The new crustacean microcoprolite Broennimannia persica n. ichnogen., n. ichnosp., is described from the Paleocene (Danian-Selandian) of eastern Iran. The new microcoprolite is characterized by a bilateral symmetrical system of longitudinal “canals” that in transverse sections display an irregular (labyrinthine) outline with concave infoldings and rounded protuberances. The crescentic outline of parts of the canal system recalls the long-ranging ichnogenus Palaxius Brönnimann & Norton (Carboniferous to Pleistocene). In the latter ichnogenus, however, the canals are isolated. A general determination key for microcoprolites in transverse section is provided. The new microcoprolite occurs in poorly sorted foram-algal grainstones/packstones with a fine-grained siliciclastic content. The associated microfauna and –flora includes common miliolids, among the agglutinating Haymanella, Stomatorbina sp., serpulids, and dasycladalean algae (Cymopolia sp., Trinocladus atacis Segonzac). From Iran, fossil microcoprolites have been described so far only from the Upper Jurassic, Lower Cretaceous, and Miocene. Broennimannia persica represents the first Paleocene record from this area.
Internally structured crustacean microcoprolites are common in the geological record and have been described ichnotaxonomically from the Palaeozoic (Devonian; Herbig 1993) to Recent (see overview in Knaust 2020). For example, they are widely reported from Cretaceous shallow-water carbonates (e.g., Brönnimann and Masse 1969; Vialov 1978; Senowbari-Daryan and Grötsch 1992; Senowbari-Daryan and Kuss 1992; Fenninger and Hubmann 1994; Blau and Grün 1997; Senowbari-Daryan et al. 2009; Kietzmann and Palma 2014). However, there are few reports from the Paleocene (e.g., Senowbari-Daryan and Kube 2003: Palaxius; Buchs et al. 2009), Eocene (Brönnimann and Norton 1960; Gavrilov and Shchepetova 2000; Peckmann et al. 2007: Palaxius), Oligocene (Altini 1942; Paréjas 1948: Palaxius), Miocene (Brönnimann and Norton 1960: Palaxius; Ellliott 1962: Favreina; Schweigert et al. 1997: Helicerina), or the Pleistocene (Gischler et al. 2017: Palaxius). The classification of these ichnogenera and ichnospecies is based on the arrangement (pattern) and number of the longitudinal canals in transverse sections (e.g., Brönnimann 1972; Knaust 2020). As known from modern crabs, these result from a complex system of folds in the wall of the digestive tract where the ingested food is circulating (e.g., Powell 1974; Dall and Moriarty 1983). These folds may be irregular, longitudinal, with and without spines (Moon and Kim 1999), thereby accounting for the variety of different canal patterns reported in the fossil record of microcoprolites.
From Iran, there are some records of crustacean microcoprolites in the literature. Brönnimann (1977) described Favreina tabasensis from the Upper Jurassic of the Tabas area of central Iran. From oil–wells drilled in the Persian Gulf, crustacean microcoprolites were recently reported from the Valanginian-Hauterivian Gadvan Formation (Favreina iranensis), the Aptian Dariyan Formation (Palaxius minaensis), and the Miocene Asmari Formation (Palaxius asmariensis) (Dalvand et al. 2015). Other Middle Jurassic to Lower Cretaceous records are from the Zagros Zone, SW Iran (Rostami and Ahmadi 2013; Motaharian et al. 2014).
The detailed micropalaeontological investigation of a section of Paleocene shallow-marine carbonates in eastern Iran yielded a new microcoprolite described here as Broennimannia persica n. ichnogen., n. ichnosp.
Geological setting
Overview
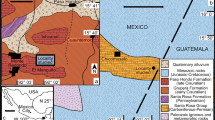
The structural geology of central Iran is characterized by a complex array of blocks and microplates (e.g., Stöcklin 1968) (Fig. 1a). The so-called Central-East Iranian Microcontinent (CEIM; Takin 1972), consists of the Yazd, Tabas, and Lut blocks (Berberian and King 1981; Camp and Griffis 1982; Tirrul et al. 1983). The Lut Block is separated from the East Iranian Range (or Sistan Suture Zone; Tirrul et al. 1983; Aghanabati 2004) in its middle and upper parts by the Nehbandan Fault. However, there is no sharp boundary in the upper part between these two geological zones (e.g., Zanchi et al. 2009; Fig. 1). The studied section is situated in this area and consists of Upper Cretaceous sediments, ophiolitic series, flysch-type sediments, and Paleogene rocks. The thickness of the Paleogene sediments varies considerably, even within short distances.
Fig. 1
Geological and geographic location of the Grong section in eastern Iran. a Approximate location plotted on the tectonic map of Iran (modified from Zanchi et al. 2009). AA, Anatolian–Armenian Block; AMC, Anarak Metamorphic Complex; KDF, Kopeh Dagh Foredeep, MZT, Main Zagros Trust). b Geographic location of the Grong section, the type–locality of Broennimannia persica ichngen. et ichnosp. nov
The studied section, here named the Grong section (after the eponymous mountain and village nearby), is located ~ 60 km west of Birjand town and 5 km north-west of Gheisar village in the Ching-dar syncline (Fig. 1b). The Greenwich coordinates of the section base are N 32° 52′ 27.3″ and E 58° 42′ 7.6″. It can be found on the geological map of the Kusf quadrangle 1:100,000 (Eftekhar-Nezad 1986). Lithologically it consists of lithostratigraphically undefined Upper Cretaceous and Paleogene sediments: conglomerates, sandstones, marls, sandy limestones and limestones (Fig. 2). The analyzed thin sandy limestone beds of samples M1-3, -5, and -12, yielded no microfossils. The assumed Upper Cretaceous age is based only on the geological map of Eftekhar-Nezad (1986), and its superposition by Paleogene sediments of the Palang Formation. The conglomerates separating this unit from a package of sandy, grey, massive limestones might correspond to the Cretaceous-Paleogene boundary. The new microcoprolite was observed in the basal part of this unit (samples M2-1, -2), together with indeterminant small rotaliids, and dasycladalean algae indicating a Paleocene age, Clypeina elliotti Beckmann & Beckmann and Trinocladus atacis Segonzac (see Barattolo 2002) (Fig. 3a, b). A lithologically similar facies, also containing Broennimannia persica ichnogen. et ichnosp. nov. at its base re-appears higher up in the section. Here the new ichnotaxon is associated with the agglutinating miliolid Haymanella gr. paleocenica Sirel (Fig. 3f), dasycladalean and halimedacean algae. This microfacies can be compared with the Haymanella-algal limestone reported by Sirel (1998, 1999) from Turkey, or Danian carbonates of the Yazd Block, central Iran (Schlagintweit and Rashidi 2019). As at its base, the Paleogene sediments are overlain by conglomerates at the top containing clasts of volcanic rocks and upper Maastrichtian carbonates containing fragments of the large benthic foraminifera Siderolites, Orbitoides, and Omphalocyclus.
Fig. 2
Stratigraphic log of the Grong section, showing lithostratigraphic units, position of samples and distribution of benthic foraminifera, calcareous algae and the microcoprolite Broennimannia persica ichnogen. et ichnosp. nov
Contrasting with other sections of Paleocene carbonates in Central Iran or the Zagros Zone of southwestern Iran (e.g., Rahaghi 1983; Schlagintweit and Rashidi 2019), the studied section is devoid of biostratigraphically important benthic foraminifera. The lack of typical large benthic foraminifera (see Serra-Kiel et al. 1998) is particularly striking. Cocoarota orali İnan has been observed in the first samples overlying the conglomerates at the base of the Paleogene sediments (Fig. 3c). This species ranges from the Maastrichtian to middle Eocene (İnan 2003). The rotaliid taxon Rotorbinella hensoni (Smout) (including the morphotype R. detrecta Hottinger) higher up in the section (Fig. 3d) indicates the shallow benthic zones (SBZ) 1 to 3 of Serra-Kiel et al. (1998), that is, late Danian to Thanetian (e.g., Hottinger 2014). The range of Miscellanites minutus (Rahaghi) (Fig. 3e) was originally restricted to SBZ 3 by Hottinger (2009) and extended to the SBZ 2 recently by Consorti and Köroğlu (2019). No benthic foraminifera typical for (or restricted to) the Thanetian (SBZ 3 pro parte) have been observed. This leads us to conclude that the Paleocene sediments of the Grong section can be assigned to the Danian-Selandian interval. The observed dasycladalean algae such as Dissocladella savitriae Pia (Fig. 3h, j) or Cymopolia drobnae Radoičić support this age (Barattolo 2002). The known range of Trinocladus atacis (Fig. 3i), Thanetian fide Barattolo (2002), must be extended.
Palaeoenvironment
Microcoprolites have been reported from diverse palaeoenvironments such as (i) the basal parts of transgressive series representing near-shore environments (e.g., Fenninger and Hubmann 1994), (ii) peritidal carbonates (tidal flats) and hostile hypersaline ponds with thrombolites (e.g., Rindsberg and Kopaska-Merkel 2013), (iii) reefal (here commonly in cavities; Senowbari-Daryan 1979) and near-reefal carbonates (e.g., Senowbari-Daryan et al. 1979), or (iv) hydrothermal vent sites in shallow (Bujtor 2011, 2012; Senowbari-Daryan et al. 2007) and deep marine settings (Buchs et al. 2009). Rather commonly, the palaeoenvironments where microcoprolites occur in great abundance are described as “hostile to most forms of life” (Rindsberg and Kopaska-Merkel 2013). Crustacean microcoprolites may also be reworked into storm deposits (Kietzmann and Palma 2014).
Concerning the occurrence of the Broennimannia persica, we note its presence in two distinct levels both displaying similar microfacies (Figs. 2, 3a, b). In these poorly sorted grainstones (with a fine–grained siliciclastic content), the microcoprolites are associated with dasycladalean algae, more frequent in the upper level and here together with the agglutinating miliolid Haymanella gr. paleocenica Sirel. The lower level is underlain by terrestrial conglomerates and corresponds to the basal part of a transgressive unit, thus reflecting a near-shore depositional setting. In addition, the second level corresponds to the base of a lithologic change marked by the siliciclastic influx and is interpreted as deposits of a very shallow, siliciclastic–influenced inner lagoon, evidenced also by the common occurrence of dasycladalean algae. The overall high-energy environment is documented by the grainstone texture.
Material and depository
The images of the new microcoprolite refer to five thin–sections deposited in the Geosciences Museum of Mashhad (in the Geological Survey of North-Iran East territory) (Table 1).
Table 1 Numbers of thin-sections (original field numbers) and official depository numbers of the Geosciences Museum of Mashhad (in the Geological Survey of North-Iran East territory)
Derivatio nominis. Dedicated to Paul Brönnimann for his numerous fundamental ichnotaxonomic works on crustacean microcoprolites.
Diagnosis. Rod-shaped coprolite, rounded to slightly oval in transverse section (in some cases with a small concave “dorsal” depression), displaying a bilaterally symmetric pattern of longitudinal canals with irregular constrictions and swellings of labyrinthine aspect. The upper (“dorsal”) canals are larger than the lower (“ventral”) ones. The concave infoldings of the canals face the surface of the microcoprolite. The microcoprolite is composed of homogenous micrite without a “ventral” cap.
Remarks. Fossil microcoprolites have different cross-sections of their canals arranged bilaterally symmetrically (Fig. 4). They may be of a different shape such as in Fundalutum (Fig. 4a) or they may display a homogeneous morphology: rounded (Favreina, Fig. 4e), Y-shaped within the plane of symmetry (Helicerina, Lercarina, Fig. 5c, d), sickle-shaped (Palaxius, Fig. 4g), or triangular (Parafavreina, Fig. 4f). Sulcusina displays a median canal (or furrow) and rounded canals like Favreina (Fig. 4b). Others display a light–grey micritic “ventral” cap (Payandea, Thoronetia, Fig. 4i, j). Broennimannia differs from all these morphotypes by its relatively large canals displaying irregular constrictions and swellings, giving it a labyrinthine aspect (Fig. 4h). It is noteworthy that Kietzmann and Palma (2014, p. 216) discussed the possibility that Fundalutum is not a new taxon, but might be “a poorly preserved specimen of Palaxius caucaensis in which narrower parts of the canals are not preserved“. The presence of concave depressions of the canals facing the microcoprolite surface in Broennimannia recalls the ichnogenus Palaxius. This aspect may especially be relevant in cases where the degree of preservation is low. In Palaxius, the crescentic- to hook-shaped canals are isolated (e.g., like a crescent moon), whereas in Broennimannia the canals display a complex laterally connected system with numerous concave infoldings or depressions. A general determination key for microcoprolites is presented in Fig. 5.
Fig. 4
Idealized cross sections (drawings) of microcoprolites (without scale). aFundalutum Senowbari-Daryan and Kuss (1992). bSulcusina Schlagintweit et al. (2016). cHelicerina Brönnimann and Masse (1969). dLercarina Senowbari-Daryan (1988). eFavreina Brönnimann. fParafavreina Brönnimann et al. (1972). gPalaxius Brönnimann and Norton (1960). hBroennimannia, this work. iPayandea Blau et al. (1993). jThoronetia Brönnimann et al. (1972). s.p., symmetry plane; v.c., ventral cap
General determination key for microcoprolites in transverse sections. For further details the reader is referred to the original diagnoses provided by the authors
Broennimannia persica ichngen. et ichnosp. nov. from the Paleocene Grong section of eastern Iran. a, d, g–i transverse sections. Holotype specimen in h. b–c, e, n slightly oblique transverse sections. f, j fragmentary, slightly oblique transverse sections. k–m oblique sections. o longitudinal section. Thin-sections: M2-2 (a), M3-12-2 (b–f, h, j, k, n), M3-12-3 (g, i, m)
Derivation nominis. For Persia, the historically common name for Iran.
Holotype. The specimen illustrated in Fig. 6h; thin–section Gmm 13980F25 (M3 12-2, see Fig. 2 for sample location). The diameter of the holotype is 0.87 mm.
Horizon and locality. Paleogene (Danian–Selandian) carbonates (lithostratigraphically not defined) of the Grong section located ~ 60 km west of Birjand town (Fig. 1b). The Greenwich coordinates of the section base are N 32°52′27.3″, E 58°42′7.6″.
Diagnosis. As for the ichnogenus.
Description. Broennimannia persica is a rod-like microcoprolite represented by different random sections. Transverse sections are rounded to slightly oval. In some specimens, a shallow concave depression can be observed on the “ventral” side. Some specimens display an elliptical outline (Fig. 6i) that might be due to compression. The microcoprolite itself is homogeneously micritic with a smooth surface and no further differentiation of the material or texture (e.g., “ventral cap”). There are four canals (or canal systems) arranged in bilateral symmetry (Fig. 6a, g–i). All these canals display an irregular labyrinthine outline with constrictions, concave depressions or infoldings and rounded protuberances. There are two pairs of canals on each side with identical morphology but different in outline (1–1′ versus 2–2′, Fig. 4h). The upper two canals have a larger diameter and display a straight and closed inner side parallel to the median plane. On the periphery, i.e. following the rounded margin of the microcoprolite, there are two (up to three?) concave depressions. One basal opening slightly shifted towards the direction of the median plane, is present (e.g., Fig. 6h). The base of the canal (perpendicular to the plane of symmetry) may be straight or obliquely arranged. The two smaller canals at the “ventral” side display one inner opening towards the median plane. In some specimens, the canal appears as displaying a continuous outline with constrictions (Fig. 6h), whereas in others it is split into several canals with an irregularly crescentic outline with short terminal swellings recalling the ichnogenus Palaxius (Fig. 6i).
Dimensions.
Diameter: 0.5–1.2 mm (mostly around 0.7 to 1.0 mm)
Length: up to 1.2 mm.
Concluding remarks
A new ichnotaxon of crustacean microcoprolite, Broennimannia persica n. ichnogen., n. ichnosp., is described from the Danian–Selandian shallow-water carbonates of eastern Iran. From Iran, fossil microcoprolites have been described so far from the Upper Jurassic, Lower Cretaceous and the Miocene, but not from the Paleocene. The new ichnogenus displays a rather complex, bilaterally symmetrical canal system. With its irregularly concave depressions, it shows similarities to Palaxius. Broennimannia persica occurs in poorly sorted grainstones with a fine-grained siliciclastic content of transgressive units. They reflect a near-shore, terrestrial influenced inner lagoonal depositional setting with common dasycladalean algae and porcelaneous benthic foraminifera.
References
Aghanabati A (2004) Geology of Iran. Geological survey of Iran, p 586
Altini E (1942) Étude géologique de la chaîne côtière entre Bandirma-Gemlik. Istanbul Univ Fen Fak Mecm Ser B 8:76–137
Barattolo F (2002) Late Cretaceous-Paleogene Dasycladaleans and the K/T boundary problem. In: Bucur II, Filipescu S (eds), Research advances in calcareous algae and microbial carbonates, Proc of the 4th IFAA Reg Meeting, Cluj Napoca, Romania, 29 August–5 September 2001, pp 17–40
Berberian M, King GCP (1981) Towards a paleogeography and tectonic evolution of Iran. Can J Earth Sci 18:210–265
Blau J, Lukas V, Stein R (1987) Octotriangulella bonenburgensis n. gen., n. sp., ein Anomuren-Koprolith aus dem unteren Lias (Sinemur). N Jb Geol Paläont Mh 11(1987):641–646
Blau J, Grün B, Senf M (1993) Crustaceen-Koprolithen aus der Trias der westlichen Tethys (Lienzer Dolomiten, Österreich; Pragser Dolomiten, Italien) und Gondwana-Westrand (oberes Magdalenatal, Kolumbien, Südamerika). Paläont Z 67:193–214
Brönnimann P (1977) A new favreine coprolite (Crustacea, Decapoda) from the Jurassic of Iran and the Cretaceous of the Dinarids, Jugoslavia. Paläont Z 51:94–101
Brönnimann P, Norton P (1960) On the classification of fossil faecal pellets and description of new forms from Cuba, Guatemala and Libya. Mitt Ges Bergbaustud Österr 21:929–940
Brönnimann P, Caron JP, Zaninetti L (1972) Parafavreina, n. gen., a new thalassinid anomuran (Crustacea, Decapoda) coprolite form-genus from the Triassic and Liassic of Europa and North Africa. Mitt Gesell Geol Bergbaustud Österr 21:941–956
Buchs DM, Guex J, Stucki J, Baumgartner PO (2009) Paleocene Thalassinidea colonization in deep-sea environment and the coprolite Palaxius osaensis n. ichnosp, Southern Costa Rica. Rev Micropaléont 52:123–129
Bujtor L (2011) The early Valanginian ammonite, brachiopod and crustacean fauna of the Mecsek Mts. and its relationships with the embryonic shallow water hydrothermal vent at Zengővárkony (Mecsek Mts., South Hungary). Cretac Res 32:565–574
Bujtor L (2012) A Valanginian crustacean microcoprolite ichnofauna from the shallow-marine hydrothermal vent site of Zengővárkony (Mecsek Mts., Hungary). Facies 58:249–260
Consorti L, Köroğlu F (2019) Maastrichtian-Paleocene larger foraminifera biostratigraphy and facies of the Şahinkaya Member (NE Sakarya Zone, Turkey): insights into the Eastern Pontides arc sedimentary cover. J Asian Earth Sci 183:103965
Dall W, Moriarty DJW (1983) Functional aspects of nutrition and digestion. In: Mantel LH (ed) The biology of Crustaceans, vol 5. Internal anatomy and physiological regulation. Academic Press, New York, pp 215–261
Dalvand M, Ashrafzadeh AR, Ahmadi Z (2015) Crustacen microcoprolites from Lower Cretaceous and Oligo-Miocene deposits, Persian Gulf. Iran. Journ Paleontol 34:211–216
Fenninger A, Hubmann B (1994) Helicerina kainachensis n. sp. – Crustaceen-Koprolithen aus Kalken der Kainacher Gosau (Obersanton bis Untercampan, Österreich, Steiermark). N Jb Geol Paläont Mh 1:23–33
Gavrilov YO, Shchepetova EV (2000) Diagenetic mineral formation in biogenic structures (Paleogene, Northeastern Caucasus). Lithol Min Res 35(6):546–555
Gischler E, Blau J, Peckman J (2017) Palaxius floridanus n. isp., a new structured callianassid crustacean microcoprolite from the Pleistocene of south Florida. J Paleont 91(5):981–986
Hottinger L (2009) The Paleocene and earliest Eocene foraminiferal Family Miscellaneidae: neither nummulitids nor rotaliids. Notebooks on Geology Article 2009/06
Hottinger L (2014) Paleogene larger rotaliid foraminifera from the Western and Central Neotethys. Springer, Heidelberg, p 196
Kietzmann DA, Palma RM (2014) Early Cretaceous crustacean microcoprolites from Sierra de la Cara Cura, Neuquén Basin, Argentina: taphonomy, environmental distribution, and stratigraphic correlation. Cretac Res 49:214–228
Moon YW, Kim HH (1999) Morphological study of the digestive tract of the mud crab (Hemigrapsus penicillatus De Haan) and the symbiotic crab (Pinnotheres cyclinus Shen). Korean J Biol Sci 3(4):407–412
Motaharian A, Aghanabati A, Ahmadi V, Meisami A (2014) Crustacean microcoprolites of Middle and Upper Jurassic in Zagros (Southwest Iran). MAGTN Res Rep 2:295–304
Powell RR (1974) The functional morphology of the fore-gut of the thalassinid crustaceans, Callianassa californiensis and Upogebia pugettensis. Univ California Publ Zool 102:1–3
Rindsberg AK, Kopaska-Merkel DC (2013) Paleoecology and diagenesis of Parafavreina ziczac isp nov, a crustacean microcoprolite from the Upper Jurassic (Oxfordian) Smackover Formation of Alabama. Bull Alabama Mus Nat Hist 31(2):74–93
Rostami F, Ahmadi V (2013) Crustacean Microcoprolites from the Upper Jurassic - Lower Cretaceous of the Northern East and East Shiraz (Fars Province). Geosci Journ 22(86):131–134
Schlagintweit F, Rashidi K (2019) Serrakielina chatorshiana gen. et sp. nov., and other (larger) benthic Foraminifera from Danian-Selandian carbonates of Mount Chah Torsh (Yazd Block, Central Iran). Micropaleontol 65(4):305–338
Schlagintweit F, Muñoz-Moreno A, Martín-Chivelet J, Rosales I (2016) Sulcusina iberica n. ichnogen., n. ichnosp., a new crustacean microcoprolite from the Upper Cretaceous of Spain. Cretac Res 72:26–31
Schweigert G, Seegis DB, Fels A, Leinfelder RR (1997) New internally structured decapod microcoprolites from Germany (Late Triassic/Early Miocene), Southern Spain (Early/Middle Jurassic) and Portugal (Late Jurassic): taxonomy, palaeoecology and evolutionary implications. Paläont Z 71(1–2):51–69
Senowbari-Daryan B (1979) Anomuren-Koprolithen aus der Obertrias der Osterhorngruppe (Hintersee/Salzburg, Österreich). Ann Naturhist Mus Wien 82:99–107
Senowbari-Daryan B (1988) Lercarina tintinnicanalis n. gen., n. sp. (Crustaceen-Koprolith) aus dem unteren Perm von Sizilien. Mitt Gesell Geol Bergbaustud Österr 34/35:301–309
Senowbari-Daryan B, Grötsch J (1992) Palaxius salataensis: a Cretaceous anomuran microcoprolite from the MIT Guyot (northwest Pacific Ocean). Ichnos 2:85–88
Senowbari-Daryan B, Kube B (2003) The ichnogenus Palaxius (crustacean coprolite) and description of P. hydranensis n. sp. from the Upper Triassic (Norian part of “Pantokrator” limestone) of Hydra (Greece). Paläont Z 77(1):115–122
Senowbari-Daryan B, Schäfer P, Catalano R (1979) Helicerina siciliana n. sp.,a new anomuran coprolite from Upper Triassic reef limestones near Palermo (Sicily). Boll Soc Paleontol Ital 18(2):3–8
Serra-Kiel J, Hottinger L, Caus E, Drobne K, Ferràndez C, Jauhri AK, Less G, Pavlovec R, Pignatti J, Samsó JM, Schaub H, Sirel E, Strougo A, Tambareau Y, Tosquella J, Zakrevskaya E (1998) Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bull Soc Geol France 169:281–299
Sirel E (1998) Foraminiferal description and biostratigraphy of the Paleocene - lower Eocene shallow-water limestones and discussion of the Cretaceous - Tertiary boundary in Turkey. General directorate of the Mineral Research and Exploration (Turkey), Monography Series:1–117
Sirel E (1999) Four new genera (Haymanella, Kayseriella, Elazigella, and Orduella) and one new species of Hottingerina from the Paleocene of Turkey. Micropaleont 45:113–137
Zanchi A, Zanchetta S, Garzanti E, Balini M, Berra F, Mattei M, Muttoni G (2009) The Cimmerian evolution of the Nakhlak-Anarak area, Central Iran, and its bearing on the reconstruction of the history of the Eurasian margin. In: Brunet MF, Wilmsen M, Granath JW (eds) South Caspian to Central Iraan Basin. Geol Soc London Spec Publ vol.312, pp 261–286
Thanks to Lorenzo Consorti (Trieste) for commenting on the Rotaliidae. The two reviewers Diego Kietzmann (Buenos Aires) and Andrew A. Rindsberg (Livingston) provided helpful comments. Maurice Tucker (Bristol) is thanked for his careful editing.
Author information
Authors and Affiliations
Lerchenauerstr. 167, 80935, Munich, Germany
Felix Schlagintweit
Department of Geology, Yazd University, 89195-741, Yazd, Iran
Koorosh Rashidi
Department of Geology, Faculty of Science, Birjand University, Birjand, Iran
Schlagintweit, F., Rashidi, K. & Raisossadat, S.N. Broennimannia persica n. ichnogen., n. ichnosp., a new crustacean microcoprolite from the Paleocene of eastern Iran.
Facies66, 15 (2020). https://doi.org/10.1007/s10347-020-00599-5