Musick et al. 2000; Cortés 2004) have increased interest in investigating the impacts that fishery removals of sharks can have on marine ecosystems. Despite calls for an ecosystem approach to fisheries management (NRC 1998; NMFS 1999b), there is still little quantitative information on diet, consumption, and predator-prey interactions pertinent to sharks (NMFS 1999a).
Quantitative diet analysis, and its use in bioenergetic models, has become important to shark population and ecosystem modeling (See e.g., Kitchell et al. 2002; Lowe 2002; Schindler et al. 2002; Carlson 2007; Neer et al. 2007). Although metabolic rate is regarded as the largest and most variable component of the bioenergetic model (Lowe 2001) and sensitivity analyses have demonstrated that metabolism can have the greatest effect on predicted consumption rates (Bartell et al. 1986; Essington 2003), early attempts to develop consumption rates for sharks through the bioenergetic approach relied on borrowing metabolic rates from different species (e.g., Stillwell and Kohler 1982, 1993). Technological advances have enabled progress in estimating species-specific metabolic rates for a variety of sharks (see review in Carlson et al. 2004). Estimates of daily ration using species-specific physiological parameters are now available for juvenile lemon shark, Negaprion brevirostris (Cortés and Gruber 1990), scalloped hammerhead shark, Sphyrna lewini (Lowe 2001) and sandbar shark, Carcharhinus plumbeus (Dowd et al. 2006).
The bonnethead, Sphyrna tiburo, is a relatively small species of shark that is common in coastal areas off the southeastern United States and Gulf of Mexico. In some coastal habitats, bonnetheads are the dominant shark species and potentially occupy top-tier trophic levels (McCandless et al. 2002). Previous studies on bonnetheads showed latitudinal gradients in growth, maturity, and size of near-term embryos in the eastern Gulf of Mexico (Parsons 1993; Carlson and Parsons 1997; Lombardi-Carlson et al. 2003). Parsons (1993) hypothesized that this variation was the result of differences in energy consumption and metabolism along the latitudinal gradient. To test this hypothesis, we examine the feeding ecology and daily ration of bonnetheads in the eastern Gulf of Mexico. Specifically, we (1) describe and quantify the diet and feeding ecology of bonnetheads by life stage and area (following Lombardi-Carlson et al. 2003), and (2) model consumption using a bioenergetic approach for life stages and sexes from each area, examining the sensitivity of the model to input parameters.
Methods
Specimen collection and diet analysis
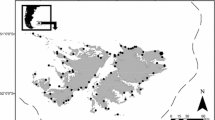
Bonnetheads were collected from fishery-independent surveys in three areas in the eastern US Gulf of Mexico from March through September 1998–2000: Northwest Florida (St Andrew Bay, Crooked Island Sound, St Joseph Bay, and the gulf-side of St Vincent Island; ∼29°40′N, 85°13′W), Tampa Bay near Anclote Key (∼28°10′N, 82°42.5′W), and Florida Bay (∼24°50′N, 80°48′W) (Fig. 1).

Map of sampling areas. Sharks were collected in northwest Florida from St Andrew Bay to St Vincent Island, Tampa Bay near Anclote Key, and Florida Bay, Florida
In northwest Florida, sharks were collected with gillnets of different stretch-mesh sizes (following Carlson and Brusher 1999). In Tampa Bay and Florida Bay, sharks were collected in the same way except using gillnets with a single stretch mesh of 11.4 cm. For each shark sampled, total length (TL, cm) was measured and sex and life stage were determined. Sharks were placed on ice. Upon returning to the laboratory, stomachs were removed, placed in plastic bags, and frozen at −20°C until processing.
Stomachs were thawed for 1 h, opened, and rinsed with water over a 595 μm sieve. Items found in the stomachs were identified to the lowest possible taxon, counted, measured for length (nearest cm), and weighed (wet weight, nearest 0.001 g).
Diets were assessed by life stage in each area: young-of-the-year (YOY) (i.e., age 0+; characterized by having an open or healed, but visible, umbilical scar), juvenile (characterized as not yet being mature) and (3) adult. Adult males were characterized as having calcified claspers and well-developed testes. Adult females were characterized as having developed oocytes or the presence of pups. A preliminary analysis on a subset of stomachs from each of the three areas revealed that diet of males and females was not substantially different, thus we opted not to pursue further diet comparisons by sex.
Diet was quantified using percent by number (%N), percent by weight (%W), and percent frequency of occurrence (%O). Items rarely found in stomachs (e.g., rocks, hooks, coral, and benthos), parasites (e.g., nematode worms), and digested material were not included in stomach content analysis. Plant material was included in stomach content analysis. When calculating %N for plant material, one unit equaled one leaf of seagrass or one mat of algae. The index of relative importance (IRI; Pinkas et al. 1971) was calculated as IRI = %O (%N + %W). The IRI for each item was divided by the total IRI for all items to get the index of relative importance on a percent basis (%IRI; Cortés 1997).
After a full stomach content analysis, identifiable items found in the stomachs were grouped into eight major diet categories (DC) to facilitate comparisons among life stages and sites: (1) crabs, (2) shrimps, (3) lobsters, (4) crustaceans other than crabs, shrimps, and lobsters (including unidentified decapods), (5) cephalopods, (6) non-cephalopod molluscs (including unidentified molluscs), (7) plant material (including shoal grass, Halodule wrightii, turtlegrass, Thalassia testudinum, and other plant material), and (8) teleosts. The index of relative importance on a percent basis was computed for the eight major diet categories (%IRIDC) and used in all analyses.
Cumulative prey curves were constructed a posteriori for each life stage in each area to determine whether an adequate number of stomachs had been collected to accurately describe diets (Ferry and Cailliet 1996). All identifiable, non-excluded diet items were counted as unique. When a cumulative prey curve reaches an asymptote, the number of stomachs analyzed is considered sufficient in describing dietary habits.
Ontogenetic and geographic variations in diet
Two methods were used to test for changes in diet by life stage and area. First, dietary overlap was calculated using Pianka’s overlap index (Ecological Methodology v5.1 software; Krebs 1999). All resources were assumed equally abundant and resource state values were presented as %IRIDC. Overlap index values range from 0 (no overlap) to 1.0 (complete overlap), with values ≥0.6 considered “biologically significant” overlap (Pianka 1976). The observed overlap values were then compared to a distribution of expected values based on null-model simulations. The distribution of null-model data came from 1,000 randomizations of the diet data (R3 randomization algorithm; Winemiller and Pianka 1990). Simulations were performed using EcoSim v7.42 software (Gotelli and Entsminger 2005). The observed value was considered statistically different from the null distribution if it was greater than or less than the simulated index 95% of the time (P < 0.05; Winemiller and Pianka 1990). An observed value significantly lower than the simulation index would suggest competition and diet partitioning. An observed value significantly higher than the simulation index would suggest a lack of competition or strong competition that has not yet led to diet partitioning. Second, correspondence analysis was used to detect trends in diet relative to location and life stage (following Graham and Vrijenhoek 1988). In the contingency tables, life stage-area combinations were entered as rows and diet categories as columns. A Chi-square test was used to verify that the rows and columns were independent.
Prey size-predator size analysis
To estimate changes in prey size with increasing shark size, an absolute prey size-predator size diagram was plotted for each area. All prey sizes used in this analysis were crustacean carapace length (CL, cm) from crustacean prey found whole in the stomachs. Quantile regression techniques (Scharf et al. 2000) were used to determine the mean (50th quantile) and upper and lower bounds (90th and 10th quantiles) of the relation between prey size and shark size. A frequency histogram of relative and cumulative prey size-predator size ratios was also created to examine the patterns of prey size use in each area.
Bioenergetic model
Estimates of consumption were developed following the balanced bioenergetic approach of Winberg (1960) expressed as: C = (M r + M s) + (G S + G R) + (W F + W u), where C = consumption; M r = routine metabolic rate; M s = specific dynamic action due to energetic costs of digestion; G S = energy allocated to somatic growth; G R = energy allocated to reproduction; and W F and W u = the energy lost to the production of feces and urine, respectively. All rates were expressed in kilocalories per day (kcal day−1). Daily ration was calculated as: DR = C/F/W, where C = consumption (kcal day−1); F = energy value of the food source (kcal g−1 wet weight); and W = mass of the shark (g). Daily ration was then expressed as percent body weight per day (C/F/W × 100 = %BW day−1). The mass of an individual was taken from the mean of a distribution of individuals by life stage from monthly field collections from each area throughout the year. Proportions of food were taken from the %IRIDC for each life stage in each area. The caloric values of the non-plant diet categories were taken from studies by Thayer et al. (1973) and Steimle and Terranova (1985). The caloric value for plant material was calculated as the average of caloric values found in Lobel and Ogden (1981) and Tenore (1981) for the most common plant materials found in the diet.
Bioenergetic models were constructed for sex, area, and life stage. Because of the high proportion of plant material in the diet and the uncertainty regarding its energetic contribution, bioenergetic models were constructed both including and excluding plant material. Species-specific information on routine metabolism across a range of body sizes and temperatures was taken from studies by Parsons (1990) and Carlson and Parsons (1999). The relationship of routine metabolism to mass was expressed as: VO2 = 68.9 + 177.8W, where VO2 = oxygen consumption rate (mg O2 h−1) and W = mass of the shark (g). Oxygen consumption rate was converted to calories using the oxycalorific coefficient for fish of 3.25 cal mg O −12 (Brafield and Solomon 1972). Because each area is subjected to differences in annual water temperature, metabolic rates were corrected by assuming a Q 10 of 2.3 (Carlson and Parsons 1999). Similarly, temperature was obtained from yearly averages of field survey data and NOAA oceanographic buoys within each area. Generally, a constant activity multiplier (Winberg 1960) is applied to estimates of standard metabolic rate to express the increase from standard to routine/active metabolic rate in the field (e.g. Kitchell et al. 1977; Hansen et al. 1993). We chose not to apply an activity multiplier in our bioenergetic model. We were confident our estimates of metabolic rate matched field-based estimates based on results from Parsons (1990), Parsons and Carlson (1998), and Carlson and Parsons (2003). Specific dynamic actions were set at 6% for YOY and 12% for adults based on Sims and Davies (1994). Energetic loss due to feces and urine was set at 27% of consumption based on results of Wetherbee and Gruber (1993).
Area-specific estimates of growth were applied based on Lombardi-Carlson et al. (2003). Growth rates (converted to mass) were obtained from von Bertalanffy growth functions. The predicted growth between age classes was used to model growth among life stages and areas. The growth of YOY was estimated as the growth for sharks from birth to age one. Growth for juveniles was determined for those ages up to maturity depending on area, and growth for adults was calculated for those ages determined to be mature to the maximum observed age (Lombardi-Carlson et al. 2003).
To estimate the energetic investment in reproduction, we used yearly determinations of male and female reproductive tissues (mass). For females, the number of embryos produced each year was multiplied by the average mass of near-term embryos. Male reproductive investment was determined by converting the length of each testis to mass (based on Parsons 1993). Mass for both growth and reproduction was converted to kilocalories by using the energy density of shark tissue of 1.294 kcal g−1 (wet weight) based on estimates for lemon shark by Cortés and Gruber (1990).
Most estimates of daily ration for sharks have been based on point estimates despite the variability and uncertainty in input parameters (Wetherbee and Cortés 2004). We used Monte Carlo simulation to assess uncertainty in the model input parameters (Bartell et al. 1986). Probability density functions were developed to describe temperature, life-stage specific mass of an individual throughout the year, mass of embryos, litter size, growth, excretion, specific dynamic action, and diet. Excretion was represented by a triangular distribution with 27% of consumption as the likeliest value using ±10% as lower and upper bounds. Specific dynamic action was a uniform distribution with 6 and 12% of total consumption as the lower and upper bounds. Annual growth rates obtained from the von Bertalanffy growth equation parameters were assigned lognormal distributions with coefficients of variation of 10% (Bartell et al. 1986). Normal distributions were assigned to mass of individuals, embryos, gonads, and environmental temperature. Diet was a custom distribution with the relative probability of diet components represented by the %IRIDC.
The simulation process involved randomly selecting a value from the set of input parameters from the probability density functions and calculating daily ration. This process was repeated 2,000 times, yielding frequency distributions, means, and confidence intervals for parameter estimates (calculated as the 2.5th and 97.5th percentiles). All simulations were run with Microsoft Excel© spreadsheet software equipped with risk analysis software (Crystal Ball© 2000 Academic Edition v5.2.2, Decisioneering Inc.). Sensitivity analysis was performed to examine the implications of the uncertainty and variability of inputs on daily ration. Multiple-factor analysis of variance (ANOVA) was used to test for differences in log-transformed daily ration simulations among and within life stages, sex, and area.
Results
Northwest Florida diet
A total of 191 stomachs were collected from sharks in northwest Florida. Of those, 68 (1 empty) were from YOY, 82 (0 empty) were from juveniles, and 39 (1 empty) were from adults (Appendix 1). Cumulative prey curves showed general trends toward an asymptote (Appendix 2), indicating that enough stomachs were sorted to adequately describe the diet of all life stages in northwest Florida.
For YOY, plant material was the most important of the eight diet categories (62.1 %IRIDC), followed by crustaceans (other than crabs, lobsters, and shrimp; 22.2 %IRIDC) and crabs (13.5 %IRIDC). Shrimp and molluscs contributed very little to the diet (2.1 %IRIDC and 0.1 %IRIDC, respectively). All other categories were not important in the diet (Table 1A).
Crabs were the dominant category in the diet of juveniles (63.1 %IRIDC). Plant material was the second-most important diet category (26.8 %IRIDC). Crustaceans (other than crabs, lobsters, and shrimp; 6.6 %IRIDC) were much less important in the diet. Molluscs (other than cephalopods) were a more important diet category for juveniles (2.8 %IRIDC) than for YOY. Shrimp were of little importance in the diet (0.7 %IRIDC). All other categories were not important (Table 1A).
Adults fed almost entirely on crabs (73.1 %IRIDC). Crustaceans (other than crabs, lobsters, and shrimp) were the second-most important diet category (15.1 %IRIDC). Plant material remained important in the diet of adults (8.8 %IRIDC). Molluscs other than cephalopods also remained important (2.9 %IRIDC). All other categories did not contribute to the diet (Table 1A).
Tampa Bay diet
Stomachs sorted from Tampa Bay numbered 170. Of those, 79 (2 empty) were juveniles and 88 (1 empty) were adults (Appendix 3). Sample size was not large enough to quantitatively describe the diet of YOY sharks in Tampa Bay (n = 3). The cumulative prey curves trended toward an asymptote (Appendix 4), showing that the number of stomachs analyzed was likely sufficient in describing the diet in this area.
Crabs were the most important diet category for both juveniles and adults in Tampa Bay (46.6 %IRIDC and 49.8 %IRIDC, respectively), followed very closely by plant material (42.4 %IRIDC and 45.2 %IRIDC, respectively). For juveniles, crustaceans (other than crabs, lobsters, and shrimp; 7.3 %IRIDC) and cephalopods (3.2 %IRIDC) were important. For adults, crustaceans (other than crabs, lobsters, and shrimp) were important (3.2 %IRIDC); however, cephalopods were less important (0.9 %IRIDC). For both life stages, non-cephalopods molluscs, shrimp, and teleosts were of little importance in the diet. One lobster (Panulirus argus) occurred in the diet of juvenile sharks (Table 1B and Appendix 3).
Florida Bay diet
One hundred and fifty-four stomachs were sorted from Florida Bay. Stomachs taken from juveniles numbered 72 (0 empty) and 82 (3 empty) were from adult sharks (Appendix 5). Similar to Tampa Bay, sample size was too small to quantitatively describe the diet of YOY sharks (n = 1). The asymptotic trends of the cumulative prey curves were not as evident as in the two northernmost areas (Appendix 6), thus more stomachs may need to be collected and sorted to better describe the diet of bonnetheads in Florida Bay.
Juvenile diet was dominated by crustaceans (other than crabs, lobsters, and shrimp; 38.1 %IRIDC) and plant material (30.2 %IRIDC). Crabs (11.8 %IRIDC), lobsters (7.7 %IRIDC), cephalopods (7.0 %IRIDC), and shrimp (4.3 %IRIDC) were also important categories in the diet. Categories of little dietary importance included teleosts and non-cephalopod mollusks (Table 1C).
Adult sharks fed mostly on a mix of crustaceans (other than crabs, lobsters, and shrimp; 29.7 %IRIDC) and cephalopods (29.3 %IRIDC). Plant material was not as important in the diet of adults in Florida Bay; however, it was still the third-most important diet category (15.8 %IRIDC). Crabs, lobsters, shrimp, and teleosts were also important categories in the diet (8.1 %IRIDC, 7.5 %IRIDC, and 5.5 %IRIDC, 4.1 %IRIDC, respectively). Non-cephalopod mollusks remained unimportant in the diet (Table 1C).
Ontogenetic and geographic variations in diet
Pianka’s overlap index was not significant for comparisons between YOY and the other two life stages within northwest Florida (0.57 and 0.37, respectively; Table 2—P o). Diet overlap was high between juveniles and adults in northwest Florida (0.96). Diet overlap was high (>0.78) between juveniles and adults in both Tampa Bay and Florida Bay. All life stages in northwest Florida showed biologically significant diet overlap with juveniles and adults in Tampa Bay (all >0.78). Sharks in Florida Bay showed non-significant biological diet overlap with sharks from the more northern sites, with the exception of juveniles in Florida Bay and YOY in northwest Florida (0.84). Null model analysis (simulated overlap values) mirrored the observed biologically significant diet overlap values (Table 2—P o*). All comparisons that were biologically significant (P o) showed lower than expected simulated overlap values when using the null model (P o*).
Axes 1 and 2 in the correspondence analysis accounted for 90.0% of the total variation among diets of life stages in all areas (24.1 and 65.9%, respectively; Fig. 2). For sharks collected in northwest Florida, YOY grouped with plant material and crustaceans other than crabs, lobsters, and shrimp. Juveniles and adults in northwest Florida grouped close to each other and closely with crabs and molluscs other than cephalopods. Both life-stages in Tampa Bay also grouped close to each other as well as crabs, plant material, and molluscs other than cephalopods. Juveniles in Florida Bay grouped close to YOY in northwest Florida and also with shrimp, plant material, and crustaceans other than crabs, lobsters, and shrimp. Adults in Florida Bay grouped close to cephalopods, lobsters, shrimp, and teleosts. The Chi-square test for life stage-area combinations and diet category was significant (χ2 = 434.02; P < 0.0001; df = 42), indicating dependence.

Plot of life stage and major diet category principal components for axis 1 and 2 of a correspondence analysis using %IRIDC data for bonnetheads. Open circles area-life stage combination; X diet category. NWF northwest Florida; TB Tampa Bay; FB Florida Bay. YOY young-of-the-year; JUV juveniles; MAT adults. CRA crab; LOB lobster; SHR shrimp; CRU crustaceans other than crabs, shrimps, lobsters (including unidentified decapods); CEP cephalopods; MOL other molluscs (including unidentified molluscs); PLA plant material; TEL teleosts
Prey size-predator size relationships
Median, maximum, and minimum prey sizes increased with shark size for bonnetheads in northwest Florida and Tampa Bay (Fig. 3a, b). Absolute prey size did not increase significantly with predator size in Florida Bay (P = 0.085; Fig. 3c). The maximum prey size consumed by sharks in Florida Bay was also not significant (Fig. 3c; 90th quantile, P = 0.113); however, the minimum prey size consumed was (10th quantile, P = 0.037). These results are likely due to small sample size in this area (n = 40). Only sharks in northwest Florida continued to include small prey in their diet with increasing size (Fig. 3A; 10th quantile, P = 1.115).

Prey size-predator size scatter diagram for bonnetheads (n = 108) in the eastern Gulf of Mexico from a northwest Florida, b Tampa Bay, and c Florida. Lines quantile regressions used to examine changes in prey size eaten with increasing shark size. Solid line median prey size (50th quantile). Dotted line minimum and maximum prey sizes (10th and 90th quantiles). Each symbol is a single crustacean prey eaten by a shark. CL carapace length (cm), TL shark total length (cm). * Indicates P < 0.05. ** Indicates P < 0.01. *** Indicates P < 0.001. Regression equations are in Appendix 7
Bonnetheads in the eastern Gulf of Mexico consumed prey that were small fractions of their total length; 95.6% of all prey measured were less than 13% of shark TL (Fig. 4). Almost all of the measured prey consumed in northwest Florida was less than 10% of shark TL; however, over one-third was less than 5% of shark TL (Fig. 4A). Sharks in Tampa Bay consumed prey less than 10% of their total length 87.5% of the time while consuming prey less than 5% of their TL only 7.5% of the time (Fig. 4b). Sharks in Florida Bay consumed a wider range of prey sizes; 77.5% of prey consumed was less than 10% shark TL and 25.0% was less than 5% shark TL (Fig. 4c).

Relative frequency distributions of prey size-predator size ratios for bonnetheads in the eastern Gulf of Mexico from a northwest Florida, b Tampa Bay, and c Florida. Bars relative frequencies at 1% intervals of prey size-predator size ratios. Filled circles cumulative frequencies at 1% intervals
Bioenergetic model
Regardless of the inclusion of plant material, the bioenergetic model predicted increasing daily ration estimates with decreasing latitude and decreasing daily ration estimates with ontogeny (Table 3A-B). In three out of seven comparisons, females had higher daily ration estimates than males. Overall, YOY sharks in northwest Florida had the highest estimates of daily ration (5.33 %BW day−1 for males and 5.46 %BW day−1 for females, including plant material; 4.08 %BW day−1 for males and 4.38 %BW day−1 for females, excluding plant material). For juveniles, sharks in northwest Florida had the lowest estimates of daily ration (1.34 %BW day−1 for males and 1.41 %BW day−1 for females, including plant material; 1.29 %BW day−1 for males and 1.33 %BW day−1 for females, excluding plant material), those in Florida Bay had the highest (2.71 %BW day−1 for males and 1.68 %BW day−1 for females, including plant material; 2.36 %BW day−1 for males and 1.48 %BW day−1 for females, excluding plant material), and Tampa Bay juveniles had intermediate values (1.44 %BW day−1 for males and 1.72 %BW day−1 for females, including plant material; 1.22 %BW day−1 for males and 1.45 %BW day−1 for females, excluding plant material). The trend for adult sharks was the same as for juveniles. Adults in northwest Florida had the lowest estimates (0.55 %BW day−1 for males and 0.37 %BW day−1 for females, both including and excluding plant material), those in Florida Bay had the highest estimates (1.18 %BW day−1 for males and 0.75 %BW day−1 for females, including plant material; 1.11 %BW day−1 for males and 0.71 %BW day−1 for females, excluding plant material), while those in Tampa Bay were in between (0.73 %BW day−1 for males and 0.60 %BW day−1 for females, including plant material; 0.63 %BW day−1 for males and 0.51 %BW day−1 for females, excluding plant material). Regardless of the inclusion of plant material, there were significant differences in daily ration for life stages and sex between areas (3-factor ANOVAs comparing juveniles to adults only, all P < 0.001) and life stage and sex within areas (2-factor ANOVAs, all P < 0.0001).
Monte Carlo simulations resulted in wide confidence intervals around mean estimates of daily ration by sex, size, and area (Table 3). Sensitivity analysis revealed that, of the input parameters, uncertainty in mass of an individual and temperature contributed the most to variation in the estimate of daily ration (−0.83 and 0.28%, respectively). Sensitivity analysis also indicated that the diet of YOY bonnetheads from northwest Florida contributed to the uncertainty in daily ration estimates (−0.31%). Mass of an individual and temperature contributed primarily to the estimate of metabolism. The caloric values of the diet categories also influenced the final estimate of daily ration.
Discussion
Crustaceans were common in diets of all life stages in all areas. Sharks in the two northernmost areas consumed mostly crabs while sharks in Florida Bay ate mostly cephalopods and crustaceans other than crab and shrimp. The importance of lobsters and cephalopods in the diet of sharks in Florida Bay was the most interesting difference in the diets among areas. Cortés et al. (1996) found that bonnethead diet consisted almost entirely of crustaceans in Tampa Bay and Charlotte Harbor. Parsons (1987) found cephalopods to be the most important item in the diets of bonnetheads in Florida Bay.
Ontogenetic diet shifts are common in sharks (see Wetherbee and Cortés 2004, and references therein); however, Cortés et al. (1996) concluded that bonnetheads in Tampa Bay and Charlotte Harbor, Florida, underwent dietary shifts depending on season and habitat but not ontogeny. Seasonal differences in diet were not examined in this study. Shifts in diet with ontogeny occurred only in northwest Florida. Sharks in Tampa Bay and Florida Bay did not show an ontogenetic shift in diet; however we did not examine YOY stomach contents from these areas. Diet of YOY in northwest Florida was composed almost entirely of crustaceans, whereas both juveniles and adults fed almost entirely on crabs. Plant material was abundant in stomach contents in all areas and all life stages.
Geographic differences in diet are documented for many shark species (e.g., lemon shark, N. brevirostris, Cortés and Gruber 1990; sandbar shark, C. plumbeus, Ellis 2003; starspotted smoothhound, Mustelus manazo, Yamaguchi and Taniuchi 2000; Atlantic sharpnose shark, Rhizoprionodon terraenovae, Bethea et al. 2006). The differences in diet with latitude found in the present study are most likely due to prey availability and habitat differences associated with the latitudinal gradient between temperate/subtropical and subtropical/tropical geographic regions. For example, prey species usually associated with temperate/subtropical seagrass beds, sand flats, and muddy substrates (e.g., portunid crabs) were found more often in the diets of sharks from northwest Florida and Tampa Bay, whereas invertebrates usually associated with hard bottom in subtropical/tropical regions (e.g. Florida spiny lobster, P. argus, and Octopus sp.) were more common in the diets of sharks from Florida Bay.
Bonnethead diet consists of relatively small prey. Relative frequency distributions of prey size-predator size ratios for bonnetheads in the eastern Gulf of Mexico are comparable to those of elasmobranch predators in the northwestern Atlantic whose diets also consist of mostly small, invertebrate prey (less than 20% of fish length; e.g., smooth dogfish, Mustelus canis, spiny dogfish, Squalus acanthias, winter skate, Raja ocellata, and little skate, R. erinacea, in Scharf et al. 2000). Previously reported size ranges of blue crab, Callinectes sapidus, found in bonnethead stomachs from the central eastern Gulf of Mexico were also small (0.9–6.0 cm carapace length; Cortés et al. 1996). Bonnetheads in the Gulf of Mexico are similar to elasmobranchs from the northwestern Atlantic in that they do not show a significant increase in the ranges of absolute prey sizes taken with increasing predator size (Scharf et al. 2000). Unlike other small coastal shark species, the bonnethead uses plate-like teeth to crush hard-shelled invertebrates (Wilga and Motta 2000). This mode of prey capture/handling may be more closely related to the size rather than speed of potential prey or experience/efficiency of the predator.
Bonnetheads have one of the lowest percentages of empty stomachs reported for shark species. Previously reported estimates for the bonnethead include 7% by Parsons (1987) and 5% by Cortés et al. (1996). In this study, 1.5% of all stomachs were empty. By comparison, Bethea et al. (2003) reported 24, 49, and 50% empty stomachs for juvenile blacktip, C. limbatus, finetooth, C. isodon, and spinner sharks, C. brevipinna, respectively, collected in the northern Gulf of Mexico. In all these studies, sharks were collected with gillnets, likely ruling out gear bias associated with the proportion of empty stomachs (e.g., sharks collected with longlines generally have a higher proportion of empty stomachs; Wetherbee and Cortés 2004). The low occurrence of empty stomachs suggests that bonnetheads may feed more frequently than other sharks. Although experimental estimates of feeding frequency are rare (Wetherbee and Cortés 2004), feeding in bonnetheads appears to be asynchronous and does not show time preference (Cortés et al. 1996).
Meal size and food type may also be related to the low percentage of empty stomachs found in bonnetheads. Gastric evacuation time increased by as much as 50% when meal size was increased by a factor of 8.4 for scalloped hammerhead sharks, S. lewini (Bush and Holland 2002). Bonnetheads in this study typically had abundant food in the stomach and were most often observed with whole prey items rather than pieces. Food type and its organic composition (i.e., digestibility) influence the rate at which food is ingested and leaves the stomach, and consequently affects the degree of vacuity of the stomach. The diet of bonnetheads is dominated by crustaceans, particularly crabs, in most areas. Crabs generally take longer to digest than cephalopods because they have a chitinous exoskeleton. In turn, both crabs and octopi take longer to digest than fish (Jackson et al. 1992). In a study on the sandbar shark, C. plumbeus, Medved et al. (1988) found gastric evacuation took 20 h longer when sharks were fed crab than when sharks were fed fish. Berens (2005) showed a 5–6 h lag in digestion time for gag, Mycteroperca microlepis, when fed crustaceans versus fish prey.
Determining consumption and daily ration through the bioenergetic approach can be problematic due to the reliability of the estimates of growth and metabolism (Ney 1993). However, the major inputs used in this model were derived from bonnethead-specific experiments and life history information. Daily ration estimates for bonnetheads using this bioenergetic model were similar to previous values obtained using gastric evacuation laboratory experiments and examination of stomach contents of sharks caught in the wild. Tyminski et al. (1999) reported estimates of daily ration for bonnetheads collected off Tampa Bay of 2.2–4.3 %BW day-1. Sensitivity analysis in the present study indicated that all bioenergetic models for bonnetheads were most sensitive to parameters that determined metabolic rate (i.e., mass of an individual and temperature). These results reinforce previous analyses that demonstrated the parameters determining metabolic rate (e.g., swimming speed) have the greatest effect on predicted consumption rates (Bartell et al. 1986; Essington 2003).
Bonnetheads in the eastern Gulf of Mexico display latitudinal variation in life history traits; growth rate, maximum size, and age-at-maturity were found to increase with an increase in latitude of 5° (Lombardi-Carlson et al. 2003). This trend is thought to be the result of local phenotypic responses to environmental conditions rather than differences in mitochondrial DNA (Lombardi-Carlson et al. 2003). The opposite trend was found for estimates of consumption. Regardless of whether or not we assumed plant material was part of the diet, juvenile and adult bonnetheads in northwest Florida had the lowest estimate of daily ration and those in Florida Bay had the highest. Parsons (1993) originally hypothesized that bonnetheads continuously exposed to warmer temperatures, such as those in Florida Bay, would have elevated standard metabolism that would incur additional energy consumption requirements to maintain growth and reproduction. Bonnetheads that continuously occupy warmer waters on a yearly basis may also use more of their consumed energy for maintenance rather than for somatic growth and reproduction, leading to lower growth rates and reproductive output over their entire lifespan. The results from the bioenergetic models in this study tend to support this hypothesis. In an individual-based bioenergetic model, Neer et al. (2007) determined cownose rays, Rhinoptera bonasus, consume approximately 11% more energy per day in warmer water temperature scenarios to compensate for the increased metabolic costs. This was further reflected in decreases in growth rates under those conditions. Faster individual growth rates predicted for the cooler water scenarios allowed individuals to attain a larger size more quickly, which, with weight-dependent mortality, led to greater survival at the population level (Neer et al. 2007).
Plant material was often the second-most important diet category in life stages across sites, making up more than 15 %IRIDC in six of the seven life stage-area combinations, more than 30 %IRIDC in 4 of the 7 combinations, and 62 %IRIDC in YOY diet in northwest Florida. In the past, plant material in bonnethead diet was considered incidental to prey capture and dismissed as reflective of benthic feeding habits (Cortés et al. 1996). Undigested plant material was rarely observed in the intestine or spiral valve of bonnetheads in this study and observations of newly captive (<8 h) bonnetheads in aquaria showed no evidence of plant material in the feces (J.K. Carlson pers. observ.). Even if it is ingested incidentally, these observations could indicate that plant material is being broken down and possibly assimilated. Carbohydrases and lipases that may aid in the digestion of plant material have been identified in pancreatic secretions from several elasmobranchs (Sullivan 1907; Babkin 1929, review in Cortés et al. 2007).
Evidence of herbivory in marine teleosts is well documented. For example, seagrass is common in the diet of parrotfish, Sparisoma radians (Lobel and Ogden 1981; Goecker et al. 2005), pinfish, Lagodon rhomboides, (Weinstein et al. 1982), and halfbeaks, Hyporhamphus sp. (Klumpp and Nichols 1983). Montgomery and Targett (1992) found that pinfish assimilate a significant proportion of the eelgrass, Zostera marina, they consume. Whether or not bonnetheads assimilate plant material is unknown; however, daily ration was higher when plant material was assumed to be part of the diet. More research is necessary to fully evaluate the role plant material plays in the overall nutrition of the bonnethead.
References
Babkin BP (1929) Studies on the pancreatic secretion in skates. Biol Bull 57:272–291
Bartell SM, Breck JE, Gardner RH, Brenkert AL (1986) Individual parameter perturbation and error analysis of fish bioenergetics models. Can J Fish Aquat Sci 43:160–168
Berens EJ (2005) Gastric evacuation and digestion state indices for gag Mycteroperca microlepis consuming fish and crustacean prey. MS thesis, The University of Florida, Gainesville
Bethea DM, Buckel JA, Carlson JK (2003) Foraging ecology of the early life stages of four sympatric shark species. Mar Ecol Prog Ser 268:245–264
Bethea DM, Carlson JK, Buckel JA, Satterwhite M (2006) Ontogenetic and site-related trends in the diet of the Atlantic sharpnose shark Rhizoprionodon terraenovae from the northeast Gulf of Mexico. Bull Mar Sci 78:287–307
Brafield AE, Solomon DJ (1972) Oxy-calorific coefficients for animals respiring nitrogenous substrates. Comp. Biochem Physiol 43A:837–841
Bush AC, Holland KH (2002) Food limitation in a nursery area: estimates of daily ration in juvenile scalloped hammerhead, Sphyrna lewini, in Kaneohe Bay, Oahu, Hawaii. J Exp Mar Biol Ecol 278:157–178
Carlson JK (2007) Modeling the role of sharks in the trophic dynamics of Apalachicola Bay. Am Fish Soc Symp Ser 50:281–300
Carlson JK, Brusher JH (1999) An index of abundance for coastal species of juvenile sharks from the northeast Gulf of Mexico. Mar Fish Rev 61:37–45
Carlson JK, Parsons GR (1997) Age and growth of the bonnethead, Sphyrna tiburo, from northwest Florida, with comments on clinal variation. Environ Biol Fish 50:331–341
Carlson JK, Parsons GR (1999) Seasonal differences in routine oxygen consumption rates of the bonnethead shark. J Fish Biol 55:876–879
Carlson JK, Parsons GR (2003) Respiratory and hematological responses of the bonnethead shark, Sphyrna tiburo, to acute changes in dissolved oxygen. J Exp Mar Biol Ecol 294:15–26
Carlson JK, Goldman K, Lowe C (2004) Metabolism, energetic demands, and endothermy. In: Carrier JC, Musick JA, Heithaus MR (eds) Biology of sharks and their relatives. CRC Press, Boca Raton, pp 203–224
Cortés E (1997) A critical review of methods of studying fish feeding based on analysis of stomach contents: application to elasmobranch fishes. Can J Fish Aquat Sci 54:726–738
Cortés E (1999) Standardized diet compositions and trophic levels of sharks. ICES J Mar Sci 56:707–717
Cortés E (2004) Life history patterns, demography and population dynamics. In: JC Carrier JA Musick MR Heithaus (eds) Biology of sharks and their relatives. CRC Press, Boca Raton, pp 449–469
Cortés E, Gruber SH (1990) Diet, feeding habits and estimates of daily ration of young lemon sharks, Negaprion brevirostris (Poey). Copeia 1990:204–218
Cortés E, Manire CA, Hueter RE (1996) Diet, feeding ecology, and diel feeding chronology of the bonnethead shark, Sphyrna tiburo, in southwest Florida. Bull Mar Sci 58:353–367
Cortés E, Papastamatiou YP, Carlson JK, Ferry-Grahm L, Wetherbee BM (2007) An overview of the feeding ecology and physiology of elasmobranch fishes. In: (eds) Feeding and digestive functions in fishes. Science Publishers, (in press)
Dowd WW, Brill RW, Bushnell PG, Musick JA (2006) Estimating consumptions rates if juvenile sandbar sharks (Carcharhinus plumbeus) in Chesapeake Bay, Virginia, using a bioenergetics model. Fish Bull 104:332–342
Ellis JK (2003) Diet of the sandbar shark, Carcharhinus plumbeus, in Chesapeake Bay and adjacent waters. MS thesis, The College of William and Mary, Williamsburg
Essington TE (2003) Development and sensitivity analysis of bioenergetic models for skipjack tuna and albacore: a comparison of alternative life histories. Trans Am Fish Soc 132:759–770
Ferry LA, Cailliet GM (1996) Sample size and data: Are we characterizing and comparing diet properly? In: Makinlay D, Shearer K (eds) Feeding ecology and nutrition in fish. Proceedings of the symposium on the feeding ecology and nutrition in fish, international congress on the biology of fishes, San Francisco. American Fisheries Society
Goecker ME, Heck KL, Valentine JF (2005) Effects of nitrogen concentration in turtlegrass Thalassia testudinum on consumption by the bucktooth parrotfish Sparisoma radians. Mar Ecol Prog Ser 286:239–248
Gotelli NJ, Entsminger GL (2005) EcoSim: Null models software for ecology. Version 7.72. Acquired Intelligence/Kesey-Bear, Burlington. Available at: http://www.garyentsminger.com/ecosim/index.htm
Graham JH, Vrijenhoek RC (1988) Detrended correspondence analysis of dietary data. Trans Am Fish Soc 117:29–36
Hansen MJ, Bosclair D, Brandt SB, Hewett SW, Kitchell JF, Lucas MC, Ney JJ (1993) Applications of bioenergetics models to fish ecology and management: where do we go from here? Trans Am Fish Soc 122:1019–1030
Jackson S, Place AR, Seiderer LJ (1992) Chitin digestion and assimilation by seabirds. Auk 109:758–770
Kitchell JF, Stewart DJ, Weininger D (1977) Application of a bioenergetics model to yellow perch (Perca flavescens) and walleye (Stizostedion vitreum vitreum). J Fish Res Board Can 34:1922–1935
Kitchell JF, Essington EE, Boggs CH, Schindler DE, Walters CJ (2002) The role of sharks and longline fisheries in a pelagic ecosystem of the central Pacific. Ecosystems 5:202–216
Klumpp DW, Nicols PD (1983) Nutrition of the southern garfish Hyporhamphus melanochir: gut passage rate and daily comsumption of two food types and assimilation of seagrass components. Mar Ecol Prog Ser 12:207–216
Krebs CJ (1999) Ecological methodology. Version 5.1. Department of Zoology, University of British Columbia. Available at: http://www.nhsbig.inhs.uiuc.edu/wes/krebs.html
Lobel PS, Ogden JC (1981) Foraging by the herbivorous parrotfish Sparisoma radians. Mar Bio 64:173–183
Lombardi-Carlson LA, Cortés E, Parsons GR, Manire CA (2003) Latitudinal variation in life-history traits of bonnethead shark, Sphyrna tiburo, (Carcharhiniformes:Sphyrnidae) from the eastern Gulf of Mexico. Mar Freshw Res 54:875–883
Lowe CG (2001) Metabolic rates of juvenile scalloped hammerhead sharks (Sphyrna lewini). Mar Biol 139:447–453
Lowe CG (2002) Bioenergetics of free-ranging juvenile scalloped hammerhead sharks (Sphyrna lewini) in Kane’ohe Bay, O’ahu, HI. J Exp Mar Biol Ecol 278:141–156
McCandless CT, Pratt Jr HL, Kohler NE (2002) Shark nursery grounds of the Gulf of Mexico and the east coast waters of the United States: an overview. An internal report to NOAA’s Highly Migratory Species Office. NOAA Fisheries Narragansett Lab
Medved RJ, Stillwell CE, Casey JG (1988) The rate of food consumption of young sandbar sharks (Carcharhinus plumbeus) in Chincoteague Bay, Virginia. Fish Bull US 83:395–402
Montgomery JLM, Targett TE (1992) The nutritional role of seagrass in the diet of omnivorous pinfish Lagodon rhomboids (L.). J Exp Mar Biol Ecol 158:37–57
Musick JA, Harbin MM, Berkeley SA, Burgess GH, Eklund AM, Findley L, Gilmore RG, Golden JT, Ha DS, Huntsman GR, McGovern JC, Parker SJ, Poss SG, Sala E, Schmidt TW, Sedberry GR, Weeks H, Wright SG (2000) Marine, estuarine, and diadromous fish stocks at risk for extinction in North America (exclusive of Pacific salmonids). Fisheries 25:6–30
National Marine Fisheries Service (NMFS) (1999a) Fishery management plan of the Atlantic tunas, swordfish and sharks. Silver Spring. National oceanic and atmospheric administration, US Department of Commerce, Washington
National Marine Fisheries Service (NMFS) (1999b) Ecosystem-based fishery management: a report to congress by the ecosystems principles advisory panel. national oceanic and atmospheric administration, US Department of Commerce, Washington
National Research Council (NRC) (1998) Improving fish stock assessments. National Academy Press, Washington
Neer JA, Rose KA, Cortés E (2007) Simulating the effects of temperature on individual and population growth of the cownose ray, Rhinoptera bonasus: a coupled bioenergetics and matrix modeling approach. Mar Ecol Prog Ser 329:211–223
Ney JJ (1993) Bioenergetics modeling today: Growing pains on the cutting edge. Trans Am Fish Soc 122:736–748
Parsons GR (1987) Life history and bioenergetics of the bonnethead shark, Sphyrna tiburo (Linnaeus): A comparison of two populations. PhD dissertation, University of South Florida, Tampa
Parsons GR (1990) Metabolism and swimming efficiency of the bonnethead shark, Sphyrna tiburo. Mar Biol 104:363–367
Parsons GR (1993) Geographic variation in reproduction between two populations of the bonnethead shark, Sphyrna tiburo. Environ Biol Fish 38:25–35
Parsons GR, Carlson JK (1998) Physiological and behavioral responses to hypoxia in the bonnethead shark, Sphyrna tiburo: routine swimming and respiratory regulation. Fish Physiol Biochem 19:189–196
Pianka ER (1976) Competition and niche theory. In: May RM (Ed) Theoretical ecology: principals and applications. W. D. Saunders, Philadelphia
Pinkas L, Oliphant MS, Iverson ILK (1971) Food habits of albacore, bluefin tuna, and bonito in Californian waters. Calif Fish Game 152:1–105
Scharf FS, Juanes F, Roundtree RA (2000) Predator-prey relationships of marine fish predators: interspecific variation and effects of ontogeny and body size on trophic-niche breadth. Mar Ecol Prog Ser 208:229–248
Schindler DE, Essington TE, Kitchell JF, Boggs C, Hilborn R (2002) Sharks and tunas: fisheries impacts on predators with contrasting life histories. Ecol Appl 12:735–748
Sims DW, Davies SJ (1994) Does specific dynamic action (SDA) regulate return of appetite in the lesser spotted dogfish, Scyliorhinus canicula? J Fish Biol 45:341–348
Steimle FW, Terranova RJ (1985) Energy equivalents of marine organisms from the continental shelf of the temperate northwest Altantic. J Northw Atl Fish Sci 6:117–124
Stevens JD, Bonfil R, Dulvy NK, Walker PA (2000) The effect of fishing on sharks, rays, and chimaeras (chondrichthyans), and the implications for marine ecosystems. ICES J Mar Sci 57:476–494
Stillwell CE, Kohler NE (1982) Food, feeding habits, and estimates of daily ration of the shortfin mako (Isurus oxyrinchus) in the northwest Atlantic. Can J Fish Aquat Sci 39:407–414
Stillwell CE, Kohler NE (1993) Food habits of the sandbar shark, Carcharhinus plumbeus, off the US northeast coast, with estimates of daily ration. Fish Bull US 91:138–150
Sullivan MS (1907) The physiology of the digestive tract of elasmobranchs. US Fish Wildl Serv Fish Bull 27:3–27
Tenore KR (1981) Organic nitrogen and caloric content of detritus. Estuar Coast Shelf Sci 12:39–47
Thayer GW, Schaff WE, Angelovic JW, LaCroix MW (1973) Caloric measurements of some estuarine organisms. Fish Bull US 71:289–296
Tyminski JP, Cortés E, Manire CA, Hueter RE (1999) Gastric evacuation and estimates of daily ration in the bonnethead shark, Sphyrna tiburo. American Society of Ichthyologists and Herpetologists. 79th Annual Meeting, Pennsylvania State University, University Park, June 24–30
Wetherbee BM, Cortés E (2004) Food consumption and feeding habits. In: Carrier JC, Musick JA, Heithaus MR (eds) Biology of sharks and their relatives. CRC Press, Boca Raton, pp 225–246
Wetherbee BM, Gruber SH (1993) Absorption efficiency of the lemon shark Negaprion brevirostris at varying rates of energy intake. Copeia 1993:416–425
Wilga CD, Motta PJ (2000) Durophagy in sharks: Feeding mechanics of the hammerhead Sphyrna tiburo. J Exp Biol 203:2781–2796
Winberg GG (1960) Rate of metabolism and food requirements of fishes. Fish Res Board Can Trans Ser 194:1–202
Winemiller KO, Pianka ER (1990) Organization and natural assemblages of desert lizards and tropical fishes. Ecol Monogr 60:27–55
Weinstein MP, Heck Jr KL, Giebel PE, Gates JE (1982) The role of herbivory in pinfish (Lagodon rhomboids): a preliminary investigation. Bull Mar Sci 32:791–795
Yamaguchi A, Taniuchi T (2000) Food variations and ontogenetic dietary shift of the starspotted-dogfish Mustelus manazo at 5 locations in Japan and Taiwan. Fish Sci 66:1039–1048
Acknowledgments
We thank the staff and personnel of the NOAA Fisheries Panama City Laboratory in Panama City, Florida. Special thanks to go J. Tyminski for supervising the collection of sharks in Tampa Bay and Florida Bay. This project would not have been possible without the hard work of several volunteers, interns, and technicians at Mote Marine Laboratory and the NOAA Fisheries Panama City Laboratory. E. Dutton designed and wrote the program used to calculate cumulative prey curves. This project was funded in part by the United States Environmental Protection Agency (US EPA) grant #R826128-01-0. This manuscript has not received the US EPA’s required peer and policy review; therefore, it does not necessarily reflect the views of the agency. In addition, reference to trade names does not imply endorsement by NOAA Fisheries.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by J. P. Grassle.
Electronic supplementary material
Below is the link to the electronic supplementary material.
{kind=link}
{kind=link}
{kind=link}
Rights and permissions
About this article
Cite this article
Bethea, D.M., Hale, L., Carlson, J.K. et al. Geographic and ontogenetic variation in the diet and daily ration of the bonnethead shark, Sphyrna tiburo, from the eastern Gulf of Mexico. Mar Biol 152, 1009–1020 (2007). https://doi.org/10.1007/s00227-007-0728-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-007-0728-7