
Abstract
The Permian bryozoan fauna of the Surmaq Formation exposed in a section near Kuh-e Hambast (Hambast Mountains, central Iran) includes ten species. Four species (Fistulipora sawatai Sakagami 1999, Fistulipora takauchiensis Sakagami 1961, Fistulipora monticulosa Nikiforova 1933, and Eridopora parasitica, Waagen and Wentzel 1886) indicate a Middle Permian (Murgabian) age of the formation. Six additional taxa, three cystoporates Fistulipora sp. 1, Fistulipora sp. 2, and Fistuliporidae gen. et sp. indet., as well as three trepostomes Dyscritella sp., Trepostomata gen. et sp. indet. 1 and Trepostomata gen. et sp. indet. 2 could not be identified at the genus and species level. The investigated fauna refers to the Middle Permian of Thailand, Pakistan, Indonesia, and Japan.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Bryozoans are usually abundant in Permian sediments of Iran. However, their diversity as well as their stratigraphic and palaeobiogeographic importance have not been thoroughly investigated. Few publications have been made on Iranian Permian bryozoans in the last century. Fantini Sestini (1965) described three species from the Permian Ruteh Limestone of the Elborz Mountains (northern Iran). Later, Sakagami (1980) described 15 species from the Permian of central Iran (Abadeh region). This fauna displayed a close relation to the Dzhulfian (= Changhsingian) stage, as well as connections to the Permian of Pamir, Russian platform, Salt Range, Kashmir, South China, and Timor.
Four bryozoan faunas from the Permian Jamal Formation of central and northeast Iran are described in a recent series of publications: (1) Thirty-one bryozoan species were identified from the Permian Jamal Formation exposed in the Lakaftari area, south of the town of Bagher-Abad, northeast of Esfahan (central Iran) (Ernst et al. 2006a). This fauna is regarded as Middle Permian (Murgabian) in age, and shows palaeobiogeographic connections to Australia, Indonesia, Thailand, and Transcaucasia; (2) Six bryozoan species were described from the Permian Jamal Formation, of Kuh-e Bagh-e Vang (Shotori Mountains, northeast Iran) (Ernst et al. 2006b). This association indicates a Lower Permian age and shows palaeogeographic connections with bryozoan faunas known from Australia and Urals Mountains; (3) Nine bryozoan species were described from the Permian Jamal Formation outcropped near the village of Chiruk, Shotori Mountains, northeast Iran (Ernst et al. 2008). The bryozoan Filiramoporina sp. from the last faunal association shows relations to the Lower Permian of Kansas, North America. Other species are known from the Permian of Afghanistan, Indonesia, Russian Plate, and the Far East; (4) Seven bryozoan species were described from the Jamal Formation exposed near Deh-e Mohammad, Shotori Mountains (northeast Iran) displaying palaeobiogeographic connections to the Lower Permian of Pamir (Tajikistan), Indonesia, Thailand, and Kansas (North America) (Ernst et al. 2009).
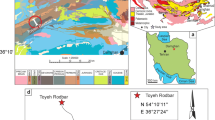
The present paper is a further contribution to the knowledge of the Permian bryozoans of Iran. The investigation is based on material collected from a section of the Surmaq Formation exposed near the Kuh-e Hambast (Hambast Mountains, south of the town of Abadeh, central Iran) (Fig. 1a, b). The described bryozoan fauna of the Surmaq Formation from Kuh-e Hambast includes ten species: seven cystoporates and three trepostomates:

a Geographic position of the studied locality, south of the town of Abadeh, marked with an asterisk. b Lithological column of the sampled section showing thick-bedded carbonate units (1–3). The described bryozoan fauna comes from the unit 1
Fistulipora sawatai Sakagami 1999
Fistulipora takauchiensis Sakagami 1961
Fistulipora monticulosa Nikiforova 1933
Fistulipora sp. 1
Fistulipora sp. 2
Eridopora parasitica (Waagen and Wentzel 1886)
Fistuliporidae gen. et sp. indet.
Dyscritella sp.
Trepostomata gen. et sp. indet. 1
Trepostomata gen. et sp. indet. 2
Beside these, unidentifiable fragments of fenestrates were found in thin sections (Spinofenestella sp., Minilya sp., Penniretepora sp.).
The described fauna can be correlated with the Middle Permian of Thailand, Pakistan, Tibet, Indonesia, and Japan, and indicates the Middle Permian (Murgabian) age of the Surmaq Formation from Kuh-e Hambast. Fistulipora sawatai Sakagami 1999 is known from the ?Guadalupian of Thailand. Fistulipora takauchiensis Sakagami 1961 was originally described from the Takauchi Limestone (Lepidolina-Yabeina zone, Middle Permian, Upper Guadalupian of Japan). Fistulipora monticulosa Nikiforova 1933 is known from Gnishik Horizon (Middle Permian, Murgabian) of Transcaucasia, Middle Permian of central Turkey, Jamal Formation (Middle Permian, Murgabian) of Lakaftari area in central Iran as well as Upper Permian (Araxopora araxensis-Horizon) of Abadeh area in central Iran. Eridopora parasitica (Waagen and Wentzel 1886) was reported from the Permian of Indonesia and Thailand, Lower Permian (Artinskian) of Caucasus, Middle Permian of Oman, Lower and Middle Permian of Xizang (Tibet), and the Upper Permian; Pakistan (Salt Range).
Geological setting and geographic position of studied area
The study area lies northwest of the Kuh-e Hambast in the Hambast Mountains, about 20 km southeast of the town of Abadeh (Fig. 1). The Hambast Mountains belong to the so-called Sanandaj-Sirjan structural belt (Stöcklin 1968), a tectonic zone of almost 1,500 km, running NW–SE, more or less parallel to the Zagros belt to the southwest. The Sanandaj-Sirjan zone belongs to the southwest part of the central Iranian plate. It differs from the other part of the Iranian plate in its structural geology, and corresponds to the Zagros belt. The Sanandaj-Sirjan belt is bound by the Zagros belt in the southwest, by the “Main Zagros fault”, and by the central Iranian plate in the northeast by different geological structures, generally NW–SE-trending small basins containing Upper Cenozoic deposits. Lithologically, the occurrence of abundant metamorphic rocks of the Sanandaj-Sirjan zone differentiates this zone from the other central Iranian plates and from the neighboring Zagros belt.
The stratigraphic subdivision of Permian sediments of Hambast Mountains was proposed by Taraz (1969, 1974), who recognized seven stratigraphical units. Based on foraminiferal investigations, the “Iranian-Japanese Research Group” (1981) introduced the name Surmaq Formation (after the nearest small town to the mountains) for units 1–3, Abadeh Formation for units 4–5, and finally the Hambast Formation for units 6–7 of Taraz. Kobayashi and Ishii (2003) restudied the foraminifera of the region, based on several sections. Apparently, the section N-R of the latter authors corresponds to or is very close to our section from where the bryozoans, described in this paper, were collected. According to their stratigraphy, our sampled section corresponds to the Surmaq Formation (unit 1 of Taraz) and indicates a Middle Permian (Murgabian) age for the fossils in our collection.
The studied section is about 120 m thick and starts with sandy limestones at its base. Three thick-bedded carbonate units (1–3) are distinct in the section (Fig. 1b). The first one is about 28 m thick, and represents reefs or reefal limestones. Hypercalcified sponges, Tubiphytes and Tubiphytes-like organisms, bryozoans, and corals are the most abundant reef-building organisms. The sponge fauna of the Hambast Formation was described by Senowbari-Daryan et al. (2007). The described bryozoan fauna comes from the same unit. The second thick-bedded limestone unit (2) is about 15 m thick and contains abundant brachiopods. The thick-bedded carbonate unit (3) is represented by dolomites at the top of the section.
Materials and methods
Bryozoans were investigated in thin sections using a transmitted light microscope. Statistics were summarized with arithmetic mean, sample standard deviation, coefficient of variation, and minimum and maximum value. The studied material includes 20 large thin sections and is deposited in the collections of the Palaeoenvironment Research Division, University of Erlangen-Nuremberg (under the label “Senowbari-Daryan, Perm Hambast, thin sections “Ham-…”, “PCH-” and “PA-…”)”.
Systematic palaeontology
Phylum: Bryozoa Ehrenberg 1831
Class: Stenolaemata Borg 1926
Order: Cystoporata Astrova 1964
Family: Fistuliporidae Ulrich 1882
Genus: Fistulipora M’Coy 1849
Fistulipora sawatai Sakagami 1999

a–e Fistulipora sawatai Sakagami 1999. a Oblique section through a branched colony. b Tangential section showing autozooecial and megazooecial apertures. c Tangential section showing autozooecial and megazooecial apertures. d Longitudinal section showing secondary overgrowth, autozooecial chambers, and vesicles. e Tangential section showing autozooecial and megazooecial apertures. f–h Fistulipora takauchiensis Sakagami 1961. f Tangential section showing autozooecial apertures. g, h Longitudinal section of a massive colony. i Fistulipora sp. 1, longitudinal section showing secondary overgrowth, autozooecial chambers, and vesicles
1999 Fistulipora sawatai Sakagami: 83–84, pl. 19, Figs. 1–3
Material
Five colonies Ham10, PA/0/1, PA/0/2, PA/4, and PA/22.
Description
Encrusting and ramose colonies. Encrusting colonies 0.48–1.95 mm in thickness. Secondary overgrowth common. Branches 4.7–5.0 mm in diameter. Autozooecial apertures rounded to oval, spaced 3.5–4.5 in 2 mm on the colony surface in growth direction, separated usually by 1–2 rows of vesicles. Lunaria are prominent, with ends projecting deeply into the autozooecial chamber. Basal diaphragms are rare to absent. Vesicles polygonal in cross section, having rounded roofs in longitudinal section, spaced 15–20 in 1-mm colony thickness. The outer granular skeleton is well developed. Megazooecia are large, and appear randomly throughout the colony, and have large triangular or horseshoe-shaped lunaria.
Comparison
The present species is similar to Fistulipora sawatai Sakagami 1999 from the Middle Permian (? Guadalupian) of Thailand in the kind of lunaria and presence megazooecia. Fistulipora zhejiangensis Lu 1986 from the Lengwu Member of the Maokou Stage (? Upper Permian) of central Hunan—Western Zhejiang (China) is similar to the present species in presence of characteristic megazooecia. However, megazooecia in F. zhejiangensis are larger than those in the present species (0.55–0.75 vs. 0.30–0.43 mm). Furthermore, F. zhejiangensis differs from the present material in the presence of abundant autozooecial diaphragms.
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran. Middle Permian (? Murgabian); Khao Hin Kling area, north-central Thailand.
Fistulipora takauchiensis Sakagami 1961
1961 Fistulipora takauchiensis Sakagami: 17–18, pl. 3, figs. 1–2, pl. 4, figs. 1–5
Material
Three colonies Ham4, PA/15, and PA.
Description
Massive colonies, 10–12 mm in diameter. Autozooecia are long, and radiate from the center of the colony. Autozooecial apertures are rounded to oval, and spaced 4–5 in 2 mm on the colony surface in the growth direction, usually separated by 1–3 rows of vesicles. Lunaria are large, with ends projecting deeply into the autozooecial chamber. Basal diaphragms are common to abundant, planar, thin, 4–7 spaced in 1 mm of autozooecial length. Vesicles are large, polygonal in cross section, having rounded roofs in longitudinal section, 10–18 spaced in 1-mm colony thickness. The outer granular skeleton is weakly developed.
Comparison
Fistulipora takauchiensis Sakagami 1961 is similar to F. crescens Crockford 1944 from the Noonkanbah Series (Lower Permian) of Western Australia. However, it differs from F. crescens in having smaller autozooecia (0.19–0.35 vs. 0.29–0.43 mm in F. crescens).
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran. Takauchi Limestone (Lepidolina-Yabeina zone), Middle Permian (Midian); Japan.
Fistulipora monticulosa Nikiforova 1933

a Fistulipora sp. 1, tangential section showing autozooecial apertures. b–d Fistulipora sp. 2. b Oblique section through a branched colony. c Longitudinal section through a branched colony. d Tangential section showing autozooecial apertures. e–j, h Fistulipora monticulosa Nikiforova 1933. e Longitudinal section showing autozooecial chambers and vesicles, f tangential section showing autozooecial apertures, h autozooecial aperture. g Eridopora parasitica (Waagen and Wentzel 1886), tangential section showing autozooecial apertures
1933 Fistulipora monticulosa Nikiforova: 10, pl. 1, figs. 9–15, text-figs. 3, 4
1970 Fistulipora monticulosa Nikiforova 1933—Morozova: 61–62, pl. 1, fig. 2
1976 Fistulipora sp. cf. F. monticulosa Sakagami: 400, pl. 42, figs. 3, 4
1980 Fistulipora pseudomonticulosa Sakagami: 272, pl. 31, figs. 4–6
2006a Fistulipora monticulosa Nikiforova 1933—Ernst, Senowbari-Daryan and Hamedani: 546–547, figs. 2a, b, d, e, g
Material
Single colony PCH28.
Description
Encrusting colonies 0.93 mm in thickness. Autozooecial apertures are rounded to oval, spaced 3.8–4.5 in 2 mm on the colony surface in growth direction, and are usually separated by a single row of vesicles. Lunaria distinct, horseshoe-shaped to slightly triangular. Basal diaphragms are absent. Vesicles polygonal in cross section, moderately large, flat, and have rounded roofs in longitudinal section, spaced 9–10 in 1-mm colony thickness. Outer granular skeleton is well developed.
Comparison
Fistulipora monticulosa is similar to F. timorensis Bassler 1929, which is widely distributed in Permian rocks of Asia (Indonesia, Thailand, Primorye, Iran). F. timorensis has larger apertures (aperture width 0.35 vs. 0.30 mm at average in F. monticulosa).
Occurrence
Gnishik Horizon, Middle Permian (Murgabian); Transcaucasia (Nakhichevan). Middle Permian; Sainbeyli, Central Turkey. Upper Permian (Araxopora araxensis-Horizon); Abadeh, central Iran. Jamal Formation, Middle Permian (Murgabian); Lakaftari, central Iran. Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Fistulipora sp. 1
Material
Single colony Ham6.
Description
Encrusting colony 0.24–0.29 mm in thickness. Secondary overgrowth common. Autozooecial apertures rounded to oval, spaced 4.3–4.4 in 2 mm on the colony surface in growth direction, usually separated by 1–3 rows of vesicles. Lunaria distinct, horseshoe-shaped. Basal diaphragms are absent. Vesicles are polygonal in cross section, flat, and have rounded roofs in longitudinal section, and are spaced 15–17 in 1-mm colony thickness. Outer granular skeleton is well developed.
Comparison
Fistulipora sp. 1 is similar to F. rutogensis Xia, 1991 from the Chainaha Formation (Middle Permian) of Rutog County, China. The present species differs from F. rutogensis in having weakly developed lunaria and smaller autozooecia (autozooecial apertures width 0.20–0.26 vs. 0.24–0.30 mm in F. rutogensis).
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Fistulipora sp. 2
Material
Three colonies PA/27/(1–3).
Description
Branched colonies, 3.9–4.8 mm in diameter, with 2.9–3.0-mm-wide endoozones and 0.5–0.9-mm-wide exozones. Autozooecial apertures are rounded to oval, spaced 3.5–4.5 in 2 mm on the colony surface in growth direction, usually separated by 1–2 rows of vesicles. Lunaria distinct, horseshoe-shaped. Basal diaphragms are rare to absent. Vesicles are polygonal in cross section and have rounded roofs in the longitudinal section, spaced 10–15 in 1-mm colony thickness. Outer granular skeleton is well developed.
Comparison
Fistulipora sp. 2 is similar to F. maanensis Yang 1956 from the Changhsing Formation (Upper Permian) of Hupeh, China. The latter species has thicker colonies (branch diameter 15.0 vs. 2.9–3.0 mm in present species) and larger apertures (autozooecial aperture width 0.26–0.30 vs. 0.17–0.28 mm in the present species).
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Genus: Eridopora Ulrich 1882
Eridopora parasitica (Waagen and Wentzel 1886)
(=E. major Bassler 1929)

a–c Eridopora parasitica (Waagen and Wentzel 1886). a Tangential section showing autozooecial apertures, b longitudinal section, c tangential section showing autozooecial aperture with a lunarium and lunarial nodes. d–h Fistuliporidae gen. et sp. indet. d Tangential section, e longitudinal section of a branched colony, f oblique section through a branched colony, g central part of the branch displaying hemiphragms (arrows) in autozooecial apertures, h tangential section. i, j Dyscritella sp. i Longitudinal section of an encrusting colony, j cross section of an encrusting colony
1886 Fistulipora parasitica Waagen and Wentzel: 923, pl. 45, fig. 6, pl. 105, figs. 1–4
1929 Eridopora major Bassler: 52, pl. 225 (1), figs. 1–4
1975 Eridopora major Bassler 1929—Gorjunova: 45–46, pl. 3, fig. 1
1980 Eridopora cf. parasitica (Waagen and Wentzel 1886)—Sakagami: 273, pl. 31, figs. 7–9
1986 Eridopora parasitica (Waagen and Wentzel 1886)—Xia: 232, pl. 132, figs. 8–9, pl. 15, figs. 5, 7
1988 Eridopora major Bassler 1929—Sakagami in Yanagida: pl. 12, fig. 4, pl. 14, fig. 1
1988 Eridopora? sp. indet.—Sakagami in Yanagida: pl. 15, figs. 1–2
1991 Eridopora major Bassler 1929—Xia: 189, pl. 7, figs. 8–9
1999 Eridopora parasitica (Waagen and Wentzel 1886)—Sakagami: 84, pl. 19, figs. 4–6
1997 Eridopora parasitica (Waagen and Wentzel 1886)—Sakagami and Pillevuit: 206, figs. 2–3
2000 Eridopora parasitica (Waagen and Wentzel 1886)—Sakagami: 146–147, figs. 4.1–2
Material
Six colonies PA/30, PA/7/2, PA/32/2, PA/32/3, PA/32/5, and Ham7.
Description
Encrusting colonies, 0.8–3.6 mm thick. Autozooecial apertures are rounded to oval, with well-developed triangular lunaria, spaced 2–4 in 2 mm at the colony surface. Lunaria large, prominent, 0.20 mm thick in their middle part, and contain 6–7 nodes. Autozooecial diaphragms are abundant, 7–12 spaced in 1 mm of autozooecial length, planar, thin. Vesicles are small, angular, separating autozooecia usually in 2 rows, spaced 12–16 in 1 mm of autozooecial length. Distinct polygons developed by keels on the colony surface, enclosing apertures.
Comparison
Eridopora parasitica (Waagen and Wentzel 1886) differs from E. oculata Bassler 1929 in possessing larger apertures and thicker colonies (apertures width 0.37 vs. 0.28 mm on average in E. oculata).
Occurrence
Permian; Timor, Indonesia. Permian; Thailand. Lower Permian (Artinskian); Caucasus. Middle Permian; Oman. Upper Xarla Formation, Lower Permian; Xainza, northern Xizang (Tibet). Chainaha Formation, Middle Permian; north-western Xizang (Tibet). Middle Permian (Murgabian); Irian Jaya, Indonesia. Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran. Upper Permian; Pakistan (Salt Range).
Fistuliporidae gen. et sp. indet.
Material
Three colonies Ham7, Ham9, and PA/4.
Description
Small encrusting, globular, and apparently branched colonies. Autozooecia growing from a median axis in branched colonies (Fig. 4e–f). Autozooecial apertures are rounded to rhombic. Lunaria well developed, horseshoe-shaped. Vesicles are low, with rounded flats, are locally abundant, but locally absent or few. Long curved hemiphragms in autozooecia.
Comparison
The present species has general features of Fistulipora, but differs from it by sparse vesicles and the presence of curved hemiphragms.
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Order: Trepostomata Ulrich 1882
Suborder: Amplexoporina Astrova 1965
Family: Dyscritellidae Dunaeva and Morozova 1967
Genus: Dyscritella Girty 1911
Dyscritella sp.

a Dyscritella sp., tangential section. b–d Trepostomata gen. et sp. indet. 1. b Longitudinal section displaying walls with tubules (arrow), c longitudinal section displaying hemiphragms (arrows), d tangential section displaying tubules in autozooecial walls (arrows). e–h Trepostomata gen. et sp. indet. 2. e Longitudinal section, showing mesozooecia and autozooecium, f longitudinal section showing autozooecia, mesozooecia, acanthostyles and secondary overgrowth, g fragment of a massive colony, h tangential section, showing autozooecial apertures, acanthostyles and mesozooecia
Material
Two colonies Ham9 and PA/4.
Description
Encrusting colony, 0.36–0.50 mm thick. Endozones short. Autozooecia growing from epitheca, inclined in endozone, intersecting colony surface at right angles. Autozooecial diaphragms rare, planar. Autozooecial apertures are polygonal, 8–11 spaced in 2 mm. Autozooecial walls are 0.036–0.048 mm thick in exozone and display fine reversal U-shaped lamination. Acanthostyles are abundant, 3–5 surround each autozooecial aperture, and have narrow hyaline cores and wide laminated sheaths. Exilazooecia rare, more abundant in maculae, restricted to exozone, having polygonal apertures. Maculae consist of larger autozooecia and abundant exilazooecia.
Comparison
The present species is similar to Dyscritella leptosa Ernst et al. 2006a from the Middle Permian of the Lakaftari area of central Iran. However, the latter species has abundant exilazooecia and smaller autozooecial apertures (average aperture width 0.19 vs. 0.12 mm in present material).
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Trepostomata gen. et sp. indet. 1
Material
Two colonies Ham6 and Ham9.
Description
Encrusting colonies, 0.4–1.05 mm thick. Secondary overgrowths common. Autozooecial apertures are rounded to angular. Diaphragms in autozooecia are absent. Hemiphragms common, short. Exilazooecia are abundant, and have rounded to polygonal transverse section shape, and 4-5 surrounding each aperture. Acanthostyles absent. Tubules (“capillaries” of authors) in the autozooecial walls are abundant, and are aggregated in clusters along autozooecial boundaries, 0.02 mm in diameter. Autozooecial walls are indistinctly laminated, 0.012–0.025 mm thick in the endozone; regularly thickened, coarsely laminated, 0.06–0.12 mm thick in exozone.
Comparison
The present material is similar to Stenophragmidium Bassler 1952 in having encrusting colony and hemiphragms (Cleary and Wyse Jackson 2006). However, all species of this genus possess acanthostyles, which are absent in the present material. Hinganella Romantchuk, 1967 shows similar wall structure with tubules aggregated in clusters. However, this genus lacks hemiphragms.
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Trepostomata gen. et sp. indet. 2
Material
Eight colonies Ham9, PA/3/4, PA/4, PA/7/2, PA/22, and PA/27/(1–3).
Description
Massive and encrusting colonies. Colony thickness 0.60–0.75 mm. Autozooecia tubular, with rounded apertures. On average, five apertures spaced in 2 mm at colony surface. Mesozooecia abundant, large, polygonal in cross section, containing abundant closely spaced diaphragms, having straight walls, separating autozooecia in 1–3 rows. Acanthostyles abundant, 4–7 surrounding each autozooecial aperture and occurring in junctions between mesozooecia, having wide laminated sheaths and distinct hyaline cores. Autozooecial walls thin, finely laminated.
Comparison
The present species is similar to Leioclema Ulrich 1882 in having abundant mesozooecia and acanthostyles. However, Leioclema has angular apertures and mainly beaded mesozooecia. Hinaclema Sakagami and Sugimura 1987 is also similar to the present species, but has heterozooecia without diaphragms.
Occurrence
Surmaq Formation, Middle Permian (Murgabian); Hambast Mountains, south of Abadeh, central Iran.
Discussion
Permian Bryozoa are relatively well known worldwide and therefore are often used for palaeobiogeography and stratigraphy. However, bryozoan faunas from the Permian sediments in some regions remain unstudied, despite their abundance and diversity. The progressive investigation of the Permian bryozoans from Iran uncovers new insights in the global and regional palaeobiogeography. The described bryozoan fauna of the Surmaq Formation from Kuh-e Hambast (Hambast Mountains, central Iran) can be correlated with the Middle Permian of Thailand, Tibet, Pakistan, Indonesia, and Japan. From the taxonomic point of view, this fauna is dominated by cystoporates (seven species), followed by trepostomes (three species). Besides these, fragments of three fenestrate genera are also present in studied thin sections. The majority of the described species display an encrusting colony shape (seven species), followed by two ramose and one massive species. Locally, bryozoans seem to contribute significantly to the stabilization of the reefal sediments with their encrusting colonies.
References
Astrova GG (1964) Novyi otryad paleozoiskikh mshanok [New order of Paleozoic Bryozoa] (in Russian). Paleontologicheskij zhurnal 1:22–31
Astrova GG (1965) Morphologia, istoria razvitia i sistema ordovikskikh i siluriiskikh mshanok [Morphology, history of development and system of the Ordovician and Silurian Bryozoa] (in Russian). Tr Paleontologicheskogo Inst Akad Nauk SSSR 106:1–432
Bassler RS (1929) The Permian Bryozoa of Timor. Palaontologie Timor 16(28):37–90
Bassler RS (1952) Taxonomic notes on genera of fossil and Recent Bryozoa. J Wash Acad Sci 42(12):381–385
Borg F (1926) Studies on recent cyclostomatous Bryozoa. Zool Bidr fran Uppsala 10:181–507
Cleary D, Wyse Jackson PN (2006) Stenophragmidium Bassler, 1952 (Trepostomida: Bryozoa) from the Mississippian of Ireland and Britain. Isr J Earth Sci 25(1):1–25
Crockford J (1944) Bryozoa from the Permian of Western Australia. Part I. Cyclostomata and Cryptostomata from the North-West Basin and Kimberly district. Proc Linnean Soc N S Wales 69(3–4):139–175
Dunaeva NN, Morozova IP (1967) Osobennosti razvitiya i sistematicheskoe polozhenie nekotorykh paleozoiskikh trepostomat (in Russian) [Evolution and systematic position of certain Late Palaeozoic Trepostomata]. Paleontologicheskij Zh 4:86–94
Ehrenberg CG (1831) Symbolae Physicae, seu Icones et descptiones Corporum Naturalium novorum aut minus cognitorum, quae ex itineribus per Libyam, Aegiptum, Nubiam, Dongalaam, Syriam, Arabiam et Habessiniam, studia annis 1820–25, redirent. Pars Zoologica 4, Animalia Evertebrata exclusis Insectis: 831 p. Berlin (Mittler)
Ernst A, Senowbari-Daryan B, Hamedani A (2006a) Middle Permian Bryozoa from the Lakaftari area, central Iran. Geodiversitas 28:543–590
Ernst A, Senowbari-Daryan B, Rashidi K (2006b) Lower Permian Bryozoa of the Jamal Formation from Bagh-e Vang (Shotori Mountains, northeast Iran). Facies 52:627–635. doi:10.1007/s10347-006-0084-y
Ernst A, Senowbari-Daryan B, Rashidi K (2008) Permian Bryozoa from the Jamal Formation of Shotori Mountains (northeast Iran). Rev Paleobiologie 27(2):395–408
Ernst A, Senowbari-Daryan B, Rashidi K (2009) Rhabdomesid and cystoporid bryozoans from the Permian of Deh-e Mohammad, Shotori Mountains (north-eastern Iran). Geobios 42(2):133–140
Fantini Sestini FN (1965) The geology of the upper Djadjerud and Lar valleys (north-Iran); II, Palaeontology; bryozoans, brachiopods and molluscs from Ruteh limestone (Permian). Rev Ital Paleontologia 71(1):13–108
Girty GH (1911) New genera and species from the Fayetteville Shale of Arkansas. Ann N Y Acad Sci 20:189–238. doi:10.1111/j.1749-6632.1910.tb55149.x
Gorjunova RV (1975) Permskie mshank Pamira. Tr Paleontologicheskogo Inst Akad Nauk SSSR (in Russian). Permian Bryozoa Pamir 148:1–127
Iranian-Japanese Research Group (1981) The Permian and Lower Triassic system in Abadeh Region, central Iran. Mem Fac Sci Kyoto Univ Ser Geol Min 47(2):61–133
Kobayashi F, Ishii KI (2003) Paleobiogeographic analysis of Yahtashian to Midian fusulinacean faunas of the Surmaq Formation in the Abadeh region, central Iran. J Foramin Res 33:155–165. doi:10.2113/0330155
Lu L (1986) Maokou Stage bryozoans from central Hunan—western Zhejiang. Mem Nanjing Inst Geol Palaeont (in Chinese). Acad Sin 22:103–144
M’Coy F (1849) On some new genera and species of Palaeozoic corals and foraminifera. Ann Mag Nat Hist 3(13):119–136
Morozova IP (1970) Mshanki pozdnei permi [Late Permian Bryozoa] (in Russian). Trudy Paleontologicheskogo Instituta Akademii Nauk SSSR 122:1–347
Nikiforova AI (1933) Verkhnepaleozoiskie mshanki Dzhulfinskogo raiona [Upper Paleozoic Bryozoans of Dshulfa District] (in Russian). Trudy Vsesoyusnogo Geologo-razvedochnogo obyedineniya 364:1–44
Romantchuk TV (1967) Novye mshanki otryada Trepostomata iz verkhnei permi Khabarovskogo kraya [New bryozoans of the Order Trepostomida from the Permian of the Khabarovsk Region] (in Russian). Paleontologicheskij Zhurnal 2:63–69
Sakagami S (1961) Japanese Permian Bryozoa. Paleont Soc Jpn Spec Pap 7:1–58
Sakagami S (1976) Paleobiogeography of the Permian Bryozoa on the basis of the Thai-Malayan district. Contributions to the geology and palaeontology of Southeast Asia. Geol Paleont SE Asia 17:155–172
Sakagami S (1980) Permian Ectoprocta (Bryozoa) from the Abadeh Region, Central Iran. Trans Proc Palaeontol Soc Jpn New Ser 118:269–289
Sakagami S (1999) Permian bryozoans from some localities in the Khao Kling area near Phetchabun, north-central Thailand. Bull Kitakyushu Mus Nat Hist 18:77–103
Sakagami S (2000) Middle Permian Bryozoa from Irian Jaya, Indonesia. Bull Natl Sci Mus Tokyo Ser C (Geology and Paleontology) 26:139–168
Sakagami S, Pillevuit A (1997) Permian bryozoans from the exotic formations in Oman. Paleontol Res 1:200–224
Sakagami S in Yanagida J (and A Research Group 1988). Biostratigraphic Study of Paleozoic and Mesozoic Groups in Central and Northern Thailand. An Interim Report. Department of Geology, Faculty of Sciences, Kyushu University, Kyushu, 46 p
Sakagami S, Sugimura A (1987) Hinaclema, a new Carboniferous bryozoan genus from the Hina Limestone, southwest Japan. Proc Jpn Acad Ser B Phys Biol Sci 63:246–249
Senowbari-Daryan B, Hamedani A, Rashidi K (2007) Sponges from the Permian of Hambast Mountains, south of Abadeh, central Iran. Facies 53:575–614
Stöcklin J (1968) Structural history and tectonics of Iran: a review. Am Assoc Petrol Geol Bull 52:1229–1258
Taraz H (1969) Permo-Triassic section in central Iran. Am Assoc Petrol Geol Bull 53:668–693
Taraz H (1974) Geology of the Surmaq-Deh Bid Area, Abadeh Region, Central Iran. Geol Survey Iran, Tehran, Rep No. 37:1–148
Ulrich EO (1882) American Palaeozoic Bryozoa. J Cincinnati Soc Nat Hist 5:121–175
Waagen W, Wentzel J (1886) Salt-Range Fossils. Productus-Limestone Fossils: Coelenterata. Memoirs of the Geological Survey of India, Paleontologica Indica, Series 13 1(6):835–924
Xia F (1986) Carboniferous and early Permian bryozoans from Xainza northern Xizang. Bull Nanjing Inst Geol Palaeont Acad Sin 10:201–254
Xia F (1991) Early-Middle Permian bryozoans from Rutog region Xizang (Tibet). In: Sun D, Xu J et al (eds) Papers for scientific co-expedition of the Nanjing Institute of Geology and Regional Geological Survey Team Geological Bureau of Xizang Stratigraphy and Palaeontology of Permian, Jurassic and Cretaceous of the Rutog Region Xizang (Tibet) [in Chinese]. Nanjing University Press, Nanjing, pp 166–214
Yang K (1956) Some cyclostomatous Bryozoa from the Permian rocks of western Hupeh. Acta Palaeontol Sin 4(2):163–173
Acknowledgements
This study was carried out within the framework of research project Se 416/1 supported by the Deutsche Forschungsgemeinschaft to B. Senowbari-Daryan. Caroline Buttler, Cardiff, and Hans Arne Nakrem, Oslo, are thanked for helpful comments on bryozoan taxonomy. Patrick Wyse Jackson, Dublin, and Dario Sciunnach, Milan, are thanked for their helpful and constructive reviews.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Ernst, A., Senowbari-Daryan, B. & Rashidi, K. Bryozoa from the Surmaq Formation (Permian) of the Hambast Mountains, south of Abadeh, central Iran. Facies 55, 595–608 (2009). https://doi.org/10.1007/s10347-009-0188-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-009-0188-2