
Abstract
The incertae sedis Carpathoporella Dragastan, 1995, reported from the Lower Cretaceous of the Western Tethyan domain, is usually interpreted as remains of calcareous algae (Dasycladales or Characeae). New thin-section material from the Aptian of Albania sheds light not only on its biogenic nature but also on the morphological variability of this taxon. In fact, Carpathoporella represents the debris of colonial, bushy, most likely gorgonid octocorals with tuberculated spheroids that may be fused at least near the basal root-like holdfast. Colony branching originates from longitudinally grooved calcareous branches or internodes. Possible relationships to other Upper Cretaceous to Palaeogene genera are discussed and a revised critical inventory of Cretaceous octocorals is presented. Due to the evidenced morphological features, Carpathoporella could either represent an ancestral isidid octocoral of the order Alcyonacea such as Moltkia Steenstrup or, due to the likely primary aragonitic skeletal mineralogy, a representative of Epiphaxum Lonsdale of the order Helioporacea. Due to morphological analogies, the new combination Carpathoporella elliotti (Radoičić) is proposed. In any case, the Lower Cretaceous record from Tethyan peri-reefal shallow-water carbonates is highlighted since numerous skeletal findings of fossil gorgonid Octocorallia were so far only known from Upper Cretaceous and younger strata of outer shelf environments of the boreal realm. The origin of deep-water Upper Cretaceous octocorals from Lower Cretaceous shallow-water taxa such as Carpathoporella is proposed as a possible further example of onshore/offshore evolutionary pattern.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Microproblematica (or incertae sedis) are microorganisms, sometimes only parts of them, that are, due to the poverty of characteristic features, or due to contradictory characteristics, not compatible with existing taxonomic concepts. Therefore, these fossils cannot be assigned with certainty to an organism or a group of organisms (e.g., Schmid 1996). Small, variously star-shaped or elongated calcitic bodies with or without a central hollow and assigned to the enigmatic incertae sedis Coptocampylodon Elliott and Carpathoporella Dragastan are widespread in Upper Jurassic to Upper Cretaceous shallow-marine carbonates of the Tethyan realm (e.g., Elliott 1963; Dragastan 1967, 1989; Radoičić 1969; Basson and Edgell 1971; Schlagintweit et al. 2002). Since the establishment of both taxa, their validity, possible synonymies, and biogenic nature has long been disputed with contradictory opinions. Coptocampylodon was interpreted by Elliott (1963) as calcitic remains of octocorals (“soft corals”), later as anomuran coprolites or algal remains (see micropalaeontological part for details). New material from Lower Cretaceous carbonates on top of the Mirdita ophiolites in Albania sheds light not only on the systematic position but also on the intraspecific morphological variability of a taxon invalidy described for the first time by Patrulius (1966) as Coptocampylodon fontis, later typified as Carpathoporella occidentalis Dragastan, 1995, non 1967, non 1989. It will be shown in the present paper that this taxon cannot be ascribed to debris of calcareous algae but can be referred to different skeletal parts of fossil octocorals. Noteworthy, also for Pienina oblonga Borza and Misík, 1976, another microproblematicum that can occur in Lower Cretaceous shallow-water limestones, a possible octocoral origin is discussed (Granier 1988; Mišík 1998).
Geological setting
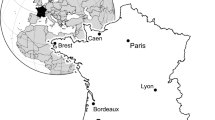
In the Mirdita Zone of central Albania, large areas on top of the radiolaritic-ophiolitic trench fills (=mélange) are covered by Neocomian shallow-water carbonates (Xhomo et al. 2002), e.g., the Munella and Mali i Shejtit carbonate platforms (MCP, MSP) (see Gawlick et al. 2008 and Schlagintweit et al. 2008 for details) (Fig. 1). At the so-called Gezolla section, representing the southernmost part of the MCP (Fig. 2), clasts of (Upper) Berriasian-Valanginian shallow-water carbonates with abundant benthic foraminifera, namely Andersenolina campanella (Fig. 3a) and Pseudocyclammina lituus (Yokoyama) (Fig. 3b), are reworked within basal breccias associated with coal series. The following reddish to grey limestones above the breccias are devoid of biostratigraphic useful microfossils. Dasycladalean algae of the Nikšić type assemblage (Sokač and Nikler 1973; Schindler and Conrad 1994) such as Suppiluliumaella elliotti Bakalova (? = Crinella carsica Sokač and Nikler) (Fig. 3c) or Montenegrella div. sp. (Fig. 3e) occur in the lower and middle part of the section (samples Al 832, 836, 839, 840, 842 and 844; see Fig. 2b). It is not the intention here to document further on this group of dasycladales needing taxonomic reconsideration (e.g., Barattolo 1984; Radoičić 2002). The occurrence of these dasycladales within samples containing extraclasts and iron-stained matrix compares to the occurrence in the Nišić area of Montenegro where they are reported above reddish karstic bauxites also with “Coptocampylodon fontis” (Sokač and Nikler 1971, 1973). The occurrence of the dasycladalean alga Montiella ellitzae (Bakalova) (Fig. 3d) in sample Al 848 roughly accounts for a Barremian-Aptian age at least for the topmost parts (e.g., Bucur 1999). From the highest part of the Gezolla section, Peza and Marku (2002) reported Cylindroporella sudgeni Elliott, Carpathoporella fontis (Patrulis) and Paleodictyoconus arabicus Henson (=Montseciella arabica acc. to Cherchi and Schroeder 1999 and Schroeder et al. 2002) pointing to an Aptian age of the upper part of the profile. Therefore these shallow-water carbonates belong to the MSP. Summarizing, the Gezolla section represents another transgressive sedimentary cycle following the Neocomian Munella platform starting in the Upper Hauterivian or Lower Barremian and named as the Mali I Shejtit platform (Gawlick et al. 2008) as documented by the occurrence of the benthic foraminifer Campanellula capuensis De Castro (e.g., Velić 2007) at another sampled section.

Geological map of central Mirdita Zone in Albania with shallow-water carbonate platforms on top of the ophiolites resp. the radiolaritic-ophiolitic trench fills (mélange)—the Mali i Sheijtit and Munella Mountains (Mali i Munelles). Arrow shows locality of Gezolla section. Map simplified on base of Harta Gjeologjike e Shqiperise 1:200,000 (Xhomo et al. 2002), including own results (Gawlick et al. 2008)

a Simplified tectonic situation at the southern rim of Munella platform characterized by high-angle normal faults and position of Gezolla section. b Gezolla section and position of the sample AL 847 containing the described material

Microfossils from the reddish, breccia-rich limestones from reworked pebbles of the Lower Cretaceous Munella Carbonate Platform at the base of Gezolla section, Mirdita Zone of Albania, Munella Mountains. a Benthic foraminifer Andersenolina campanella (Arnaud-Vanneau, Boisseau and Darsac). Sample Al 837. b Benthic foraminifer Pseudocyclammina lituus (Yokoyama). Sample Al 838. c Dasycladale Suppiluliumaella elliotti Bakalova (? = Crinella carsica Nikler and Sokac). Sample Al 840. d Dasycladale Montiella ellitzae (Bakalova). Sample Al 848. e Dasycladales Suppiluliumaella elliotti Bakalova (left) and large Suppiluliumaella? sp. (right). Note Bacinella irregularis Radoičić filling the main axis. f Packstone with lithoclasts and iron-stained matrix (right); left a transverse section of the dasycladale Dissocladella sp. or Suppiluliumaella sp. Sample Al 843. g Packstone with abundant remains of Carpathoporella occidentalis Dragastan; white rectangle marks the detail shown in h. Sample Al 847. h Detail from g. Sample Al 847a. For a–c, e, f scale bar is 1 mm; for d, h scale bar is 0.5 mm, for g scale bar is 2 mm
Fossil versus recent octocorals: a brief overview with special reference to Cretaceous taxa
Octocorals, frequently also referred to as soft corals, are a subclass of sessile Anthozoa, being mostly colonial with forms ranging from encrusting sheets to complex branching forms (e.g., Bayer 1956; Bayer et al. 1983). As the name suggests, it refers to their polyps having eight tentacles. Many (but not all) octocorals can be classified as cold-water or deep-water corals. Recent forms range in size from only a few centimeters up to a few meters and occupy habitats ranging from the intertidal zone down to abyssal depths and a geographical distribution from pole to pole (e.g., Bayer 1961). Some octocoral families are characteristic for warm shallow-water areas where they are conspicuous components of reef communities, especially in the tropical Indo-Pacific realm (e.g., Bayer 1961; Fabricius and Alderslade 2001; Grasshoff and Bargibant 2001), others are almost exclusively inhabitants of cold deep-water habitats (e.g., Wing and Barnard 2004). Recently, representatives of the genus Narella Gray have been recorded from a depth of 4,594 m in the North-Pacific realm (Cairns and Baco 2007). With respect to fossil counterparts, for example, depths of 100–300 m were estimated by Bernecker and Weidlich (2006) for the well-known Danian coral mounds of Fakse, Denmark. Especially deep-sea octocorals are slow growing, possessing a longevity potential of up to 200 years (Andrews et al. 2002, 2005).
In the “Treatise of invertebrate palaeontology” of 1956, Bayer mentions 45 living families of which 13 are known from the fossil record. From the Upper Cretaceous onwards, six families are recorded (Bayer 1956; Voigt 1958). At present, approximately 340 genera of octocorals are known (Williams and Cairns 2006) including over 3,000 extant species (Daly et al. 2007). For the approximate numbers of genera and species within each of the extant octocoral families, see Daly et al. (2007). Octocorals belong to a group of animals, such as also Holothuroidea, that are composed of multi-element skeletons disarticulating rather quickly post-mortem. Findings of fossil octocoral remains are therefore generally rare with occasional records in the Palaeozoic (e.g., Lindström 1978; Bengtson 1981; Reich 2001, 2002; Cope 2005). The oldest record might be from the Cambrian Burgess Shale (Ausich and Babcock 1998, 2000). Generally, there is a discontinuous record, and the relationship of the Palaeozoic spicules ascribed to the order Alcyonacea to Mesozoic or younger findings, is obscure. Octocorals are well recorded from Upper Cretaceous (e.g., Voigt 1958; König 1991; Zìtt and Nekvasilová 1993; Wittler 2001a; Helm and Schülke 2003; Löser 2003) or Cenozoic (e.g., Nielsen 1913, 1917, 1925; Langer 1989; Giammona and Stanton 1980; Kocurko and Kocurko 1992; Kocurko 1993; Bernecker and Weidlich 2005, 2006; Lozouet and Molodtsova 2008) sediments. The taxon Prographularia triadica from the Upper Triassic of the Northern Calcareous Alps was interpreted by Frech (1890), followed by Haas (1909) and Voigt (1958) as remains of an alcyonarian octocoral, an interpretation rejected by Jeletzky and Zapfe (1967), resulting in the lack of a confirmed octocoral record from the Triassic. This data set is also highlighted in the compilation of fossil octocorals provided by Sepkoski (2002) with one genus from the Palaeozoic, one genus from the Lower Jurassic, six genera (including Coptocamylodon as valid taxon) from the Lower Cretaceous and already 15 genera from the Upper Cretaceous expressing the pronounced phylogenetic radiation during this time. In detail, the data set of Sepkoski (2002) for the Lower Cretaceous lists four genera of the order Helioporacea (=blue corals) (Proheliopora Kuzmicheva, Polytremacis d’Orbigny, Pseudopolytremacis Morycowa, Epiphaxum Lonsdale), one genus from the order Pennatulacea (=sea pans) (Virgularia Lamarck) and two genera of the order Alcyonacea (Coptocamylodon Elliott, Corallium Cuvier). The rare available literature data, however, are not consistent and in parts also contradictory necessitating to add some further comments on the Sepkoski list and the proposed stratigraphic occurrences therein.
Heliopora de Blainville
This genus should appear in the Upper Cretaceous (Santonian) according to Sepkoski (2002). As a living fossil, the taxon Heliopora was treated in detail by Colgan (1984) with the oldest species being H. japonica from the Lower Cretaceous Miyako Group of Japan (Eguchi 1948). In the original work, however, no details on the stratigraphic position were indicated. Today, the stratigraphy of the Miyako Group is well established and the coral facies encompasses the Upper Aptian (e.g., Iba and Sano 2007). Recently, Heliopora japonica was transferred to the genus Pseudopolytremacis Morycowa (Löser and Ferry 2006; Tomás et al. 2008). Thus, the today-living Heliopora coerulea (Pallas) represents the oldest fossil record of this genus appearing in the Albian (Upper Greensand) of England (Duncan 1879).
Epiphaxum Lonsdale
The oldest described species of the genus Epiphaxum Lonsdale was E. labyrinthicum from the Albian of Texas; this taxon, however, belongs to the genus Polytremacis (Löser 2005, p. 259). Other described species of Epiphaxum belong to Aulopsammia Reuss (Löser 2000, p. 34). The type-species Epiphaxum auloporoides Lonsdale is well recorded from the Upper Cretaceous (Turonian) to Danian (Nielsen 1925; Voigt 1958; Wittler 2001a). Another fossil representative was recently described as Epiphaxum arbuscula from the Cenozoic (Eocene to Miocene) of France (Lozouet and Molodtsova 2008). The place of Epiphaxum within the Octocorallia is uncertain, and therefore regarded as an enigmatic genus (e.g., Daly et al. 2007). Löser (2005, p. 259) states, that Epiphaxum “cannot be claimed to be a coral at all”. Noteworthy, that the Helioporacea (blue corals) hold some kind of exceptional position as representing octocorals characterized by a massive aragonitic skeleton.
Corallium Cuvier
Our research could not confirm the supposed occurrence of the genus Corallium Cuvier in the Lower Cretaceous. The fossil representatives Corallium pindborgi (Nielsen 1925) and Corallium becki (Milne Edwards and Haime 1851) from the Danian of Denmark are both considered synonyms of Moltkia isis Steenstrup, 1846 (see Voigt 1958). The only remaining Cretaceous species is the Campanian-Maastrichtian Corallium elegantum Kuzmicheva, 1987.
Proheliopora Kuzmicheva
According to Löser (1993), Proheliopora Kuzmicheva represents a synonym of Pseudopolytremacis Morycowa.
Pseudopolytremacis Morycowa
P. spinoseptata, the type-species of the genus (Morycowa 1971), originally described from the Lower Cretaceous of Romania, was also reported from the Upper Jurassic (Oxfordian-Kimmeridgian) of Slovenia (Turnšek 1997).
Moltkia Steenstrup
Concerning the genus name, Moltkia, introduced by Steenstrup (1846), it should be noted that a homonym angiosperm plant of the family Boraginaceae exists (Lehmann 1817). Both represent so-called inter-code homonyms and are vaild, as both codes (ICZN-ICBN) are independent; only a recommendation to avoid the introduction of new generic names that already exist under another code exists (e.g., Hawksworth et al. 1994). When dealing with fossil (here: Cretaceous) octocorals, there arises the general difficulty of directly assigning them to extant genera. This will be shortly demonstrated with the couple Isis and Parisis.
Isis Linné—Parisis Verril
There are several species assigned to Isis Linné and described from the Upper Cretaceous and Paleogene (Nielsen 1913; Voigt 1958; Helm and Schülke 2003). The two Cenomanian species Isis tenuistriata (Reuss) and Isis miranda (Pocta) have been shown to represent differently shaped internodes of Moltkia foveolata (Reuss) (see Lehner 1937). Bayer (1955b, p. 211) already doubted the reference of some of the fossil representatives to Isis, assuming that they rather might belong to Parisis Verrill. Helm and Schülke (2003) illustrated a nice, almost completely preserved, specimen of Isis ramosa Voigt from the Campanian of northwest Germany showing a throughout rigid fan-shaped corallum completely different from recent Isididae with alternating calcareous internodes and horny nodes. Helm and Schülke (2003) and also Bernecker and Weidlich (2006) obviously escaped notice of the monography of Upper Cretaceous to Palaeogene corals of the former USSR provided by Kuzmicheva (1987). In this work, all the Upper Cretaceous-Palaeogene taxa originally described as species of Isis (including also Isis ramosa) were treated as representatives of Parisis, but Parisis is also characterized by alternating horny (nodes) and calcareous (internodes) skeletal parts (e.g., Bayer 1955b), thus, excluding the new combination concerning Isis? ramosa as proposed by Kuzmicheva (1987). The compact and strongly calcified skeleton (without horny nodes) and the branching colony shape of “Isis” ramosa rather points to the family Ellisellidae of the order Alcyonacea (e.g., Bayer and Deichmann 1960) offering the possibility that first representatives of this family could have originated in the Upper Cretaceous period. The colonial axis of “I.” ramosa should then show the occurrence of a radial pattern of calcification, being a characteristic feature of the Ellisellidae (Bayer 1955b). Already Bayer (1955b, p. 219) shortly discussed the possibility that the “Ellisellidae arose from ancestral isidids by elimination of the horny nodes” and with this respect was referring to the fossil genus Moltkia. Another example is related to Isis melitensis Goldfuss, 1826 considered as belonging to Parisis by some palaeontologists (e.g., Kuzmicheva 1987), yet others treat it as belonging to Keratoisis Wright, 1869 (e.g., Langer 1989; Noé and Dullo 2006). In conclusion, there is still much uncertainty amongst palaeontologists to which genera the fossil species actually belong. It also makes clear that with respect to fossil isidids, a generic determination based only on isolated calcareous internodes is not reliably possible.
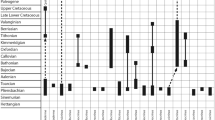
In Fig. 4 we tried to establish a stratigraphic range chart of Cretaceous octocoral genera being aware that the incomplete record and the controversial opinions about the systematics/synonymies of individual taxa lead to the provisional character of the chart presented. It shows that the evolution of the modern octocorals started at the end of the Lower Cretaceous (genera Heliopora) and more pronounced during the Upper Cretaceous (genera Epiphaxum, Nicella, Corallium, Parisis). The compilation includes two genera that are only known from the Cretaceous, Krimella Kuzmicheva and Paramoltkia Löser. These two taxa are also so-called singletons, genera occurring in only one stage (Kiessling and Baron-Szabo 2004). It can be assumed, however, that this only results from the poor state of knowledge expressed also by the fact that both were so far only reported from their type localities. In any case, the short discussion of Cretaceous octocorals in general, the dubious records of some genera from the Lower Cretaceous and the taxonomic problems when referring fossil to recent taxa highlights every new finding of any stratigraphic interval at all and especially those older than the Upper Cretaceous.

Simplified Cretaceous stratigraphic range chart of octocoral genera (Pennatulacea excluded), critically revised on the basis of bibliographical data (Voigt 1958; Kuzmicheva 1987; Löser 2000; Sepkoski 2002). Note that the genus Primnoa Lamouroux was excluded as it is considered to lack fossil representatives (see Voigt 1958; Cairns and Bayer 2005, 2009, and details in the text). The compilation shows that the evolution of the modern octocorals started in the Lower Cretaceous (genus Heliopora) and more pronounced during the Upper Cretaceous (genera Nicella, Corallium, Parisis, Epiphaxum)
The numerous records from the Upper Cretaceous to Palaeogene, especially the Maastrichtian–Danian interval, are mainly from marly or carbonatic deep-water sediments but also from the littoral zone (e.g., Nielsen 1913, 1917, 1925; Voigt 1958; Leloux 1999). Whereas all these occurrences are from the boreal realm, e.g., the Danish Basin, with typical cool-water carbonates (e.g., Bernecker and Weidlich 2005, 2006; Bjerager and Surlyk 2007), proved occurrences from shallow-water carbonates of the (sub-)tropical Tethyan realm are virtually lacking. Within the marly Upper Cretaceous lithologies, the occurring dissociated remains of octocorals and recovered from washing samples or extracted from the carbonates (e.g., Nielsen and Jacobsen 2004) can be well ascribed to their basal holdfasts and internodes whereas findings of their sclerites are comparably seldom (e.g., Alexandrowicz 1977) and findings of more or less complete specimens represent a real rarity (Bayer 1955a: sea-pens; Helm and Schülke 2003). The sclerites of Recent octocorals embedded in coenenchyme, the colonial tissue between the polyps and consisting of mesogloea (see Bayer et al. 1983), are one of the most important features for their taxonomic classification (e.g., Janes and Wah 2007). In some families, their arrangement and occurrence within the whole colony is “as important as the shape of the spicules themselves” (Bayer 1956). This approach can understandably not be applied to fossil material; they can just be classified and described according to the glossary of Bayer et al. (1983). Therefore, the classification of fossil octocoral sclerites established by Deflandre-Rigaud (1955, 1956, 1957) is irreproducible and taxonmonically misleading as already remarked by Giammona and Stanton (1980, p. 73). This scheme refers to mainly spindle-shaped sclerites that should all belong to the only one morpho-genus Micralcyonarites Deflandre-Rigaud, 1955, with more than 20 different form-species.
Sclerites of modern octocorals are composed of tightly packed crystals of calcite with variable amount of magnesium (high Mg-calcite) (e.g., Weinbauer and Vellmirov 1995; Rahman and Oomori 2008), but also calcite with an admixture of aragonite or aragonite alone (e.g., Bayer 1955b, 1981a). Stanley et al. (2002) conclude that the Mg content in the skeletal calcite of anatomically simple organisms correlates directly with the Mg/Ca ratio of the ancient seawater. For groups such as the alcyonarians (with high-Mg calcite today), the authors assume that they probably produced skeletons with low-Mg calcite in Late Cretaceous seas. In addition, also the temperature of the seawater can be a factor influencing the skeletal Mg/Ca ratio (Sherwood et al. 2006). Such a complex interplay of factors, however, should only have an impact when comparing shallow-water with deep-water taxa.
The internodes/branches maybe purely calcareous, such as in the isidids, or with a central axis composed of scleroprotein, gorgonin, and hydroxylapatite (e.g., Lewis and Von Wallis 1991; Macintyre et al. 2000; Bayer and Macintyre 2001). Bayer and Macintyre (2001) suppose that the axes and holdfast mineralogy could be of importance for octocoral systematics. Such an assumption was already discussed by Kocurko (1988) stressing that the microstructure of the holdfasts may be aragonitic, calcitic or composed of calcareous layers alternating with gorgonin. Noteworthy, that within four recent families (Ellisellidae, Ifalukellidae, Chrysogorgiidae, Primnoidae) the latter type of holdfast microstructure is prevailing (Grasshoff and Zibrowius 1983). By all means, the original mineralogy and microstructural features are difficult to assess in fossil taxa as shortly discussed above. It is likely that the remains of fossil octocorals “are not as rare as previously thought” (Kocurko 1988, p. 114), but obviously often go unrecognized, although especially their calcified sclerites, axes, and holdfasts have a fairly good potential for preservation. Our contribution is therefore of special interest and represents a further jigsaw piece for resolving the question recently put forward by Reich (2007): “Where are all the fossil octocorals”?
Coptocampylodon–Carpathoporella: a historical review
The taxon Coptocampylodon was established by Elliott (1963) with the type species C. lineolatus (Lower Cretaceous of Iraq) as follows: “Small solid cylindrical calcareous bodies, longer axis gently curved or irregular, circular in cross section but deeply incised by longitudinal grooves, ends irregularly rounded”. The material of Elliott collected from two different formations contained isolated specimens (holotype) from the Hauterivian and some specimens recovered from Lower Cretaceous thin-sections (paratypes). The isolated, slightly curved specimens (with rounded ends) exhibiting longitudinal ribs are comparable large, attaining diameters of up to 1 mm and lengths of up to 3 mm, whereas the transverse sections in the thin-section material only attain diameters of up to 0.5 mm. As one discriminating feature of “species” established later on, the number of the marginal-longitudinal grooves became important (e.g., Patrulius 1966; Dragastan 1989; Schlagintweit et al. 2002; Ljubović-Obradović and Radoičić 2003). Elliott (p. 298) concluded that Coptocampylodon “comprises the skeletal remains of a small octocoral in which horny and calcareous joints alternated”.
A second representative was described by Patrulius (1966) as Coptocampylodon fontis from the Lower Cretaceous of Romania differing from the type species by a higher number of grooves (9–15) and the often-occurring central canal. Noteworthy, the variability of C. fontis also comprises solid specimens lacking this feature. The external diameters (0.20–0.42 mm), however, are in the range of the paratype specimens of C. lineolatus. As the occurrence of a central canal is not compatible with the generic diagnosis of Coptocampylon, C. fontis cannot be included in the latter without emendation by incorporating also non-solid forms, thus, enlarging the generic diagnosis or—if one considers this as a criterion of generic importance—establishing a new genus. By the way, the occurrence of a central canal was also the reason for Patrulius to doubt an interpretation as skeletal elements of Octocorallia as done by Elliott for Coptocampylodon lineolatus. Last but not least, mention should be made that Patrulius did not designate a holotype, thus, Coptocampylodon fontis represents a nomen nudum (see Granier and Deloffre 1993). In 1967, Dragastan introduced the genus Carpathoporella (type species C. occidentalis) from the Lower Cretaceous of Romania by taking the existence/absence of a central canal as generic criterion and assigned it to the dasycladalean algae; a holotype was not indicated, thus representing a nomen nudum. Radoičić (1969) described another new species as Coptocamylodon elliotti from the Middle Cretaceous of the Dinarides, also possessing a central cavity. Radoičić draw the following conclusions: (a) Carpathoporella occidentalis Dragastan is identical with Coptocampylodon fontis Patrulius; (b) Coptocampylodon lineolatus Elliott corresponds to tufts of secondary laterals of Triploporella-type dasycladalean algae; and (c) representatives of Coptocampylodon might belong to the debris of different groups of organisms. In his classification of fossil dasycladaleans, Deloffre (1988) treated Coptocampylodon with C. fontis as a possible dasycladale alga (family Acetabulariaceae) and Carpathoporella as a “mélange des formes” excluded from the dasycladales.
The identity of Coptocampylodon fontis and Carpathoporella occidentalis was later accepted by Dragastan (1989) introducing the new combination Carpathoporella fontis (Patrulius). Also, Coptocampylodon elliotti was considered as belonging to the genus Carpathoporella. Regarding the biogenic origin of Carpathoporella fontis, Dragastan envisaged a possible characean nature. In their revision of Jurassic-Cretaceous dasycladalean algae, Granier and Deloffre (1993) stated the invalidity of Carpathoporella Dragastan, 1989 nom. nud. based on Carpathoporella fontis (Patrulius) as representing a homonym of Carpathoporella Dragastan, 1967 nom. nud. based on Carpathoporella occidentalis. Regarding Coptocampylodon lineolatus Elliott, the coprolite nature for the isolated holotype, as suggested by Cuvillier et al. (1969), was supported by Granier and Deloffre (1993); the paratypes from the thin-sections, however, should correspond to algal spicules. The genus Carpathoporella with the type species C. occidentalis was validated later by Dragastan (1995) by choosing a lectotype from the original material (pl. 1, fig. 9, in Dragastan 1967) in accordance with article 9.2 of the International Code of Botanical Nomenclature.
In 2002, another possible representative of Coptocampylodon Elliott was described as Coptocampylodon? rhaeticus by Schlagintweit et al. (2002) from the Upper Triassic of the Northern Calcareous Alps; it was interpreted as “parts…of a larger organism of unknown systematic position”. Ljubović-Obradović and Radoičić (2003) introduced the new species Coptocampylodon pantici (Turonian of Serbia), and emended the morpho-genus Coptocampylodon, indicating its origin as “dislocated tufts of the elongate trichophorous secondary laterals of some dasycladalean species, perhaps even of several genera”. Whether all Coptocampylodon “species” represent laterals of larger dasycladales, however, was later questioned by Schlagintweit and Gawlick (2007). Besides the interpretation as octocoral sclerites by Elliott (1963), a view not followed by subsequent workers, Coptocampylodon was—as already mentioned—interpreted as algal remains and the isolated specimens as crustacean coprolites (Cuvillier et al. 1969). The latter opinion was regarded as a simple case of homeomorphism by Ljubović-Obradović and Radoičić (2003). Živkovic and Bogner (2006) studied isolated specimens of Coptocampylodon lineolatus from the Eocene of Croatia demonstrating that these show similar chemical composition as the host marl. Together with the detection of coccoliths within C. lineolatus, a sediment-derived nature (deposit-feeding!) is evident, supporting the interpretation as crustacean coprolite. As lacking any internal structures, C. lineolatus cannot be compared with the different taxa showing genus and species characteristic internal canals (e.g., Cuvillier et al. 1969; Brönnimann 1976).
The specimens described in the present paper can be assigned to Carpathoporella occidentalis Dragastan 1995 (non 1967, 1989). A detailed synonymy of this taxon is included in Schlagintweit et al. (2002).
Description of material
Sample Al 847 (thin-sections Al 847-a and -b) from which the material illustrated and discussed in the present paper derives, represents a poorly washed-out packstone with debris of “Coptocampylodon-Carpathoporella” in great abundances, remains of gastropods and corals, Bacinella-Lithocodium nodules, dasycladalean algae such as Salpingoporella cf. pygmaea (Gümbel) and some benthic foraminifera (Nautiloculina sp., Vercorsella sp.) (Fig. 3g, h). The sections of Carpathoporella are totally recrystallized, consisting of sparry calcite. Most of them display spherical shapes. With respect to occurring frequencies, star-like sections with commonly five deeply lobed margins (pentamer symmetry) are comparably rare (Fig. 5a–c). Mostly, the edges are even, rarely concave, the space between is typically V-shaped with a rounded base. Usually, the star-like specimens are fairly small in diameter (0.18–0.25 mm) and can be well compared to the small paratypes of “Coptocampylodon lineolatus” or Coptocampylodon sp. 2 of Radoičić (2005) from the Valanginian of the Mirdita Zone of Serbia, Carpathoporella? sp. from the Lower Cretaceous of the Mid-Pacific Mountains (Masse and Arnaud-Vanneau 1999) or the supposedly “second order laterals of Selliporella neocomiensis” from the Valanginian of Iran (Hosseini and Conrad 2008). The vast majority of spherical sections have 8–12 marginal grooves of hemi-spherical to ovoid shapes, generally showing resemblances to transverse sections of aciculariacean algae (Fig. 5c pars, d–j). Normally, these forms are larger (diameters up to 0.6 mm) than the star-like sections, but there also exist rare tiny forms (diameter 0.14 mm) (Fig. 5c). Besides solid specimens, there are also forms showing a central hollow (diameter 0.075–0.185 mm) occupying about 25–45% of the total diameter (0.25–0.45 mm) (Fig. 5k–n, o pars). The number of marginal grooves varies between seven and 12. This type is here called “gearwheel typus”. The dimensions/data are in accordance with those indicated for Carpathoporella occidentalis or “Coptocampylodon” fontis nom. nud. (Patrulius 1966; Dragastan 1967, 1989). In some cases, the central hollow is not totally surrounded by calcite but shows an open side. Such specimens may occur free within the sediment (Fig. 5m) or as part of the basal fixed portion (see below).

a–y Specimens known as Carpathoporella occidentalis Dragastan, interpreted as octocoral skeletal elements. a–c (pars) Star-like sclerites showing pentamer symmetry. d–j Tuberculated spheroids, some showing resemblances with aciculariacean algae (e.g., i, j). k–o Skeletal elements with central hollow and marginal grooves. Note the opening in m, comparable to the occurrence within the basal holdfast (compare Fig. 6a). p–y Sections of longitudinally grooved internodes or axes. Note rounded hollows in p and s interpreted as calixes and internodal branching in y. Scale bars 0.2 mm for a–c, k; 0.3 mm for d–j, l–o; 0.5 mm for p–y. Sample Al 847a: c–e, h–j, n, p–r, t–v, y; Sample Al 847b: a, b, f, g, k–o, s, w, x
Another group of skeletal debris, and interpreted as longitudinal sections, shows cylindrical, straight-to-curved tubes (diameter 0.14–0.25 mm: wall thickness 0.048–0.080 mm), also exhibiting branching (Fig. 5p–z). They arise from the “grooves” of similar-shaped “gearwheel type” with a small central hollow (e.g., Fig. 5p). Usually known from the Barremian-Aptian interval, comparable sections were illustrated by Bucur et al. (2000) from the Berriasian-Hauterivian of Turkey. Extremely rare are findings where parts of the described calcitic elements are fixed to hard substrates (e.g., shells, metazoan skeletons). The attached part gains a width of 1.3 mm and a maximum height of 0.45 mm (Fig. 6a). It contains several depressions (four visible in thin-section) from which obviously several, although not preserved branches were arising. There are some smaller depressions (width about 0.1 mm) with acute-rounded endings and a somehow larger one (width about 0.3 mm) that is broadening at its base. Two fused tuberculated bodies (e.g., Fig. 5f) are still in contact with one of the smaller depressions (Fig. 5b). Within the attachment structure, these “gearwheel types” with a central hollow occur. The originally attached basal parts were also detected detached in the micritic matrix (Fig. 6c).

a Basal holdfast fixed to the substrate with several pits (1–4) representing the contact points of several branches, one containing two fused sclerites (4). Note the skeletal element with central hollow (arrow) and the basal joint-like broadening (1, left). The possible occurrence of a star- or cross-like sclerite is visible on the right (4). Scale bar 0.5 mm. b Detail of the holdfast (h) showing two fused sclerites (s) fixed to one of the pits. Scale bar 0.3 mm. c Detached basal holdfast not fixed to the substrate. Scale bar 0.5 mm. All samples Al 847a
Discussion
It is evident that the evidenced morphological characteristics are not compatible with an algal origin of Carpathoporella occidentalis. Instead, we interpret them as representing different skeletal elements of a colonial octocoral, as already Elliott (1963) did for his paratypes of Coptocampylodon lineolatus.
The spheroid bodies that make up the main part of the debris can be assigned to octocoral spicules, microscopic elements embedded in the soft tissue (coenenchyme). Following the terminology of Bayer et al. (1983), the majority of our specimens can be classified as tuberculate spheroids (Fig. 7a). Extremely rare are spheroid bodies that at one side are elongated and fitting into the insertions of the basal holdfast (Fig. 6b). This type, morphologically comparable to the double-cone type (Fig. 7b) only occurs as the first spicule connected to the holdfast, already the succeeding second sclerite is roughly spherical in shape. The grooves and ridges visible in thin-sections can be ascribed to a tuberculated surface ornamentation (e.g., Fig. 7a). Those forms with a star-like outline can be compared to the typus “stellate plate” that are flat sclerites with deeply incised margins, more or less radially symmetric, also of pentamer shapes (e.g., Bayer et al. 1983, fig. 201). From recent representatives, spicules with three (triradiate), four (cross), five (stellate plate), or six (sixradiate) margins are occurring (Fig. 7c). Whether our specimens are actually flattened plates cannot be answered from the thin-section material, as three-dimensional data are not available. Their rareness, however, can be an indication for it as the accidental sectioning exactly in the sclerite plane would be extremely rare as expected. These star-like bodies were reported already from Tithonian strata, but this material lacks forms with a central hollow (Schlagintweit and Gawlick 2007). Sclerites of Recent octocorals are very variable in diameter but often occur in different size classes (e.g., Williams 2003). Our specimens show a normal distribution with sizes ranging from 0.1 to 0.6 mm, and most values between 0.3 and 0.4 mm (Fig. 8). Their shape and distribution throughout the octocoral colony is a result of the role they serve in the mechanical system of the soft components (Lewis and Von Wallis 1991) accounting also for their intracolonial or intraspecific variability. The observed fused tuberculated sclerites in direct connection to the basal holdfast (Fig. 6b) may allow a flexible bending on a limited scale responding to water currents and more generally of course also as internal support to hold the colony erect especially at the base for ensuring the fixation to the basal substrate. In modern octocorals, the area of the sclerites within the colony plays an important role for correct identification (e.g., Janes and Wah 2007), usually impossible to determine for the fossil isolated, dispersed elements. Only in our case, the position within in the colony is evidenced. Their arrangement points to an axis composed of fused spheroids that would be an explanation for their overwhelming dominance in the material studied.

Examples of spicules (a–d) of recent octocorals and holdfasts of Upper Cretaceous representatives (e, f). a, b Tuberculate spheroids (spicules); left: from the holdfast region of Eleutherobia flammicerebra; Recent (from Williams 2003); right: from Euplexaura; Recent, (from Bayer 1956). c Double cone (sclerites); from Euplexaura; Recent (from Bayer 1956). d Crosses (above, below) and star (middle), here: five-radiate; from Ellisellidae and Anthothelidae; Recent (from Bayer 1956). e Holdfast of Moltkia sp. with diverging root-like prolongations; Upper Cretaceous (from König 1991). f Holdfast of Primnoa? costata Nielsen (? Epiphaxum, compare Voigt 1958) showing simple volcano-shape; Upper Cretaceous (from König 1991). Note that in both cases (e, f) the attribution of the holdfasts to distinct genera or species is proposed by the authors and not proven with absolute certainty

Size of sclerites of Carpathoporella occidentalis Dragastan, Lower Cretaceous of Albania, sample Al 847. Diameter in mm, n = 75. Note, that the belonging to some measured data to internodes (without central canal) cannot be excluded (see discussion in the text)
The cylindrical, slightly bending, longitudinally grooved and occasionally branching parts (e.g., Fig. 5j) may be ascribed to internodes or axis, that is the octocoral’s supporting calcareous structure. Internodes of Moltkia Steenstrup or branches of Epiphaxum auloporoides Lonsdale, for example, possess longitudinal furrows and the thickened endings exhibit calyces cavities or depressions (so-called Kelchgruben, e.g., Voigt 1958; Löser 2003) with jagged borders that could be similar to our sections of the “gearwheel types” (Fig. 9). In the juvenile parts, the calix insertions occur at both endings of each internode, whereas in the more adult parts they also occur along the sides of the internodes. The calix insertions may also be arranged oblique or perpendicularly to the internode longitudinal axis (see Voigt 1958 for details). This constellation could be an explanation for the thin-section specimens where both the longitudinal grooves-ridges and a small central “hollow” (? transverse section of a calix impression) at the endings (or between) are discernible (e.g., Fig. 5p or s). The dimensions of the calcareous internodes are variable between different species; the internodes of Moltkia for example have diameters of about 0.3–2.5 mm, the thickened ending 1–1.2 mm and the diameter of the calix insertions amounts 0.5–0.9 mm (Voigt 1958; Löser 2003). An axial canal my be present or become strongly reduced (see Fig. 3 in Voigt 1958). Therefore, some sections could in fact belong to transverse sections of internodes without axial canal (e.g., Fig. 5e), but such a possible interpretation cannot be verified here. Noteworthy is that the axial canal of Moltkia is subdivided by transverse partitions. Some of these features can be assigned to a specimen of “Carpathoporella fontis” illustrated by Radoicic (1969, pl. 3, fig. 3) (Fig. 9a). Accepting such an interpretation, the calcareous bodies showing a central hollow (e.g., Fig. 5k) could belong to both transverse sections of internodes/branches or aborted calix insertions. Remarkably, similar transverse and longitudinal sections were also illustrated by Radoičić (1969) for Coptocampylodon elliotti from the Albian of Serbia. A more detailed comparison, however, is unfortunately hindered by the fact that internodes of fossil octocorals are known from isolated specimens obtained from washing samples of marls, whereas representatives from thin-sections are—according to our knowledge—unknown from the literature. According to Löser (2003, p. 99), nothing is known about the calcareous spiculae of Moltkia, a further disadvantage for comparison with our material. Furthermore, the author states that from a palaeobiogeographic viewpoint, Moltkia is restricted to the boreal realm. This observation conforms to the general circumstance that from this area the octocorals are mainly recorded from marly lithologies and not from shallow-water carbonates, widespread in the Tethyan realm during both the Lower and Upper Cretaceous period. Here, skeletal remains of Carpathoporella-Coptocampylodon are rather widespread during the Lower Cretaceous, reported from Albania (this work), Austria, Bulgaria, Croatia, Greece, Romania, southern France, southern Germany, southern Italy, Spain, Serbia, and Venezuela (see synonymy in Schlagintweit et al. 2002). Besides representatives of the genus Moltkia, also fragments of branches or stems of Primnoa? costata or Epiphaxum auloporoides Lonsdale of the Danish Danian possess a surface covered with longitudinal ribs separated by deep furrows (Nielsen 1917, 1925) are also comparable to our sections (Fig. 9b, c). Especially the serrated outline of the calyces shows conspicuous similarities to Carpathoporella occidentalis, therefore offering further comparisons and interpretation (Fig. 9b). The holdfast of Primnoa? costata reported by Malecki (1982), however, is different from our finding as representing a simple disc with a central depression (Fig. 7e) and also the central canal of the internodes is typically star-like in transverse sections (Voigt 1958: pl. 2, fig. 4).

Internodes/branches of fossil octocorals. a Longitudinal section of Carpathoporella occidentalis (Dragastan) (from Radoičić 1969) and interpretation in terms of octocoral morphology. Between brackets terminology of Voigt (1958) for internodes of Upper Cretaceous Moltkia species (see also Fig. 9c). b Internode of Epiphaxum auloporoides Lonsdale (=Primnoa gracilis Nielsen according to Voigt 1958) showing longitudinal ribs and grooves (from Nielsen 1925). c Internode of Moltkia minuta Nielsen showing longitudinal ribs and grooves (from Voigt 1958)
Growth stages, expressed by concentric rings/bandings and reported for instance from extant and fossil gorgonians, particularly sections of their internodes (Langer 1989; Noé and Dullo 2006; Sánchez et al. 2004; Noé et al. 2007), or any other microstructural features (e.g., Bayer 1981a; Majoran 1987; Lewis et al. 1992; Macintyre et al. 2000; Sethmann et al. 2007) are not visible in our specimens due to recrystallization. The observable homogeneous sparry calcite, however, makes an original alternation of horny and calcareous material improbable.
The basal part (Fig. 6a) can be ascribed to a holdfast, one special style of attachment strategy to hard substrates (see overview of Bromley and Heinberg 2006). Generally, it is a rod-like anchorage structure that fixes sessile organisms, such as seaweed, some calcareous algae (e.g., Funaki et al. 2001; Anderson et al. 2006), stalked crinoids or sponges to a substrate that can be muddy, sandy, or rocky (Fig. 10a, b). In the fossil record, especially holdfasts from crinoids are well documented (e.g., Franzén 1977; Radwanska and Radwanski 2003; Wittler 1998, 2001b) (Fig. 10c, d) but also from octocorals, mainly from Upper Cretaceous and (Early) Cenozoic sediments (e.g., Malecki 1982; Langer 1989; König 1991; Kocurko and Kocurko 1992) (Fig. 7d, e). Some of them also have the form of what we termed “gearwheel-type” elements (Fig. 7f). There are circular holdfasts with one central insertion of the axis that can be best characterized as volcano-type or irregular forms with diverging root-like prolongations and numerous round, crater-like depressions corresponding to different insertions of axes and other intermediate forms. A simple volcano shape of the holdfast results from the post-mortem loss of the erect main branch. The surface of the holdfast may be smooth or covered with ridges or striae (e.g., Malecki 1982; Kocurko 1988), a feature not observable in our thin-sections cutting the holdfast in a vertical plane perpendicular to the substrate. Our findings are composed of coarse-grained calcite; the mineralogy of modern octocoral holdfasts maybe highly variable (aragonitic, calcitic, calcareous with alternations of gorgonin, e.g., Kocurko 1988).

Examples of recent non-calcified and fossil calcified holdfasts. a Modern seaweed (kelp) with (from above to below) blades (leafs), stipe and basal holdfast. Coin diameter = 2.8 cm. Atlantic coast of Nova Scotia, Canada. b Detail from a showing the root-like non-mineralized holdfast. c, d Calcitic holdfasts of fossil crinoids from Upper Jurassic platform margin deposits, Mount Rettenstein, Northern Calcareous Alps (see Auer et al. 2008 for details). Northern Calcareous Alps of Austria
These holdfasts or bases are usually found isolated or still in contact with the substrate alone without any other parts of the octocoral. Therefore, they were classified by Malecki (1982), according to parataxonomic criteria, with the morpho-genus Octobasis including 11 morpho-species based on material from the Lower Campanian of Poland; a type species has not been fixed by Malecki (1982). The width of the holdfasts (formerly known as bases; see Bayer et al. 1983) described by Malecki ranges from 0.8 to 8 mm, and up to 30 mm from a holdfast ascribed to the genus Isis; the diameters of the depressions vary from 0.06 to 1.2 mm. Reminding that our form (Fig. 6a) has a visible width of 1.3 mm and insertion diameters of 0.1–0.3 mm, thus falling within the ranges reported by Malecki. This approach was viewed critically by König (1991) doubting the validity of all of these morphotaxa. According to König (1991), ramified forms with an ornamentation composed of fine radial striae such as Octobasis lobata, O. dichotoma, O. sulcata, and O. flexuosa could belong to the genus Moltkia (Fig. 7d). This conclusion, however, is only an assumption because of the ornamentation of both the internodes (Fig. 9) and the bases (Fig. 7d). As these holdfasts, however, were described as isolated elements not in contact with other parts of the animals, such interpretations need final confirmation for example by findings of complete specimens (e.g., Helm and Schülke 2003). Based on the holdfast structure of our findings with numerous insertions of axes, the former colony shape can be classified as bushy, meaning abundant branches arising immediately above the holdfast and diverging upwards, not forming an obvious single main stem (see Bayer et al. 1983). On the other side, we cannot exclude the possibility that we are dealing with two different taxa, one with calcareous internodes/branches and the other with axes composed of fused sclerites, where a related basal holdfast has been detected. The greater width at the base of one of the insertions forming a joint likely has served for a better stability and protection against damage. In cases of strong currents, a comparable large leverage effect must result from the bending especially of bushy forms. Anyway, the specimen showing two fused spicules in contact to the basal holdfast represents an extraordinary finding (conservation perhaps resulting from rapid sediment burial?) as Bayer (1956) in the Treatise on invertebrate paleontology remarked: “No specimen of fossil octocoral in which the original arrangement of spicules has been preserved is known to me”.
General taxonomic and phylogenetic considerations
The branching colony morphology, the attachment by a basal holdfast, supporting skeletal axes and the fused sclerites (at least in the parts directly contacting the holdfast) account for the order Gorgonacea (“sea fans”) (e.g., Grasshoff and Bargibant 2001; DeVictor and Morton 2007), respectively, the suborder Scleraxonia (see Bayer 1981b). Other classifications put the soft corals and gorgonians in one single order Alcyonacea (see e.g., Fabricius and Alderslade 2001; Williams and Cairns 2006; Daly et al. 2007 for details). Calcified holdfasts are known from several gorgonacean families (in alphabetical order): Chrysogorgiidae, Ellisellidae, Gorgoniidae, Isididae, and Plexauridae (e.g., Grasshoff and Zibrowius 1983). Referring Carpathoporella Dragastan to a distinct genus of Octorallia is not possible for the moment, because of the isolated and dispersed skeletal elements and the lack of knowledge of the exact colony morphology. Anyway, the sections interpreted as internodes or branches show affinities to the genus Moltkia Steenstrup, 1846 (e.g., Voigt 1958; König 1991; Löser 2003) known so far from the Cenomanian to Eocene interval (König 1991) or the taxa described by Nielsen (1913, 1925) as Primnoa costata and Primnoa gracilis from the Maastrichtian-Danian. It should be noted that the generic position of the two “Primnoa” species is still doubtful. Voigt (1958) convincingly demonstrated that Primnoa gracilis Nielsen, 1925 only represents the branches of Epiphaxum auloporoides Lonsdale, 1850 and thus a synonym of the former (see also Bayer 1979, 1992), a view obviously not followed by Kuzmicheva (1987) and Bernecker and Weidlich (2006). Recent representatives of Primnoa have massive stems showing concentric laminae (growth rings) in cross sections (Andrews et al. 2002; Cairns and Bayer 2005, 2009), whereas the central part in the Maastrichtian-Danian taxa is hollow, for instance with star-shaped outline in “Primnoa” costata (Voigt 1958). Hence, none of the two Nielsen taxa were included in the “review of the genus Primnoa” provided by Cairns and Bayer (2005) with only Recent species. The taxonomic status of the Danian “Primnoa” costata needs revision.
The observed internodal branching (Fig. 5y) is well known from extant isidids (“bamboo corals”) (e.g., Bayer 1990; Noé and Dullo 2006; France 2007; Etnoyer 2008) a group of octocorals characterized by jointed axes composed of alternating gorgonin nodes and calcitic internodes (see Table 2); the basal holdfasts are root-like. In the fossil record, isolated internodes/branches showing branching were reported from Upper Cretaceous representatives of Epiphaxum Lonsdale, Parisis Verril, Moltkia Steenstrup and ?Primnoa Lamouroux (Voigt 1958; Kuzmicheva 1987; Helm and Schülke 2003). According to Voigt (1958), such findings are so far not reported earlier than the Upper Cretaceous. Bayer (1955b, p. 219) puts the interpretation of Moltkia as representing remains of an ancestral isidid octocoral up for discussion; others directly denote Moltkia as a fossil isidid (e.g., Grasshoff and Zibrowius 1983; Bernecker and Weidlich 2006). Due to clear morphological analogies to taxa like Moltkia, a possible isidid origin might be possible also for the Lower Cretaceous Carpathoporella occidentalis. So far, both Moltkia and Isis (or Parisis) are unknown from strata older than the Upper Cretaceous (Sepkoski 2002) (see Fig. 4). Generally, remains of isidid octocorals also belong to the most common forms with special preservation potential due to their extensive calcification (Kocurko 1988; Squires 1999). The calcification of the isidid internodes, however, being calcitic and showing concentric growth rings in transverse sections (e.g., Sánchez et al. 2004; Andrews et al. 2005) is altogether different from Carpathoporella. The skeletal elements of Carpathoporella occidentalis consist of coarse-grained sparry calcite without any microstructural features, pointing to diagenetic alteration of primary aragonite, as for example typical also for dascladalean algae (with some exceptions). If Carpathoporella have consisted of primary calcite, this would have been preserved with microstructural features or perhaps having a yellowish appearance in thin-sections (e.g., Noé and Dullo 2006). Therefore, analogies consist of the branches (showing furrowed exterior) of fossil species of Epiphaxum (see Fig. 9b), known since the Albian (Wells 1932) and consisting of aragonite (Lozouet and Molodtsova 2008). Also, the branch cross sections of Epiphaxum auloporoides with their central rounded hollows (different from Primnoa? costata, which has an inner star-shaped hollow) are comparable to sections of Carpathoporella.
The fact that Carpathoporella is a shallow-water taxon does not necessarily speak against any relationships to the deeper water taxa Epiphaxum or Moltkia as it has been evidenced from extant taxa that even within a single genus a gain or loss of the algal symbiosis may occur in their evolutionary history (Van Oppen et al. 2005). Moreover, a final conclusion is hindered by the impossibility of a 1:1 comparison of our thin-section material and the isolated Upper Cretaceous material showing all three-dimensional characteristics. If a direct assignment to Carpathoporella Dragastan to one of the above-mentioned taxa could be demonstrated in the future, however, it would become invalid as these were all described much earlier.
In Fig. 11, the database for Lower and Upper Cretaceous octocoral genera of the three orders Alcyonacea, Helioporacea, and Pennatulacea and the possible relationships of the genus Carpathoporella are presented. In any case, there is no confirmable evidence for the appearance of the order Alcyonaria prior to the Upper Cretaceous, whereas both Helioporacea and Pennatulacea are well recorded from the Lower Cretaceous (see also Figs. 4, 11).

Critical inventory of Lower and Upper Cretaceous octocoral genera (on the basis of Voigt 1958; Kuzmicheva1987; Löser 2000; Sepkoski 2002) and the possible relationships of the genus Carpathoporella Dragastan. Neither the doubtful occurrence of Primnoa in the Upper Cretaceous nor the holdfast parataxa established by Malecki (1982), which most likely belong to already-existing not to new genera, were included (details in the text). The Sepkoski compendium also lists Axogaster Lonsdale, 1850 as a valid Upper Cretaceous genus. It is a synonym of Moltkia Steenstrup (see discussion in Voigt 1958) and therefore is also not included. According to Williams and Cairns (2006) Pavonaria Kölliker (included in the Sepkoski list) represents a synonym of Halipteris Kölliker. The doubtful occurrence of Virgularia Lamarck in the Lower Cretaceous (see Sepkoski 2002) could not be confirmed; this taxon is reliably reported since the Campanian (e.g., Reich and Schneider 2002)
In most of the recent families and even genera of octocorals both types with the presence (zooxanthellate) and absence (azooxanthellate) of algal endosymbionts occur. As the ancestral state of algal endosymbionts in octocorals is unclear (e.g., Van Oppen et al. 2005, for details) and as both groups are occurring in modern reefal environments, the exclusive shallow-water findings of Carpathoporella do not allow a final conclusion with this respect. It is worth mentioning that Moltkia and Isis (or Parisis) are considered as belonging to the azooxanthellate group of octocorals (Bernecker and Weidlich 2005; Bjerager and Surlyk 2007). Van Oppen et al. (2005, p. 2415) stress that “many gains and/or losses of the algal symbiosis have occurred during the evolution of octocorals, even within single genus” probably because of a “low photosynthetic efficiency”. In general, corals that occur below the photic zone are all azooxanthellate, but as deep-water corals were shown to have invaded also shallow-water areas (e.g., Lindner et al. 2008: “from offshore to onshore”), this could be one explanation for changes (presence/absence) of algal symbiosis. Can it be speculated that the Upper Cretaceous deep-water octocorals from the boreal realm originated from a completely opposite onshore/offshore evolutionary pattern from Lower Cretaceous shallow-water tropical ancestors? This would be just another example of a principal evolutionary pathway reported already from a variety of other taxonomic groups (see e.g., Jablonski et al. 1983; Jacobs and Lindberg 1998 for details). The lack of skeletal elements identical or similar to Carpathoporella in Lower Cretaceous deep-water sediments would fit such an interpretation. Also noteworthy is that the three genera of the Helioporacea reported from the Lower Cretaceous (see Fig. 4) are from shallow-water carbonates of the Tethyan realm. The analysis of the existing database by Kiessling and Baron-Szabo (2004) has shown that the K/T boundary event caused an approximate extinction of 30% of scleractinian corals at the genus level, with a much stronger affection of shallow-water zooxanthellate taxa than deeper water forms lacking algal symbionts. In that way, Bernecker and Weidlich (2005) also remarked that the Maastrichtian to Danian deep-water octocorals did not show a major change at the generic level across the K/T boundary. This survival obviously was first enabled by the proposed onshore/offshore evolution assumed to have happened around the transition from the Lower to the Upper Cretaceous. Such a hypothesis is not in contrast to findings of Upper Cretaceous octocorals within shallow-water settings, but just documents a persisting occupation of this palaeoenvironment since Lower Cretaceous times.
In Fig. 12 we plotted the distribution of octocoral genera during the Cretaceous based on the stratigraphic chart presented in Fig. 4. All uncertain occurrences within individual stages were treated as definite records for the purpose of convenience and comparability. We are aware that this compilation certainly has a preliminary character, but it is an attempt to classify and interpret the researched bibliographical data also for future contribution of subsequent authors. The curve obtained from connecting the data shows a successive appearance of new genera during the entire Cretaceous period. There is no stage where the total number of genera is less than in the previous stage, which would document some kind of decline, e.g., biological crisis. For example, the Cenomanian-Turonian reef crisis (Flügel and Kiessling 2002) does not show any impact. There seems to be a more pronounced origination rate during end-Cretaceous times in the Campanian-Maastrichtian, an observation also made for dasycladalean algae (Barattolo 2002). A little more diversified information is obtained, when splitting the total number of genera into the order Helioporacea and Alcyonaria separately. It shows that the representatives of the order Helioporacea displays a more or less continuous increase until the Albian, then remaining constant throughout the Upper Cretaceous. Representatives of the order Alcyonaria appear in the Cenomanian for the first time (genus Moltkia!) increasing during end-Cretaceous times, outcompeting the Helioporacea in the Maastrichtian. Due to the uncertain suprageneric position of the genus Carpathoporella Dragastan, and differing opinions with respect to the taxonomy of individual species and genera, however, this compilation and the conclusions drawn only have provisionary character.

Number of octocoral genera (Pennatulacea excluded) (red dots) during the Cretaceous, based on the dataset of Fig. 4 and Table 1. Orange rectangles refer to the genera of the order Helioporacea and the blue asteriks to the order Alcyonaria. The genus Carpathoporella Dragastan is included in the undifferentiated data set (red dots), but excluded from the graphs of the Helioporacea and Alcyonaria, as its suprageneric taxonomic position is not verified (further explanations in the text)
Conclusions
The taxon Carpathoporella occidentalis Dragastan (=Coptocampylodon fontis nom. nud.), widespread in Lower Cretaceous peri-Mediterranean near-reefal shallow-water carbonates, cannot be interpreted as algal debris but as skeletal elements of colonial bushy octocorals, namely calcitic sclerites, axis (or internodes) and basal holdfast structures. The original description of the incertae sedis Coptocampylodon Elliott was in all likelihood a taxonomic mixing. It was based on either isolated specimens for which a coprolite nature is most likely, or smaller solid specimens in thin-sections that could be debris of dasycladalean algae or octocoral sclerites. Therefore, the other later described representatives from thin-sections that altogether do not show any feature pointing to a coprolite nature, must belong to a new (morpho)genus (genera?) or should be included in other existing taxa.
Within the Octocorallia, Carpathoporella belongs either to the order Alcyonacea (when the grooved and bended cylindrical elements are interpreted as internodes) or to the order Helioporacea (when these elements are interpreted as branches). With this respect, Carpathoporella shows similarities to the genus Moltkia Steenstrup on the one side and Epiphaxum Lonsdale on the other. A possible primary aragonitic skeletal mineralogy, however, brings Carpathoporella close to Epiphaxum Lonsdale exhibiting similar cylindrical grooved branches. A direct synonymization at the current state of knowledge, however, would be premature needing supplementary data. In such a case, the stratigraphic range of one of the aforementioned taxa must be enlarged into the Lowermost Cretaceous. For the time being, Carpathoporella Dragastan is treated as a separate genus including the two species C. occidentalis Dragastan and C. elliotti (Radoičić) nov. comb. The stratigraphy of Carpathoporella Dragastan is Berriasian?/Hauterivian to Albian/?Cenomanian. If an octocoral interpretation would also account for Coptocampylodon? rhaeticus Schlagintweit, Missoni and Gawlick, a taxon also possessing a hollow axis with outer grooves, this would be the first octocoral record from the Triassic.
The exclusive record of Carpathoporella Dragastan from Tethyan Lower Cretaceous shallow-water carbonates is of special importance since octocorals are well recorded from the boreal realm, preferentially from Upper Cretaceous or Cenozoic outer shelf environments; from Lower Cretaceous strata no skeletal elements similar or identical to Carpathoporella are known from deep-water deposits. It documents that octocorals were conspicuous inhabitants of Lower Cretaceous peri-reefal shallow-water benthic palaeocommunities of the Tethyan realm. A possible origin of Upper Cretaceous deep-water octocorals from Lower Cretaceous shallow-water ancestors in an onshore–offshore evolutionary pattern is here put up for further discussion. In any case, a better cooperation amongst researchers dealing with Recent octocorals and palaeontologists is highly desirable to avoid two concurrent systematics and to bring more light onto the phylogenetic history of the octocorals.
References
Alexandrowicz SW (1977) Sclerites of octocorals from the Upper Cretaceous of eastern Poland. J Paleontol 41:193–202
Anderson K, Close L, Dewreede RE, Lynch BJ, Ormond C, Walker M (2006) Biomechanical properties and holdfast morphology of coenocytic algae (Halimedales, Chlorophyta) in Bocas del Toro, Panama. J Exp Mar Biol Ecol 328:155–167. doi:10.1016/j.jembe.2005.07.005
Andrews AH, Cordes EE, Mahoney MM, Munk K, Coale KH, Cailliet GM, Heifetz J (2002) Age, growth and radiometric age validation of a deep-sea, habitat-forming gorgonian (Primnoa resedaeformis) from the Gulf of Alaska. Hydrobiologia 471:101–110. doi:10.1023/A:1016501320206
Andrews AH, Cailliet GM, Kerr LA, Kenneth HC, Lundstrom C, DeVogelaere AP (2005) Investigations of age and growth for three deep-sea corals from the Davidson seamount off central California. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, Berlin Heidelberg New York, pp 1021–1038
Auer M, Suzuki S, Schlagintweit F, Gawlick HJ (2008) Spatial and temporal development of siliceous basin and shallow-water carbonate sedimentation in Oxfordian Northern Calcareous Alps. Facies 55:63–87. doi:10.1007/s10347-008-0155-3
Ausich WI, Babcock LE (1998) The phylogenetic position of Echmatocrinus brachiatus, a probable octocoral from the Burgess Shale. Palaeontology 41:193–202
Ausich WI, Babcock LE (2000) Echmatocrinus, a Burgess Shale animal reconsidered. Lethaia 33:92–94. doi:10.1080/00241160050150221
Barattolo F (1984) Osservazioni su Suppiluliumaella schroederi n. sp. (alghe verdi, Dasicladali) del Cenomaniano del Matese (Appennino meridionale, Italia). Boll Soc Natur Napoli 92:1–47
Barattolo F (2002) Late Cretaceous-Paleogene dasycladaceans and the K/T boundary problem. In: Bucur II, Filipescu S (eds) Research advances in calcareous algae and microbial carbonates. Proceedings of the 4th IFAA regional meeting, Cluj-Napoca, Romania, 29 August–5 September 2001, pp 17–40
Basson PW, Edgell HS (1971) Calcareous algae from the Jurassic and Cretaceous of Lebanon. Micropaleontology 17:411–433. doi:10.2307/1484871
Bayer FM (1955a) Remarkably preserved fossil sea-pens and their recent counterparts. J Wash Acad Sci 45:294–300
Bayer FM (1955b) Contribution to the nomenclature, systematics, and morphology of the Octocorallia. Proc US Nat Mus 105:207–220
Bayer FM (1956) Octocorallia. In: Moore RC (ed) Treatise on invertebrate paleontology part F Coelenterata. Geol Soc Am Univ Kansas Press, Lawrence, pp 163–231
Bayer FM (1961) The shallow water Octocorallia of the west Indian region. Stud Fauna Curacao 12:1–373
Bayer FM (1979) The correct name of the helioporacean octocoral Lithotelesto micropora Bayer and Muzik. Proc Biol Soc Wash 92:873–875
Bayer FM (1981a) Recent advances in research on octocorals. Seminários Biologia Marinha, Academia Brasileira de Ciencias, pp. 19–28
Bayer FM (1981b) Key to the genera of Octocorallia exclusive of Pennatulacea (Coelenterata: Anthozoa), with diagnoses of new taxa. Proc Biol Soc Wash 94:902–947
Bayer FM (1990) A new isidid octocoral (Anthozoa: Gorgonacea) from New Caledonia, with descriptions of other new species from elsewhere in the Pacific Ocean. Proc Biol Soc Wash 103:205–228
Bayer FM (1992) The helioporacean octocoral Epiphaxum, recent and fossil. A monographic iconography. Stud Trop Oceanogr 15:1–76
Bayer FM, Deichmann E (1960) The Ellisellidae (Octocorallia) and their bearing on the zoogeography of the eastern Pacific. Proc Biol Soc Wash 73:175–182
Bayer FM, Macintyre IG (2001) The mineral component of the axis and holdfast of some gorgonacean octocorals (Coelenterata: Anthozoa) with special reference to the family Gorgonidae. Proc Biol Soc Wash 114:309–345
Bayer FM, Grasshoff M, Verseveldt J (1983) Illustrated trilingual glossary of morphological and anatomical terms applied to Octocorallia. E.J. Brill/Dr. W. Backhuys, Leiden, p 75
Bengtson S (1981) Atractosella, a Silurian alcyonacean octocoral. J Paleontol 55:281–294
Bernecker M, Weidlich O (2005) Azooxanthellate corals in the late Maastrichtian—early Paleocene of the Danish Basin: bryozoan and coral mounds in a boreal shelf setting. In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, Berlin Heidelberg New York, pp 3–25
Bernecker M, Weidlich O (2006) Paleocene bryozoan and coral mounds of Fakse, Denmark: habitat preferences of isidid octocorals. Courier Forsch Senckenberg 257:7–20
Bjerager M, Surlyk F (2007) Danian cool-water bryozoan mounds at Stevns Klint, Denmark—a new class of non-cemented skeletal mounds. J Sediment Res 77:634–660. doi:10.2110/jsr.2007.064
Borza K, Misík M (1976) Pienina oblonga n. gen., n. sp. aus kretazischen und paläogenen Kalken der Westkarpaten. Geol Zborn Slov akad vied 27:337–344
Bromley RG, Heinberg C (2006) Attachment strategies of organisms on hard substrates: a palaeontological view. Palaeogeogr Palaeoclimatol Palaeoecol 232:429–453. doi:10.1016/j.palaeo.2005.07.007
Brönnimann P (1976) Revision of the lectotype of Favreina salevensis (Paréjas) (Crustacea, Decapoda) and description of favreine form-species from the Jurassic and Cretaceous of Scotland, Portugal, Yugoslavia and Pakistan. Palaontol Z 50:40–56
Bucur II (1999) Stratigraphic significance of some skeletal algae (Dasycladales, Caulerpales) of the Phanerozoic. In: Farinacci A, Lord AR (eds) Depositional episodes and bioevents. Palaeopelagos Spec Publ 2:53–104
Bucur II, Koch R, Kirmaci ZM, Tasli K (2000) Les algues Dasycladales du Crétacé inférieur (Calcaire de Berdiga) de Kircaova (région de Kale-Gümüshane, NE Turquie). Rev Paleobiol 19:435–463
Cairns SD, Baco A (2007) Review and five new Alaskan species of the deep-water octocoral Narella (Octocorallia: Primnoidae). Syst Biodivers 5:391–407. doi:10.1017/S1477200007002472
Cairns SD, Bayer FM (2005) A review of the genus Primnoa (Octocorallia: Gorgonacea: Primnoidae), with the description of two new species. Bull Mar Sci 77:225–256
Cairns SD, Bayer FM (2009) Phylogenetic analysis of the Primnoidae (Cnidaria: Octocorallia). Smithsonian Contr Zool 269:1–79
Cherchi A, Schroeder R (1999) Montseciella, a new orbitolinid genus (Foraminiferida) from the uppermost Hauterivian—early Barremian of SW Europe. Treballs Mus Geol Barc 8:5–23
Colgan MW (1984) The Cretaceous coral Heliopora (Octocorallia: Coenothecalia)—a common Indo-Pacific reef builder. In: Eldredge N, Stanley SM (eds) Living fossils. Casebooks in earth sciences. Springer, Berlin Heidelberg New York, pp 266–271
Cope JCW (2005) Octocorallian and hydroid fossils from the Lower Ordovician of Wales. Palaeontology 48:433–445. doi:10.1111/j.1475-4983.2005.00455.x
Cuvillier J, Bassoullet JP, Fourcade E (1969) Coprolithes du Jurassique et du Crétacé d`Espagne et de quelques autres régions. Rev Micropal 11:183–190
Daly M, Brugler MR, Cartwright C, Collins AG, Dawson MN, Fautin DG, France SR, McFadden CS, Opresko DM, Rodriguez E, Romano SL, Stake JL (2007) The phylum Cnidaria: a review of phylogenetic patterns and diversity 300 years after Linnaeus. In: Zhang ZQ, Shear WA (eds) Linnaeus tercentenary: progress in invertebrate taxonomy. Zootaxa 1668:127–182
Deflandre-Rigaud M (1955) Sue les sclérites d’Alcyonaires fossiles et leur classification. Micralcyonarites manip. nov. du Miocène moyen d’Australie. C R Acad Sci 241:1327–1329
Deflandre-Rigaud M (1956) Les sclérites d’alcyonaires fossiles. Élèments d’ une classification. Ann Paleontol 42:1–24
Deflandre-Rigaud M (1957) A classification of fossil alcyonarian sclérites. Micropaleontology 3:357–366. doi:10.2307/1484441
Deloffre R (1988) Nouvelle taxonomie des algues Dasycladales. Bull Cent Rech Explor Prod Elf-Aquitaine 12:165–217
DeVictor ST, Morton SL (2007) Guide to the shallow water (0–200 m) octocorals of the South Atlantic Bight. Online guide. http://www.dnr.sc.gov/marine/sertc/octocoral%20guide
Dragastan O (1967) Alge calcaroase du Jurassique superior si Cretacicul inferior din Muntii Apuseni. St si cerc Geol Geofiz geogr Ser geol 12:441–454
Dragastan O (1989) Calcareous algae (new and revised), microproblematicae and Foraminiferida of Jurassic-Lower Cretaceous deposits from the Carpathian area. Rev Esp Micropal 21:5–65
Dragastan O (1995) Typification of some fossil algae and megaflora taxa from the collection of Professor Dr. Ovidiu Dragastan, University of Bucharest, Laboratory of Paleontology. Anal Univ Bucuresti Geol XLIV:105–107
Duncan PM (1879) On the Upper Greensand coral fauna of Haldon, Devonshire. Quat J Geol Soc 35:89–97
Eguchi J (1948) Fossil Helioporidae from Japan and the South Sea Islands. J Palaeont 22:362–364
Elliott GF (1963) Problematic microfossils from the Cretaceous and Paleocene of the Middle East. Palaeontology 6:293–300
Etnoyer PJ (2008) A new species of Isidella bamboo coral (Octocorallia: Alcyonacea: Isididae) from northeast Pacific seamounts. Proc Biol Soc Wash 121:541–553. doi:10.2988/08-16.1
Fabricius K, Alderslade P (2001) Soft corals and sea fans: a comprehensive guide to the tropical shallow-water genera of the Central West Pacific, the Indian Ocean and the Red Sea. Australian Inst Mar Sci, pp 1–244
Flügel E, Kiessling W (2002) Patterns of Phanerozoic reef crisis. In: Kiessling W, Flügel E, Golonka J (eds) Phanerozoic reef patterns. SEPM Spec Publ 72, pp 691–733
France SC (2007) Genetic analysis of bamboo corals (Cnidaria: Octocorallia: Isididae): does lack of colony branching distinguish Lepidisis from Keratoisis? Bull Mar Sci 81:323–333
Franzén C (1977) Crinoidal holdfasts from the Silurian of Gotland. Lethaia 10:219–234. doi:10.1111/j.1502-3931.1977.tb00617.x
Frech F (1890) Die Korallenfauna der Trias, die Korallen der juvavischen Triasprovinz (Zlambachschichten, Hallstätter Kalk, Rhaet). Palaeontogr 37:1–116
Funaki M, Nishizawa M, Sawaya T, Inoue S, Yamagishi T (2001) Mineral composition in the holdfast of three brown algae of the genus Laminaria. Fish Sci 67:295–300. doi:10.1046/j.1444-2906.2001.00236.x
Gawlick HJ, Frisch W, Hoxha L, Dumitrica P, Krystyn L, Lein R, Missoni S, Schlagintweit F (2008) Mirdita Zone ophiolites and associated sediments in Albania reveal Neotethys Ocean origin. Int J Earth Sci 97:865–881. doi:10.1007/s00531-007-0193z doi:10.1007/s00531-007-0193-z
Giammona CP, Stanton RJ Jr (1980) Octocorals from the Middle Eocene stone city formation, Texas. J Paleontol 54:71–80
Goldfuss GA (1826) Petrefacta Germaniae 1(1), Düsseldorf, pp 76
Granier B (1988) Les Pienina oblonga Borza et Mišík, 1976 sont-elles des sclérites d’Alcyonaires? Rev Micropal 29:103–108
Granier B, Deloffre R (1993) Inventaire critique des algues Dasycladales fossils–II Partie—Les algues Dasycladales du Jurassique et du Crétacé. Rev Paleobiol 12:19–65
Grasshoff M, Bargibant G (2001) Coral reef gorgonians of New Caledonia. IRD éditions Paris, 355 pp
Grasshoff M, Zibrowius H (1983) Kalkkrusten und Achsen von Hornkorallen, rezent und fossil. Senckenb Marit 15:111–145
Haas O (1909) Bericht über neue Aufsammlungen in den Zlambachmergeln der Fischerwiese bei Alt-Ausee. Beitr Palaontol Geol Osterr-Ung Orients 22:143–167
Hawksworth DL, McNeill J, Sneath PHA, Trehane RP, Tubbs PK (eds) (1994) Towards a harmonized bionomenclature for life on earth. Biol Int Spec Issue 30:1–43
Helm C, Schülke I (2003) An almost complete specimen of the Late Cretaceous (Campanian) octocoral “Isis” ramosa Voigt (Gorgonacea) from the Lower Saxony Basin, northwest Germany. Cretac Res 24:35–40. doi:10.1016/S0195-6671(03)00020-X
Hosseini SA, Conrad MA (2008) Calcareous algae, foraminifera and sequence stratigraphy of the Fahliyan Formation at Kuh-e-Surmeh (Zagros Basin, SW of Iran). Geol Croat 61:215–237
Iba Y, Sano SI (2007) Mid-Cretaceous step-wise demise of the carbonate platform biota in the Northwest Pacific and establishment of the North Pacific biotic province. Palaeogeogr Palaeoclimatol Palaeoecol 245:462–482. doi:10.1016/j.palaeo.2006.09.008
Jablonski D, Sepkoski JJ, Bottjer DJ, Sheehan PM (1983) Onshore–offshore patterns in the evolution of Phanerozoic shelf communities. Science 222:1123–1125. doi:10.1126/science.222.4628.1123
Jacobs DK, Lindberg DR (1998) Oxygen and evolutionary patterns in the sea: onshore/offshore trends and recent recruitment of deep-sea fauna. Proc Natl Acad Sci USA 95:9396–9401. doi:10.1073/pnas.95.16.9396
Janes MP, Wah LM (2007) Octocoral taxonomy laboratory manual. Results of the international workshop on the taxonomy of Octocorals, University of Kerala, India, 20–26 March 2005, p 91
Jeletzky JA, Zapfe H (1967) Celoid and orthocerid cephalopods of the Rhaetian Zlambach Marls from the Fischerwiese near Ausee, Styria (Austria). Ann Naturhist Mus 71:69–106
Kiessling W, Baron-Szabo R (2004) Extinction and recovery pattern of scleractinian corals at the Cretaceous–Tertiary boundary. Palaeogeogr Palaeoclimatol Palaeoecol 214:195–223
Kocurko MJ (1988) Notes on fossil octocorals and comparisons of some modern and ancient octocoral remains. Tulane Stud Geol Paleontol 21:105–115
Kocurko MJ (1993) Eunicella sp., octocorallia from the Red Bluff Formation, Lower Oligocene, Mississippi. Tulane Stud Geol Paleontol 26:35–40
Kocurko MJ, Kocurko DJ (1992) Fossil octocorallia of the Red Bluff Formation, Lower Oligocene, Mississippi. J Palaeontol 66:594–602
König W (1991) Oktokorallen-Basen aus dem Campanium von Misburg und Höver. Arbeitskr Palaontol Hannover 19:153–176
Kuzmicheva EI (1970) Pervyye nakhodki rannemelovykh korallov Heliopora Blainv. i Polytremacis Orb. na territorii SSSR i sistematicheskoye polozheniye etikh rodov (First discoveries of Early Cretaceous corals belonging to Heliopora and Polytremacis in the USSR and the systematic position of these genera). In: Mezozoyskiye korally SSSR (Mesozoic corals of the USSR), Papers of II allunion symposium on fossil corals of the USSR 4:93–100
Kuzmicheva EI (1975) On the taxonomic position and development of the family Helioporidae (in Russian). Paleontol Zhumal 3:11–17
Kuzmicheva EI (1980) Fossil Gorgonida. Paleontol J 14:1–12
Kuzmicheva EI (1982) Korally verkhnego Apta (Klanseya) tsentralnykh Kyzylkumov. Byulleten Moskovskogo Obshchestva Ispytateley Prirody Otdel Geologicheskiy 57:98–111. Upper Aptian (Clansayesian) corals from central Kyzylkum
Kuzmicheva EI (1987) Vershnemelovie i paleogeovie koralii SSSR (Upper Cretaceous and Paleogene corals of the USSR). Akad Nauk SSR, Moscow, p 187
Langer M (1989) Haftorgan, Internodien und Sklerite von Keratoisis melitensis (Goldfuss, 1826) (Octocorallia) in pliozänen Foraminiferenmergeln (Trubi) von Milazzo (Sizilien). Palaontol Z 63:15–24
Lehmann JGC (1817) Beschreibung einiger neuer und wenig bekannter Pflanzen. Neue Schr Naturfosch Ges Halle 3:1–26
Lehner L (1937) Fauna und Flora der Fränkischen Albüberdeckenden Kreide. II. Fauna 2. Teil und Flora. Palaeontogr LXXXVII:158–230
Leloux J (1999) Numerical distribution of Santonian to Danian corals (Scleractinia, Octocorallia) of southern Limburg, the Netherlands. Geol Mijnb 78:191–195. doi:10.1023/A:1003743301625
Lewis JC, Von Wallis E (1991) The function of surface sclerites in gorgonians (Coelenterata, Octocorallia). Biol Bull 181:275–288. doi:10.2307/1542099
Lewis JC, Barnowski TF, Telesnick GJ (1992) Characteristics of carbonates of gorgonian axes (Coelenterata, Octocorallia). Biol Bull 183:278–296. doi:10.2307/1542215
Lindner A, Cairns SD, Cunningham CW (2008) From offshore to onshore: multiple origins of shallow-water corals from deep-sea ancestors. PLoS One 3(6):e2429. doi:10.1371/journal.pone.0002429
Lindström M (1978) An octocoral from the Lower Ordovician of Sweden. Geol Palaeontol 12:41–52
Ljubović-Obradović D, Radoičić R (2003) Coptocampylodon pantici n. morpho sp. from the Turonian of NW Serbia. Stud Univ Babes-Bolyai Geol XLVIII:133–139
Lonsdale W (1850) Notes on the corals. In: Dixon F (ed) The geology and fossils of the Tertiary and Cretaceous formations of Sussex. Description of the fossils of the chalk formation. Brown, Green and Longmans, London, p 422
Löser H (1993) Morphologie und Taxonomie der Gattung Pseudopolytremacis Morycowa, 1971 (Octocorallia; Kreide). In: Oekentorp-Kuester P (ed) Proceedings of the VI international symposium on fossil Cnidaria and Porifera held in Münster, Germany, 9–14 September 1991. Courier Forschinst Senckenberg 164:211–220
Löser H (1996) A new octocoral from the Upper Cretaceous of east Bavaria. N Jb Geol Paläont Mh 8:485–489
Löser H (2000) Repertoire of species. Catalogue of Cretaceous corals 2, vol 2. Cpress, Dresden, p 784
Löser H (2003) Internodien der Gattung Moltkia (Octocorallia) aus einem Maastricht-Geschiebe (Oberkreide) von Niedersachsen (Deutschland). Berl Beitr Geschiebeforsch 2:99–101
Löser H (2005) Stratigraphy of Cretaceous coral genera. N Jb Geol Paläont Abh 238:231–277
Löser H, Ferry S (2006) Coreaux du Barrémien du Sud de la France (Ardèche et Drôme). Geobios 39:469–489. doi:10.1016/j.geobios.2005.03.005
Lozouet P, Molodtsova T (2008) Filling a gap: the first occurrences of Epiphaxum (Cnidaria: Helioporacea: Lithotelestidae) in the Eocene, Oligocene and Miocene. Palaeontol 51:241–250
Macintyre IG, Bayer FM, Logan MAV, Skinner HCW (2000) Possible vestige of early phosphatic biomineralization in gorgonian octocorals (Coelenterata). Geology 28:455–458. doi:10.1130/0091-7613(2000)28<455:PVOEPB>2.0.CO;2
Majoran S (1987) Structural investigation of octocoral sclerites. Zool Scr 16:277–287. doi:10.1111/j.1463-6409.1987.tb00074.x
Malecki J (1982) Bases of Upper Cretaceous octocorals from Poland. Acta Paleont Polon 27:65–75
Masse JP, Arnaud-Vanneau A (1999) Early Cretaceous calcareous algae of the Mid-Pacific Mountains. Rev Micropal 42:57–69. doi:10.1016/S0035-1598(99)90186-1
Milne Edwards H, Haime J (1851) A monograph of the British fossil corals, 1. Introduction; tertiary and cretaceous. Paleont Soc Monogr 3:1–71
Mišík M (1998) Pienina oblonga—skeletal parts or endoparasites of Keratosa sponges? Geol Carpath 49:401–407
Nielsen KB (1913) “Moltkia isis Steenstr.” og andere Octocorallia fra Danmarks kridttidsaflejringer. Mindeskrift Jap Steenstrup:1–19
Nielsen KB (1917) Heliopora incrustans nov. sp. with a survey of the Octocorallia in the deposits of the Danian in Denmark. Bull Geol Soc Den 5:1–13
Nielsen KB (1925) Nogle nye Octocoraller fra Danienet. Medd Danks geol Forening 6:117–126
Nielsen JK, Jacobsen SL (2004) Extraction of calcareous macrofossils from the Upper Cretaceous white chalk and other sedimentary carbonates in Denmark and Sweden: the acid-hot water method and the waterblasting technique. Palaeontol Electron 7:1–11
Noé S, Dullo WC (2006) Skeletal morphogenesis and growth mode of modern and fossil deep-water isidid gorgonians (Octocorallia) in the West Pacific (New Zealand and Sea of Okhotsk). Coral Reefs 25:303–320. doi:10.1007/s00338-006-0095-8
Noé S, Lembke-Jene L, Reveillaud J, Freiwald A (2007) Microstructure, growth banding and age determination of a primnoid gorgonian skeleton (Octocorallia) from the late Younger Dryas to earliest Holocene of the Bay of Biscay. Facies 53:177–188. doi:10.1007/s10347-007-0104-6
Patrulius D (1966) Coptocampylodon fontis n. sp., microfosil problematic al calcarelor Urgoniene din Muntii Persani. Dari de seama ale Sed LII/1 (1964–1965):391–394
Peza LH, Marku D (2002) Lower Cretaceous in the Munella Mountains (Mirdita Zone, northeastern Albania). Österr Akad Wiss. Schriftenr Erdwiss Komm 15:365–372
Radoičić R (1969) Coptocampylodon de certains sédiments jurassiques et crétacés de la Yougoslavia. Vesnik Zav geol geofiz istr 27:195–200
Radoičić R (2002) Dasycladalean algae from the Lower Cretaceous of Mt. Rujen (Vardar Zone, Macedonia). Rev Paleobiol 21:7–19
Radoičić R (2005) New Dasycladales and microbiota from the lowermost Valanginian of the Mirdita Zone. Ann Geol Penins Balk 66:27–53
Radwanska U, Radwanski A (2003) The Jurassic crinoid genus Cyclocrinus d’Orbigny, 1850: still an enigma. Acta Geol Polon 53:301–320
Rahman MA, Oomori T (2008) Structure, crystallization and mineral composition of sclerites in the alcyonarian coral. J Cryst Growth 310:3528–3534. doi:10.1016/j.jcrysgro.2008.04.056
Reich M (2001) Skleren von Oktokorallen aus einem Silur-Geschiebe Vorpommerns. Geschiebekunde Aktuell 16:59–61
Reich M (2002) Skleren von Alcyonacea (Anthozoa: Octocorallia) aus einem Silur-Geschiebe Norddeutschlands. N Jb Geol Pal Mh 9:551–561
Reich M (2007) Where are all the fossil octocorals (Cnidaria: Anthozoa)? In: Elicki O, Schneider JW (eds) Fossile Ökosysteme 77. Jahrestagung Pal Ges Abstracts, TU Freiberg Inst Geol Wiss Mitt 36:107
Reich M, Schneider S (2002) Erster Nachweis einer Seefeder (Octocorallia: Pennatulacea) aus dem Sternberger Gestein (Oligozän). Geschiebekunde aktuell 18:3–8
Sánchez J, Tracey D, Neil H, Marriott P (2004) Coral rings in the deep ocean: using SEM to date New Zealand’s bamboo corals. Water Atmos 12:22–23
Schindler U, Conrad MA (1994) The Lower Cretaceous Dasycladales from the Northwestern Friuli platform and their distribution in chronostratigraphic and cyclostratigraphic units. Rev Paleobiol 13:59–98
Schlagintweit F, Gawlick HJ (2007) Analysis of Late Jurassic to Early Cretaceous algal debris-facies of the Plassen carbonate platform in the Northern Calcareous Alps (Germany, Austria) and in the Kurbnesh area of the Mirdita zone (Albania)—a tool to reconstruct tectonics and paleogeography of eroded. Facies 53:209–227. doi:10.1007/s10347-006-0100-2
Schlagintweit F, Missoni S, Gawlick HJ (2002) Coptocampylodon? rhaeticus n. sp., a new problematic microfossil from the Rhaetian Dachstein Limestone of the Northern Calcareous Alps (Germany, Austria). Geol Croat 55:1–14
Schlagintweit F, Gawlick HJ, Missoni S, Hoxha L, Lein R, Frisch W (2008) The eroded Late Jurassic Kurbnesh carbonate platform in the Mirdita Ophiolite Zone of Albania and its bearing on the Jurassic orogeny of the Neotethys realm. Swiss J Geosci 101:125–138. doi:10.1007/s00015-008-1254-4
Schmid DU (1996) Marine Mikrobolithe und Mikroinkrustierer aus dem Oberjura. Profil 9:101–251
Schroeder R, Clavel B, Cherchi A, Busnardo R, Charollais J, Decrouez D (2002) Lignées phylétiques d’Orbitolinidés de l’intervalle Hauterivian supérieur–Aptien inférieur; leur importance stratigraphique. Rev Paleobiol 21:853–863
Sepkoski J (2002) A compendium of fossil marine animal genera. Bull Am Paleontol 363:1–560
Sethmann I, Helbig U, Wörheide G (2007) Octocoral sclerite ultrastructures and experimental approach to underlying biomineralisation principles. Cryst Eng Comm 9:1262–1268. doi:10.1039/b711068e
Sherwood OA, Heikoop JM, Sinclair DJ, Scott DB, Risk MJ, Shearer O, Azetsu-Scott A (2006) Skeletal Mg/Ca in Primnoa resedaeformis: relationship to temperature? In: Freiwald A, Roberts JM (eds) Cold-water corals and ecosystems. Springer, Berlin Heidelberg New York, pp 1061–1079
Sokač B, Nikler L (1971) Cymopolia longistila n. sp., a new calcareous alga (Dasycladaceae) from the Lower Cretaceous of the Dinaric Mountains. Geol Vjesn 24:77–84
Sokač B, Nikler L (1973) Calcareous algae from the Lower Cretaceous of the environs of Nikšić, Crna Gora (Montenegro). Palaeont Jugosl 13:7–57
Squires RL (1999) Upper Paleocene to Lower Eocene (“Meganos stage”) marine megafossils in the uppermost Santa Susana Formation, Simi Valley, Southern California. Contrib Sci 479:1–38
Stanley SM, Ries JB, Hardie LA (2002) Low-magnesium calcite produced by coralline algae in seawater of Late Cretaceous composition. Proc Natl Acad Sci USA 99:15323–15326. doi:10.1073/pnas.232569499
Steenstrup J (1846) Ueber Korallen in der Faxöer Kreide, Moltkia und Cyathidium. Amtl Ber 24. Vers dtsch Naturforsch Aerzte Sept 1846:148–150
Tomás S, Löser H, Salas R (2008) Low-light and nutrient-rich coral assemblage in an Upper Aptian carbonate platform of the southern Maestrat Basin (Iberian Chain, eastern Spain). Cretac Res 29:509–534. doi:10.1016/j.cretres.2007.09.001
Turnšek D (1997) Mesozoic corals of Slovenia. Zbirca ZRC Ser 16:1–512
Van Oppen MJ, Mieog JC, Sanchez CA, Fabricius KE (2005) Diversity of algal endosymbionts (zooaxanthellae) in octocorals: the roles of geography and host relationships. Mol Ecol 14:2403–2417. doi:10.1111/j.1365-294X.2005.02545.x
Velić I (2007) Stratigraphy and palaeobiogeography of Mesozoic benthic foraminifera of the Karst Dinarides (SE Europe). Geol Croat 60:1–114
Voigt E (1958) Untersuchungen an Oktokorallen aus der oberen Kreide. Mitt Geol Pal Inst Univ Hamburg 27:5–49
Weinbauer MG, Vellmirov B (1995) Calcium, magnesium and strontium concentrations in the calcite sclerites of Mediterranean gorgonians (Coelenterata: Octocorallia). East Coast Shelf Sci 40:87–104. doi:10.1016/0272-7714(95)90015-2
Wells JW (1932) Corals of the Trinity Group of the Comanchean of central Texas. J Paleontol 6:225–256
Williams GC (2003) Capitate taxa of the soft coral genus Eleutherobia (Octocorallia: Alcyoniidae) from Palau ad South Africa; a new species and a new combination. Zool Verhand Leiden 345:419–436
Williams GC, Cairns D (2006) Systematic list of valid octocoral genera. Published online. http://research.calacademy.org/research/izg/OCTOCLASS.htm#penna
Wing BL, Barnard DR (2004) A field guide to Alaskan corals. NOAA Tech Mem NMFS-AFSC-146, US Dept of Commerce, pp 1–68
Wittler FA (1998) Eine Wurzel von Bourgueticrinus d’Orbigny (Crinoida) aus dem Campan von Coesfeld (westl. Münsterland) epizoisch auf Echinocorys humilis. Arbeitskr Palaontol Hannover 26:84–96
Wittler FA (2001a) Zur systematischen Position der “Oktokorallenbasis” Octobasis spinosa Malecki 1982 aus der Oberkreide. Arbeitskr Palaontol Hannover 29:16–18
Wittler FA (2001b) Ein Nachweis von Epiphaxum auloporoides Lonsdale, 1850 im Oberturon von Wüllen bei Ahaus (Octocorallia, Oberkreide, NW-Deutschland). Dortmunder Beitr Landeskunde 35:237–246
Wright EP (1869) On a new genus of Gorgonidae from Portugal. Ann Mag Nat. Hist (Paris) 3:23–36
Xhomo A, Kodra A, Dimo L et al (2002) Geological Map of Albania 1:200,000 scale. Geological Survey of Albania, Republika e Shqiperise
Źìtt J, Nekvasilová O (1993) Octocoral encrusters of rock substrates in the Upper Cretaceous of Bohemia. J Czech Geol Soc 38:71–78
Živkovic S, Bogner D (2006) Coprolite status of Coptocampylodon lineolatus Elliott 1963 (incertae sedis) from Middle Eocene deep-sea sediments of Istra (Croatia). Micropal 52:371–379. doi:10.2113/gsmicropal.52.4.371
Acknowledgments
Common field work in the Munella area was carried out together with Lirim Hoxha (Geological Survey, Tirana) and Tonin Deda (Geological Institute, Tirana), who introduced the geology of Munella Mountains in the framework of the joint project “Tectonothermal Evolution of Albanides 2003–2005”. For discussions in the field many thanks to: Wolfgang Frisch (Tübingen, Germany), Richard Lein (Vienna, Austria), Sigrid Missoni (Leoben, Austria) additionally for many other technical help. Ovidiu Dragastan (Bucharest, Romania), Hannes Löser (Munich, Germany) and André Freiwald (Erlangen, Germany) are thanked for helpful comments. We kindly acknowledge the remarks of the reviewers Michaela Bernecker (Erlangen, Germany) and Ioan Bucur (Cluj-Napoca, Romania).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Schlagintweit, F., Gawlick, HJ. The incertae sedis Carpathoporella Dragastan, 1995, from the Lower Cretaceous of Albania: skeletal elements (sclerites, internodes/branches, holdfasts) of colonial octocorals. Facies 55, 553–573 (2009). https://doi.org/10.1007/s10347-009-0185-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-009-0185-5