
Abstract
Four bryozoan species were identified from the Mississippian (Viséan) Mobarak Formation of northern Iran: the cystoporates Fistulipora longituba Yang and Lu, Fistulipora djebaglinica Nikiforova, Fistulipora sp., and the trepostome Hinaclema sakagamii Schastlivtseva. The cystoporates are represented by large massive, mainly multilayered colonies, whereas Hinaclema sakagamii encrusts (often also multilayered) on two colonies of Fistulipora. The identified species show palaeobiogeographical relationships to the Mississippian of China, Kazakhstan, and Uzbekistan.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Palaeozoic bryozoans are known from different parts of Iran. However, current knowledge about bryozoan faunas in this country remains largely restricted. Ordovician bryozoans are mentioned in scattered publications, and a description of the Late Ordovician bryozoan fauna from central Iran was provided by Ross et al. (2000). Confirmed records of Silurian bryozoans are also few (summarised in Hairapetian et al. 2017). Devonian records concern mainly Frasnian and Famennian faunas (Bigey in Mistiaen et al. 2001; Ernst and Mohammadi 2009; Ernst et al. 2012, 2017). Carboniferous bryozoans of Iran are poorly known, although their significant abundance and diversity has been recorded and summarised (see Gorjunova 2006; Tolokonnikova and Yazdi-Moghadam 2013). In contrast, the Permian bryozoans of Iran have been subjected to greater study, including documentation of several assemblages from different areas of Iran ranging in age from Lower and Middle (Ernst et al. 2006, 2008, 2009a, b, 2010, 2011; Ernst and Gorgij 2013) to Upper Permian (Fantini Sestini 1965; Sakagami 1980).
Carboniferous bryozoans are abundant and diverse worldwide (Ross 1981; Bancroft 1987; Gorjunova et al. 2004), but are not well known from Iran, as mentioned above. In the present contribution, a bryozoan fauna from the Mississippian Mobarak Formation of Northern Iran is described. This fauna includes three species of the cystoporate Fistulipora M’Coy, 1849, and a species of the trepostome Hinaclema Sakagami and Sugimura, 1987. Previous studies reported the presence of brachiopods and corals as the main benthic organisms (Ogar et al. 2013; Qiao et al. 2016), but no bryozoans have been reported to date from this unit. Our study contributes to increasing knowledge of bryozoan diversity and palaeogeography in the Tethyan Gondwanan margin during the Mississippian.
Geological setting
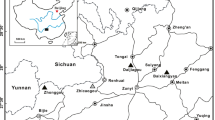
Iran is subdivided into several main tectono-sedimentary units (Fig. 1). Amongst them, the Alborz is over 2000-km long and extends from the Lesser Caucasus of Armenia and Azerbaijan republics in the northwest to the Paropamisus Mountains of northern Afghanistan in the east (Alavi 1996) (Fig. 1). Alborz is bounded to the north and northeast by the Palaeotethys collisional suture zone (Alavi 1996), to the northwest by the Caspian depression, and to the east by the Kopet-Dagh (Zonenshain and Pichon 1986) or Binaloud zone (Nogol-e-Sadat 1993). The southern and southeastern tectonic boundaries cannot be simply defined due to the different structures juxtaposed against the Alborz (Alavi 1996). The main tectonic event effective on the present shape of Alborz is thought to be the northward subduction of the Palaeotethys and subsequent collision between the Iranian Cimmerian microcontinent and the Turan plate, the southern part of Laurasia (Alavi 1991). The Alborz was subdivided into several subzones by researchers (Stöcklin 1974; Zanchi et al. 2009; Alavi 1996). Alavi (1996) divided Alborz basin into several tectono-stratigraphic units of the Upper Cambrian to Lower Ordovician epicontinental platform sequence, Middle Ordovician to Devonian magmatic rocks, Devonian to Middle Triassic continental shelf sequence Upper Triassic to Lower Jurassic foreland deposits, Middle Jurassic to Cretaceous epicontinental shelf succession, and Cenozoic synorogenic sediments. An alternative subdivision frequently used by researchers was presented by Stöcklin and Nabavi (1973) who divided northern Iran into three parts including Kopeh Dagh, Western Alborz, Azerbaijan, and Central Alborz. The latter part was further subdivided into south Central (Southern Alborz), north Central (Northern Alborz) and Anti-Alborz. During the Early Carboniferous, the Alborz Basin was an important palaeogeographical and bathymetric element along the northern margin of Gondwana (Vachard 1996; Torsvik and Cocks 2004, 2013; Muttoni et al. 2009). The Mississippian Mobarak Formation represents an extensive carbonate cycle along the northern margin of Gondwana deposited after the Palaeotethys rift opening (Vachard 1996; Mosaddegh 2000; Stampfli 2000; Lasemi 2001; Torsvik and Cocks 2004; Wendt et al. 2005; Golonka 2007).

Position of the Palaeozoic successions of the central and eastern Alborz basin of Iran. a Geographic location of the studied area in northern Iran. b Geological map of the study area, the central and eastern parts of the Alborz with location of the key section (modified after Aghanabati 2004). c Geological–structural map of studied Toyeh rodbar section. d Main roads accesses to the studied section
Mobarak Formation
The Toye rodbar section is located in Northern Iran and at the northern limit of the Alborz Mountains (Fig. 1). The Mobarak Formation in the Toye rodbar mountain area is 650-m thick and divided into three units based on the lithology (Fig. 2). The lower member of the Mobarak Formation consists mainly of dark grey thin- to medium-bedded shale to marly limestone with intercalated shale layers. The middle member consists of mainly thick- to medium-bedded limestone that is overlain by medium- to thin-bedded limestone alternating with a layer of ichnofossils, which is then overlain by marls. The upper member is characterised by dark marl and medium-bedded limestone followed by a medium-bedded limestone and medium layers of shale. The Mobarak Formation overlies the limestone of the Khosh yeilagh Formation (Upper Devonian age) and is overlain unconformably by sandstones of the younger Dorud Formation (Lower Permian age) in the Toye rodbar mountain section. The Mobarak Formation has been the focus of palaeontological research on corals and brachiopods (Ogar et al. 2013; Le et al. 2016), trace fossils and depositional environment (Nasiri et al. 2018, 2019), conodont biostratigraphy (Habibi et al. 2008), palynostratigraphy (Ghavidel-Syooki and Owens 2007), and foraminifers (Bozorgnia 1973; Mosaddegh 2000; Devuyst and Kalvoda 2007; Brenckle et al. 2009; Sardar Abadi et al. 2014). Based on these studies, the age of Mobarak Formation is defined as the Mississippian (Tournaisian-Viséan; Ogar et al. 2013; Qiao et al. 2016).

Stratigraphical section measured with sedimentological characteristics of the Mobarak Formation in the Toyeh rodbar section
The bryozoan material reported in this contribution comes from the upper interval (Viséan) of the Mobarak Formation in Northern Iran (Fig. 2).
Materials and methods
Bryozoans were investigated in thin sections using a binocular microscope in transmitted light. Morphologic character terminology is partly adopted from Anstey and Perry (1970). Thin sections are deposited at the Research Institute and Natural History Museum (Frankfurt am Main, Germany), under numbers SMF 23.756-SMF 23.839.
Systematic palaeontology
Phylum Bryozoa Ehrenberg, 1831
Class Stenolaemata Borg, 1926
Superorder Palaeostomata Ma, Buttler and Taylor, 2014
Order Cystoporata Astrova, 1964
Suborder Fistuliporina Astrova, 1964
Family Fistuliporidae Ulrich, 1882
Genus Fistulipora M‘Coy, 1849
Type species. Fistulipora minor M‘Coy, 1849; Carboniferous, England.
Diagnosis. Massive, encrusting or ramose colonies. Cylindrical autozooecia with thin walls and complete diaphragms. Apertures rounded, possessing horse-shoe-shaped lunaria. Autozooecia separated by the extrazooidal vesicular skeleton.
Remarks. Fistulipora M‘Coy, 1849 differs from Eridopora Ulrich, 1882 in having rounded, horseshoe-shaped lunaria instead of triangular ones. Furthermore, Eridopora occurs always as encrusting colonies, whereas Fistulipora may also develop massive and branched colonies.
Occurrence. Ordovician to Permian; worldwide.
Fistulipora longituba Yang and Lu, 1962

a–d Fistulipora longituba Yang and Lu, 1962. a Longitudinal section of a multilayered colony, SMF 23.773. b Longitudinal section showing autozooecial chambers and vesicles, SMF 23.778. c Tangential section showing autozooecial apertures, vesicles and a macula, SMF 23.777. d Tangential section showing autozooecial apertures and vesicles in intermacular area, SMF 23.777. e–g Fistulipora djebaglinica Nikiforova, 1933. e, f Longitudinal section showing autozooecial chambers and vesicles, SMF 23.819. g Tangential section showing autozooecial apertures and vesicles, SMF 23.825. Scale bars 5 mm (a, e), 2 mm (b, c), 1 mm (d, f, g)
1962 Fistulipora longituba Yang and Lu, p. 16, pl. 1, figs. 3–4, pl. 19, figs. 9–12.
1962 Fistulipora massiformis Yang and Lu, p. 16, pl. fig. 2, pl. 2, figs. 6–8.
1983 Fistulipora longituba Yang and Lu, 1962—Lu, p. 226, pl. 78, fig. 4, pl. 82, fig. 8.
1989 Fistulipora longituba Yang and Lu, 1962—Lu, p. 340, pl. 1, figs. 1–8, pl. 28, fig. 16.
Material. SMF 23.756–SMF 23.791.
Description. Massive multilayered colonies, 8–30 mm in thickness. Separate sheets 1.8–11.3 mm thick. Basal diaphragms common, thin, often deflected orally. Autozooecial apertures circular, slightly larger around maculae. Lunaria weakly developed, usually directed nearest macula positioned on the side closest to the macula. Vesicles medium in size, with rounded roofs, polygonal in tangential section, 9–15 surrounding each autozooecia aperture and separating the autozooecia in 1–3 rows. Autozooecial walls granular, 0.01–0.02 mm thick. Maculae flat, consisting of vesicles, 2.1–2.7 mm in diameter, spaced 9.0–11.4 mm from centre to centre.
Remarks. The studied material coincides with the species Fistulipora longituba Yang and Lu, 1962 from the Mississippian of Qilianshan (China). Lu (1989: 340) synonymised the species Fistulipora massiformis Yang and Lu, 1962 with F. longituba because of minor difference in the aperture size. Fistulipora longituba Yang and Lu, 1962 differs from F. djebaglinica Nikiforova, 1933 in having slightly larger autozooecial apertures (average aperture width 0.39 mm vs. 0.35 mm in F. djebaglinica), and in the more abundant vesicles (averagely 11.4 vesicles around autozooecial aperture vs. 8.8 vesicles in F. djebaglinica). Fistulipora longituba differs from F. prolifica Ulrich, 1884 from the Mississippian (Viséan) of the USA and France in having smaller distances between aperture centres (averagely 0.63 mm vs. 0.74 mm in F. prolifica), and smaller and more abundant vesicles.
Occurrence. Mississippian, Viséan; China. Mobarak Formation, Mississippian, Viséan; northern Iran.
Fistulipora djebaglinica Nikiforova, 1933
1933 Fistulipora djebaglinica Nikiforova, p. 5–6, pl. 1, figs. 4–5, text-figs. 1–3.
Material. SMF 23.792–SMF 23.825.
Description. Massive multilayered colonies, 6–16 mm in thickness. Separate sheets 1.4–8.1 mm thick. Basal diaphragms rare to common, thin, straight or slightly inclined. Autozooecial apertures circular to slightly oval. Lunaria well developed, prominent, directed the nearest macula positioned on the side closest to the macula. Vesicles large, 6–12 surrounding each autozooecial aperture, with flattened roofs, polygonal in tangential section, separating the autozooecia in 1–3 rows. Autozooecial walls granular, 0.01–0.02 mm thick. Maculae deeply depressed, consisting of vesicles, 1.6–2.9 mm in diameter, spaced 10–12 mm from centre to centre.
Remarks. The original description of the species Fistulipora djebaglinica Nikiforova, 1933 does not contain information about the aperture size. The spacing of apertures was given as 5–6 in 3 mm distance, which corresponds the distance between aperture centres of 0.5–0.6 mm and falls in the range of the present material (0.51–0.88 mm). The size of the maculae (ca. 2 mm vs. 1.6–2.9 mm in the present material) and their spacing are very close (7.0–7.5 mm vs. 10–12 mm in the present material). Fistulipora djebaglinica Nikiforova, 1933 differs from F. longituba Yang and Lu, 1962 in having smaller autozooecial apertures and less abundant vesicles. Fistulipora djebaglinica differs from F. tubulosa Nikiforova, 1933 in having a massive instead of hollow ramose colony and in having larger and less abundant vesicles (6–12 vesicles per autozooecial aperture vs. 15–17 in F. tubulosa). Fistulipora djebaglinica differs from F. prolifica Ulrich, 1884 in having smaller autozooecial apertures (0.35 mm vs. 0.39 mm in F. prolifica) as well as in smaller distances between autozooecial apertures (average 0.64 mm vs. 0.74 mm in F. prolifica).
Occurrence. Mississippian; Kazakhstan. Mobarak Formation, Mississippian, Viséan; northern Iran.
Fistulipora sp.

a–c Fistulipora sp. a longitudinal section showing autozooecial chambers and vesicles, SMF 23.826. b tangential section showing autozooecial apertures and vesicles, SMF 23.827. c tangential section showing autozooecial aperture with lunarium and vesicles, SMF 23.827. d–g Hinaclema sakagamii Schastlivtseva, 1991. d transverse section of a multilayered colony, SMF 23.835. e longitudinal section showing autozooecial chambers and exilazooecia, SMF 23.836. f transverse section showing exilazooecia and an acanthostyle, SMF 23.835. g tangential section showing autozooecial apertures, exilazooecia and acanthostyles, SMF 23.837. Scale bars 2 mm (d), 1 mm (a, b, e, g), 0.5 mm (c, f)
1989 Fistulipora cf. torosa Moore and Dudley, 1944—Lu, p. 340, pl. 1, figs. 9–12, pl. 2, fig. 21, pl. 29, fig. 2.
Material. Seven thin sections of a single colony SMF 23.826–SMF 23.832.
Description. Encrusting, submassive colony, 10–13 mm thick. Basal diaphragms common to abundant, thin, straight or slightly deflected. Autozooecial apertures circular to slightly oval. Macrozooecia occurring locally, 0.54–0.64 mm in width. Lunaria weakly developed, wide and thin. Vesicles large, 7–13 surrounding each autozooecial aperture, with flattened roofs, polygonal in tangential section, separating the autozooecia in 1–2 rows. Autozooecial walls granular, 0.010–0.015 mm thick. Maculae not observed.
Remarks. The present species is similar to the Fistulipora cf. torosa Moore and Dudley, 1944 of Lu (1989). It differs from F. longituba Yang and Lu, 1962 and F. djebaglinica Nikiforova, 1933 in larger and more widely separated autozooecial apertures. It differs from F. prolifica Ulrich, 1884 in having larger autozooecial apertures (average aperture width 0.43 mm vs. 0.39 mm in F. prolifica).
Occurrence. Mississippian, Viséan; China. Mobarak Formation, Mississippian, Viséan; northern Iran.
Order Trepostomata Ulrich, 1882
Family Crustoporidae Dunaeva and Morozova, 1967
Genus Hinaclema Sakagami and Sugimura, 1987
Type species. Hinaclema hinaensis Sakagami and Sugimura, 1987; Mississippian, Viséan, Japan.
Diagnosis. Colony lamellar, encrusting with secondary overgrowths. Autozooecia cylindrical, recumbent at their bases, bending sharply in exozones, intersecting the colony surface at right angles. Basal diaphragms in autozooecia rare to common. Exilazooecia abundant, separating autozooecia in multiple rows. Acanthostyles abundant throughout the colony, usually large, originating at the base of exozone. Autozooecial walls finely laminated, merged, without visible boundaries.
Remarks. Hinaclema Sakagami and Sugimura, 1987 differs from Crustopora Dunaeva, 1964 by having large acanthostyles, instead of small needle-like ones, and abundant exilazooecia. Toulopora Nakrem, Błażejowski, and Gaździcki, 2009 differs in having of acanthostyles of two sizes: sparsely distributed large ones, and a row of regularly distributed small ones inflecting each zooecial aperture.
Occurrence. Mississippian, Viséan; Japan, Uzbekistan. Mississippian, Serpukhovian (Pendleian); La Cornuda, southwestern Spain. Mobarak Formation, Mississippian, Viséan; northern Iran.
Hinaclema sakagamii Schastlivtseva, 1991

a, b Hinaclema sakagamii Schastlivtseva, 1991. a Transverse section showing exilazooecia and an acanthostyle, SMF 23.834. b Tangential section showing autozooecial apertures, exilazooecia and acanthostyles, SMF 23.837. Scale bars 0.2 mm
1991 Hinaclema sakagamii Schastlivtseva, p. 115, fig. 1.
Material. SMF 23.833–SMF 23.839.
Description. Colony lamellar, encrusting, 0.9–2.2 mm thick. Cylindrical autozooecia recumbent at their bases, bending sharply in exozones and intersecting the colony surface at right angles. Basal diaphragms common, 1–3 in each autozooecium. Autozooecial apertures rounded to oval. Exilazooecia abundant, 1–4 rows separating autozooecia, rounded-polygonal in tangential section, originating at the base of the exozone, 20–30 surrounding each autozooecial aperture. Acanthostyles abundant throughout the colony, originating at the base of the exozone, 1–3 surrounding each autozooecial aperture, having narrow hyaline cores and wide laminated sheaths. Autozooecial walls finely laminated, merged, without visible boundaries and monilae-shaped thickenings, 0.015–0.020 mm thick in endozone, and 0.025–0.035 mm thick in exozone.
Remarks. Hinaclema sakagamii Schastlivtseva, 1991 differs from H. hinaensis Sakagami and Sugimura, 1987 from the Mississippian (Viséan) of Japan and Spain in wider autozooecia (0.36–0.50 mm vs. 0.18–0.28 mm in H. hinaensis), and in more abundant exilazooecia (20–30 exilazooecia around each autozooecial aperture vs. 11–16 in H. hinaensis).
Occurrence. Mississippian, Viséan; Uzbekistan. Mobarak Formation, Mississippian, Viséan; northern Iran.
Palaeobiogeography
A general palaeobiogeographical analysis of the Carboniferous bryozoans has been provided by Ross and Ross (1985), with a summary of Late Palaeozoic (Carboniferous–Permian) bryozoan distribution. A more recent study focusing on the Mississippian bryozoans from Eurasia was that by Tolokonnikova et al. (2014), and a particular analysis of Mississippian bryozoans from Iran by Tolokonnikova and Yazdi-Moghadam (2013) who described the bryozoan fauna from the Shishtu Formation of central Iran. In these contributions, it was established that the general palaeogeographical and environmental setting of the Mississippian units of northern Iran represents a carbonate ramp platform deposited on the passive margin of the southern Palaeotethys Ocean. This extensive carbonate cycle developed along the northern margin of Gondwana deposited after the Palaeotethys rift opening.
Tolokonnikova and Yazdi-Moghadam (2013) recognised four bryozoans in the Shishtu Formation of central Iran: Nikiforovella ulbensis Nekhoroshev, 1956, Nicklesopora elegantulaformis (Nekhoroshev, 1956), Primorella cf. iranica Gorjunova, 2006, and Nikiforopora intermedia (Nikiforova, 1950). Three other species were described by Gorjunova (2006) from Viséan rocks of the Eastern Elbrus in the north of Iran: Heloclema magnificum Gorjunova, 2006, Worthenopora elbursensis Gorjunova, 2006, and Primorella iranica Gorjunova, 2006. These species were considered as endemics in the Mississippian.
The bryozoan fauna of the Mobarak Formation seems to be abundant and diverse. In this publication, a collection of large massive bryozoans with their encrusters was studied. Massive bryozoans (mostly multilayered) are represented by three species of the cystoporate genus Fistulipora M‘Coy, 1849. Two colonies contained additionally an encrusting trepostome bryozoan Hinaclema sakagamii Schastlivtseva, 1991. The study of the matrix surrounding some colonies of Fistulipora revealed fine debris of fenestrate, cystoporate, and cryptostome bryozoans, which could not be identified to a genus level.
Genus Fistulipora M‘Coy, 1849 is known from the Ordovician to Permian, and contains up to 200 species. This genus is probably polyphyletic and needs a comprehensive revision. Fistulipora was widely distributed in the Carboniferous and Permian marine basins. Three species of Fistulipora occur in the Mobarak Formation: Fistulipora longituba Yang and Lu, 1962, F. djebaglinica Nikiforova, 1933, and F. sp. Both F. longituba and F. djebaglinica have previous records in China and Kazakhstan, respectively. The species Fistulipora sp. is apparently con-specific with the bryozoan described as Fistulipora cf. torosa Moore and Dudley, 1944 by Lu (1989) from the Viséan of western China. This species is not closely related to Cyclotrypa torosa Moore and Dudley, 1944 from the Pennsylvanian of USA. However, as only one specimen is available for the study, it was not possible to establish a new species for the Iranian material.
Hinaclema was previously recorded in Japan, Uzbekistan, and southwestern Spain, while H. sakagamii Schastlivtseva, 1991, reported in this contribution, was previously recorded from Uzbekistan. This genus is known within a very narrow time interval restricted mainly to the Viséan. The species H. hinaensis Sakagami and Sugimura, 1987 has been also identified from the Serpukhovian (Pendleian) of southwestern Spain (Ernst and Rodríguez 2013).
In the exhaustive analyses of Tolokonnikova and Yazdi-Moghadam (2013) and Tolokonnikova et al. (2014), the Viséan bryozoan assemblage from central Iran shows close palaeobiogeographical affinities with Kazakhstan and Russia (eastern Transbaikalia, Kurgan region), and these authors also revealed connections between Iran and some regions of Laurussia during the Early Carboniferous (shelf of Kazakhstan and Siberian terrains). Some connections with Spain and Mongolia were also demonstrated.
The addition of our data from the Mississippian of Northern Iran reinforces the palaeogeographical affinities with Uzbekistan, Kazakhstan and China that were grouped as very closely associated by Tolokonnikova and Yazdi-Moghadam (2013). The previous analyses on the Carboniferous bryozoan fauna suggest connections to both the western and eastern part of the northern Palaeotethyan Realm. We can state here that the Mississippian bryozoans of Iran are closer to the Eastern centre of radiation (see Tolokonnikova et al. 2014) that includes Kazakhstan, Mongolia, the Kuznetsk Basin, Eastern Transbaikalia, and Uzbekistan.
References
Alavi, M. 1991. Tectonic map of the Middle East, scale: 1:5,000,000. Tehran: Geological Survey of Iran.
Alavi, M. 1996. Tectonostratigraphic synthesis and structural style of the Alborz Mountain system in Iran. Journal of Geodynamics 21 (1): 1–33.
Anstey, R.L., and T.G. Perry. 1970. Biometric procedures in taxonomic studies of Paleozoic bryozoans. Journal of Paleontology 44(3): 383–398.
Astrova, G.G. 1964. A new order of the Paleozoic Bryozoa. Paleontologicheskiy Zhurnal 1964(2): 22–31. (in Russian).
Bancroft, A.J. 1987. Biostratigraphical potential of Carboniferous Bryozoa. Courier Forschungsinstitut Senkenberg 98: 193–197.
Borg, F. 1926. Studies on Recent cyclostomatous Bryozoa. Zoologiska Bidrag från Uppsala 10: 181–507.
Bozorgnia, F. 1973. Paleozoic foraminiferal biostratigraphy of central and east Alborz Mountains, Iran. Publication of National Iranian Oil Company (Geological Laboratory) 4: 1–185.
Devuyst, F.-X., and J. Kalvoda. 2007. Early evolution of the genus Eoparastafella (Foraminifera) in Eurasia: the “interiecta group” and related forms, late Tournasisan to early Viséan (Mississippian). Journal of Foraminiferal Research 37(1): 69–89.
Dunaeva, N.N., and I.P. Morozova. 1967. Osobennosti razvitiya i sistematicheskoe polozhenie nekotorykh pozdnepaleozojskikh trepostomat. Paleontologicheskiy Zhurnal 1967(4): 86–94. (in Russian). [English translation: Evolution and systematic position of some Paleozoic Trepostomata. Paleontological Journal 1(4): 64–72].
Ehrenberg, C.G. 1831. Symbolae Physicae, seu Icones et descptiones Corporum Naturalium novorum aut minus cognitorum, quae ex itineribus per Libyam, Aegiptum, Nubiam, Dongalaam, Syriam, Arabiam et Habessiniam, studia annis 1820–25, redirent. Pars Zoologica, 4, Animalia Evertebrata exclusis Insectis. Berolini.
Ernst, A., and M. Mohammadi. 2009. Stenolaemate bryozoans from the Geirud Formation (upper Devonian/lower Carboniferous) of Central Alborz (Iran). Paläontologische Zeitschrift 83: 439–447.
Ernst, A., and M.N. Gorgij. 2013. Lower Permian bryozoan faunas from Kalmard area, central Iran. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 268(3): 275–324.
Ernst, A., and S. Rodríguez. 2013. Stenolaemate bryozoan fauna from the Mississippian of Guadiato area, southwestern Spain. Spanish Journal of Palaeontology 28(2): 173–192.
Ernst, A., B. Senowbari-Daryan, and K. Rashidi. 2006. Lower Permian Bryozoa of the Jamal Formation from Bagh-e Vang (Shotori Mountains, northeast Iran). Facies 52(1): 627–635.
Ernst, A., B. Senowbari-Daryan, and K. Rashidi. 2008. Permian Bryozoa from the Jamal Formation of Shotori Mountains (northeast Iran). Revue de Paléobiologie 27(2): 395–408.
Ernst, A., B. Senowbari-Daryan, and K. Rashidi. 2009a. Bryozoa from the Surmaq Formation (Permian) of Hambast Mountains, south of Abadeh, central Iran. Facies 55(4): 595–608.
Ernst, A., B. Senowbari-Daryan, and K. Rashidi. 2009b. Rhabdomesid and cystoporid bryozoans from the Permian of Deh-e Mohammad, Shotori Mountains (northeastern Iran). Geobios 42(2): 133–140.
Ernst, A., H. Yarahmadzahi, and M.N. Gorgij. 2010. Sakmarian Bryozoa from the Dalgan area (Sarab section), southeastern Iran. Freiberger Forschungshefte (C: Paläontologie, Stratigraphie, Fazies) 536: 75–89.
Ernst, A., T. Parvizi, and K. Rashidi. 2011. Some Bryozoa from the Upper Permian Dalan Formation of Dena Mountain in SW Iran. Freiberger Forschungshefte (C: Paläontologie, Stratigraphie, Fazies) 539: 71–81.
Ernst, A., Z. Tolokonnikova, and H. Yarahmadzahi. 2012. Upper Devonian (Frasnian) Bryozoa from the Shishtu 1 Formation of Niaz area (northeast Iran). Revue de Paléobiologie 31(1): 1–14.
Ernst, A., P. Königshof, A. Bahrami, M. Yazdi, and I. Boncheva. 2017. A Late Devonian (Frasnian) bryozoan fauna from the Central Iran. Palaeobiodiversity and Palaeoenvironments 97: 541–552.
Fantini Sestini, F.N. 1965. The geology of the upper Djadjerud and Lar valleys (north-Iran); II, Palaeontology; bryozoans, brachiopods and molluscs from Ruteh limestone (Permian). Rivista Italiana di Paleontologia 71(1): 13–108.
Ghavidel-syooki, M., and B. Owens. 2007. Palynostratigraphy and palaeogeography of the Padeha, Khoshyeilagh and Mobarak formations in the eastern Alborz Range (Kopeh-Dagh region), northeastern Iran. Revue de Micropaleontology 50(1): 129–144.
Golonka, J. 2007. Phanerozoic palaeoenviroment and paleolithofacies maps. Geologia 33: 145–209.
Gorjunova, R.V. 2006. New bryozoans from the Devonian of Afghanistan and the Carboniferous of Iran. Paleontological Journal 40: 626–635.
Gorjunova, R.V., A.B. Markov, and E.B. Naimark. 2004. Evolution and biogeography of the Palaeozoic Bryozoa: results of the numerical analysis, 1–182. Moscow: GEOS.
Habibi, T., C. Corradini, and M. Yazdi. 2008. Conodont biostratigraphy of the upper Devonian-Lower Carboniferous Shahmirzad section, central Alborz, Iran. Geobios 41(6): 763–777.
Hairapetian, V., M.G. Pour, L.E. Popov, P. Männik, and C.G. Miller. 2017. Silurian stratigraphy of Central Iran—an update. Acta Geologica Polonica 67(2): 201–233.
Lasemi, Y. 2001. Facies analysis, depositional environments, and sequence stratigraphy of the Upper Pre-Cambrian and Paleozoic rocks of Iran, 1–180. Tehran: Geological Survey of Iran Press. (in Persian).
Qiao, L., M. Falahatgar, and S.Z. Shen. 2016. A lower Viséan (Carboniferous) brachiopod fauna from the eastern Alborz Mountains, northern Iran, and its palaeobiogeographical implications. Geological Journal 52: 317–326.
Lu, L. 1983. Phylum Bryozoa. In Paleontological Atlas of northwest China. Shaanxi-Gansu-Ningxia Volume. Part 2. Upper Paleozoic, 221–244. Beijing: Geological Publishing House. (in Chinese).
Lu, L. 1989. Bryozoans from Chouniugou Formation of Late Early Carboniferous on the Northern Slope of Mt. Qilian. Palaeontologia Gathayana 4: 327–433.
Ma, J.-Y., C.J. Buttler, and P.D. Taylor. 2014. Cladistic analysis of the ‘trepostome’ Suborder Esthonioporina and the systematics of Palaeozoic bryozoans. In Bryozoan Studies 2013, eds. A. Rosso, P.N. Wyse Jackson, and J.S. Porter. Studi Trentini di Scienze Naturali 94: 153–161.
M‘Coy, F. 1849. On some new genera and species of Palaeozoic corals and foraminifera. Annals and Magazine of Natural History 3 (2): 119–136.
Mistiaen, B., H. Gholamalian, R. Gourvennec, Y. Plusquellec, F. Bigey, D. Brice, M. Feist, R. Feist, P. Ghobadi, and E.-M. Kebria. 2001. Preliminary data on the Upper Devonian (Frasnian, Famennian) and Permian fauna and flora from the Chahriseh area (Esfahan Province, central Iran) (In New systematic and palaeobiogeographic data from Palaeozoic of central Iran, eds. R. Feist, and J.-A. Talent). Annales de la Societe Geologique du Nord 8(2): 93–102.
Moore, R.C., and R.M. Dudley. 1944. Cheilotrypid bryozoans from Pennsylvanian and Permian rocks of the mid-continent region. Bulletin of Kansas Geological Survey 52(6): 229–408.
Mosaddegh, H. 2000. Microfossils, Microfacies, Sedimentary Environment and Sequence Stratigraphy of the Mobarak Formation in Central Alborz. Unpublished Ph.D. thesis, 1–269. Tehran: Tehran Teacher Training University.
Muttoni, G., M. Mattei, M. Balini, A. Zanchi, M. Gaetani, and F. Berra. 2009. The drift history of Iran from the Ordovician to the Triassic. In South Caspian to Central Iran Basins, eds. M.-F. Brunet, M. Wilmsen, and J.W. Granath. Geological Society of London, Special Publications 312: 7–29.
Nakrem, H.A., B. Błażejowski, and A. Gaździcki. 2009. Lower Permian bryozoans from southern and central Spitsbergen, Svalbard. Acta Palaeontologica Polonica 54(4): 677–698.
Nasiri, Y., R. Moussavi-Harami, A. Mahboubi, and H. Mosaddegh. 2019. Sequence stratigraphic significance of shell concentrations in the Mobarak Formation (Mississippian), Alborz Zone, Northern Iran. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen 291(2): 171–195.
Nasiri, Y., R. Moussavi-Harami, A. Mahboubi, D. Olivero, and H. Mosaddegh. 2018. Zoophycos ichnogenus distribution and paleoenvironmental analysis: examples from the Mississippian Mobarak formation (Alborz Basin, Iran). Historical Biology. https://doi.org/10.1080/08912963.2018.1540614.
Nikiforova, A.I. 1933. Kamennougol’nye otlozhenii’a stredneĭ Azii [The Carboniferous deposits of Central Asia. Contributions to the knowledge of the Lower Carboniferous Bryozoa of the Turkestan]. Trudy Vsesoiuznogo Geologo-Razvedochnogo Ob”edineniia NKTP SSSR [Transactions of the United Geological and Prospecting Service of USSR] 207: 1–78. (in Russian).
Nogol-e-Sadat, M. 1993. 1:1,000,000 Tectonic maps of Iran. Tehran: Department of Iranian Geological Center.
Ogar, V., F. Falahatgar, and M. Hossein. 2013. Visean corals of the Kiyasar area, south of Sari, Northern Iran. Revista Brasilera Paleontologia 16(3): 375–396.
Ross, C.A., and J.R.P. Ross. 1985. Carboniferous and early Permian biogeography. Geology 13: 27–30.
Ross, J.R.P. 1981. Biogeography of Carboniferous ectoproct Bryozoa. Palaeontology 24(2): 313–341.
Ross, J.R.P., M.A. Hamedi, and A.J. Wright, 2000. Late Ordovician (Caradoc) bryozoans from the Kerman district, east-central Iran. In Proceedings of the 11th International Bryozoology Association Conference, eds. C.A. Herrera, and J.B.C. Jackson, 329–335. Balboa, R.P.: Smithsonian Tropical Research Institute.
Sakagami, S. 1980. Permian Ectoprocta (Bryozoa) from the Abadeh Region, Central Iran. Transactions and Proceedings of the Palaeontological Society of Japan, New Series 118: 269–289.
Sakagami, S., and A. Sugimura. 1987. Hinaclema, a new Carboniferous bryozoan genus from the Hina Limestone, southwest Japan. Proceedings Japan Academy B (Physical and Biological Sciences) 63: 246–249.
Sardar Abadi, M., E. Kulagina, A.-C. Da Silva, H. Mossadegh, P. Steemans, and F. Boulvain. 2014. Depositional facies and regional paleogeography of the Lower Carboniferous Mobarak formation in the Alborz Basin, Northern Iran. In Central European Meeting of Sedimentary Geology: an international conference: Olomouc, Czech Republic, June 9–13, 2014, Abstracts, eds. O. Bábek, T.M. Grygar, and D. Uličný, 84–86. Olomouc: Palacký University.
Schastlivtseva, N.P. 1991. Pervaya nachodka v SSSR kamennougolnogo roda Hinaclema (mshanki) [The first find of the Carboniferous genus Hinaclema (bryozoans) in the USSR]. Paleontologicheskii Zhurnal 1991(1): 118–120. (in Russian).
Stampfli, G.M. 2000. Tethyan oceans. In Tectonics and magmatism in Turkey and surrounding area, eds. E. Bozkurt, J.A. Winchester, and J.D.A. Piper. Geological Society of London, Special Publication 173: 63–185.
Stöcklin, J. 1974. Possible ancient continental margin in Iran. In The geology of continental margins, eds. C.A. Burke and C.L. Drake, 873–887. New York: Springer.
Stöcklin, J., and N.H. Nabavi. 1973. Tectonic map of Iran (1:2,500,000). Tehran: Ministry of Mines and Metals, Geological Survey of Iran.
Tolokonnikova, Z., and M. Yazdi-Moghadam. 2013. Early Viséan bryozoans from the Shishtu II member, Shishtu Formation, central Iran. Geologos 19(4): 291–299.
Tolokonnikova, Z., A. Ernst, and P.N. Wyse Jackson. 2014. Palaeobiogeography and diversification of Tournaisian-Viséan bryozoans (lower–middle Mississippian, Carboniferous) from Eurasia. Palaeogeography, Palaeoclimatology, Palaeoecology 414: 200–211.
Torsvik, T.H., and L.R.M. Cocks. 2004. Earth geography from 400 to 250 Ma: a palaeomagnetic, faunal and facies review. Journal of the Geological Society 161: 555–572.
Torsvik, T.H., and L.R.M. Cocks. 2013. Gondwana from top to base in space and time. Gondwana Research 24: 999–1030.
Ulrich, E.O. 1882. American Palaeozoic Bryozoa. The Journal of the Cincinnati Society of Natural History 5: 121–175, 233–257.
Ulrich, E.O. 1884. American Palaeozoic Bryozoa. The Journal of the Cincinnati Society of Natural History 8: 24–51.
Vachard, D. 1996. Iran. In Carboniferous of the World 3, eds. R.H. Wagner, C.F. Winkler Prins, and L.F. Granados, 491–513. Madrid: Instituto Geológico y Minero de España.
Wendt, J., B. Kaufmann, Z. Belka, N. Farsan, and A.K. Bavandpour. 2005. Devonian/lower Carboniferous stratigraphy, facies patterns and palaeogeography of Iran. Part II. North and central Iran. Acta Geologica Polonica 55: 31–97.
Yang, J., and L. Lu. 1962. Palaeozoic bryozoans of Qilianshan. Geology of Qilianshan 4: 1–114. (in Chinese).
Zanchi, A., S. Zanchetta, E. Garzanti, M. Balini, F. Berra, M. Mattei, and G. Muttoni. 2009. The Cimmerian evolution of the Nakhlak–Anarak area, central Iran, and its bearing for the reconstruction of the history of the Eurasian margin. In South Caspian to Central Iran Basins, eds. M.-F. Brunet, M. Wilmsen, and J.W. Granath. Geological Society of London, Special Publication 312: 261–286.
Zonenshain, L.P., and X. Pichon. 1986. Deep basins of the Black Sea and Caspian Sea as remnants of Mesozoic back-arc basins. Tectonophysics 123(1–4): 181–211.
Acknowledgements
We acknowledge the Department of Geology, Faculty of Science, Ferdowsi University of Mashhad, Iran for financial support and field expenses through Grant no. 3/43573. AE thanks Deutsche Forschungsgemeinschaft (DFG) for financial support (project ER 278/6.1). MGC acknowledges support from Research Council of Argentina (CONICET). Patrick Wyse Jackson, Dublin, and Paul D. Taylor, London, are thanked for their constructive and helpful reviews.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Mike Reich.
Rights and permissions
About this article

Cite this article
Ernst, A., Nasiri, Y., Carrera, M.G. et al. Bryozoan fauna from the Mobarak Formation (Mississippian), northern Iran. PalZ 95, 105–116 (2021). https://doi.org/10.1007/s12542-019-00504-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12542-019-00504-8