
Abstract
A sclerochronological analysis was performed on Cladocora caespitosa corals from Late Pleistocene terraces near Taranto (Apulia, Italy) to reconstruct the main palaeoenvironmental conditions at the time of their growth. The fossil corallites were sampled in the Santa Teresiola uplifted bank or ‘open frame reef’ attributed to the Last Interglacial Period. The typical, annual growth pattern of the temperate coral with two alternate high- and low-density bands allowed the reconstruction of two multidecadal growth curves of 61 and 95 years. Trend analysis showed oscillations in annual growth rates similar to those observed in recent, living colonies sampled along a north–south latitudinal transect around the Italian and Croatian coasts as far as Tunisia. The mean growth rate of the fossil reef (4.2 ± 2 mm year−1) is comparable to those measured on colonies living in the coldest part of the Mediterranean Sea. The comparison with data from living Croatian banks shows how fossil C. caespitosa lived in a semi-enclosed environment characterized by seasonal inputs of fresh, cold water. The greatest variations in decadal growth rates of the fossil colonies support the hypothesis of larger amplitude of the seasonal cycles in the past. The death of the fossil bank was probably due to a sudden alluvial input that suffocated the reef with a great amount of mud. Another possible cause of the death of the bank was a prolonged increase in summer temperatures that caused colony mortality and enhanced algal colonization.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
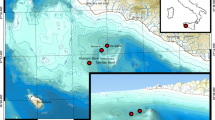
Cladocora caespitosa (L.) is a Mediterranean, zooxanthellate scleractinian coral belonging to the family Faviidae (Zibrowius 1980) that colonized the Mediterranean Sea since at least the Late Pliocene (Aguirre and Jiménez 1998; Dornbos and Wilson 1999). At present, the living C. caespitosa is one of the more important benthic carbonate producers in the Mediterranean Sea (Peirano et al. 2001), which is found from the sea surface down to 40 m in depth. Where present, it can form build-ups (banks) rising up to 1 m above the surrounding seafloor and covering several square meters. Today, living banks of coral comparable to tropical reefs were discovered in three localities along the coast of Croatia (Fig. 1) characterized by low mean annual temperatures (Kružić and Benković 2008). The greatest bank is located in one ‘sea–lake’ of the island of Mljet. This bank consists of some hundreds of C. caespitosa colonies, which are fused together and cover 650 m2 on the seabed in 4–18 m water depth (Kružić and Požar-Domac 2003).

The colonies of C. caespitosa are phaceloid, i.e., the tubular corallites develop vertically with sub-parallel growth axes of the main branches. The rectilinear continuous growth of corallites facilitates the measurement of the annual growth rate through X-ray methodology (sclerochronology) avoiding problems related to three-dimensional development of corallites of tropical, massive coral species (Peirano et al. 1999). X-ray imaging has shown that C. caespitosa deposits two density bands per year; a high-density band (HD), deposited during the rainy, cold autumn, and winter seasons and a lower-density band deposited during the hot, dry summer (Peirano et al. 1999, 2005). The analysis of the decadal growth of a great number of colonies sampled at depths between 4 and 30 m showed a sinusoidal pattern in accordance with the air temperature trend (Peirano et al. 2004). Such a temperature–growth relationship in C. caespitosa was outlined from Silenzi et al. (2005). These authors found a strict correlation between Sr/Ca-derived sea-surface temperatures (SST) of the coral skeleton and recorded SST of the ambient water mass. These studies also proved that the deepest colonies have yearly growth rates related to SST despite the fact that the living depth is below the seasonal thermocline depth.
Large fossil banks of C. caespitosa are known in the Mediterranean from the Late Pliocene (Aguirre and Jiménez 1998; Dornbos and Wilson 1999), from the Early Pleistocene (Bernasconi et al. 1997), from the Middle and Late Pleistocene and from the Holocene (for a review see Peirano et al. 1998, 2004) (Fig. 1).
In the Pleistocene, shorelines represented by both abrasion platforms and marine calcarenite deposits, frequently included C. caespitosa (Peirano et al. 1998, 2004). During warm climatic phases, huge banks or ‘open-frame reefs’ were abundant. In some cases, Senegalese fauna (Strombus bubonius, Cardita calyculata senegalensis, and Hyotissa hyotis) was found associated with the coral. Senegalese species are indicators of the warmest climate phase that characterized the Last Interglacial Period corresponding to the marine isotope substage (MISs) 5.5 (ca. 132–116 ka = Late Pleistocene) (Issel 1914; Lambeck and Chappell 2001; Shackleton et al. 2003).
This paper focuses on the growth of one huge, fossil uplifted bank of C. caespitosa near Taranto (Apulia, Italy). Growth time series are calculated through sclerochronology and compared with those of recent, living colonies and banks around the Italian, Croatian, and Tunisian coasts. The comparison allows inferring the palaeoclimatic conditions and the environment that favored the formation of the C. caespitosa bank.
Materials and methods
Fossil corallites of Cladocora caespitosa were sampled from the deposits of the Mar Piccolo of Taranto (Apulia, South Italy) (Fig. 2). The sample site, ‘Santa Teresiola’ (ST), is a fossil bank located on a raised marine terrace 12 m above mean sea-level (Dai Pra and Stearns 1977; Hearty and Dai Pra 1992; Belluomini et al. 2002). The bank is formed by numerous, contiguous colonies of coral embedded in a sandy matrix up to 1 m high covering a surface of nearly 0.6 km2. Different U/Th datings and correlation with nearest deposits suggest that the reef belongs to the MISs 5.5 (for a review see Mastronuzzi et al. 2003).

Fossil Cladocora caespitosa reef: a distribution of C. caespitosa (asterisks) and Senegalese fauna associations (plus symbol) in Late Pleistocene deposits of Mar Piccolo di Taranto with indication of the sampling site (ST Santa Teresiola). b Aspect of the reef of Santa Teresiola. c Detail of the height of the reef (the distance between the red points on the yellow bar is 20 cm). a and b from Mastronuzzi et al. (2003)
Two samples (ST1, ST2), 36 and 46 cm long, respectively, were carefully detached from the reef following the corallites growth from the top to the bottom of two different colonies that lived some meters apart.
To compare past fossil growth rates with those of recent colonies, 11 samples of C. caespitosa were collected from a depth of 7–22 m along a north–south transect along the Italian and the Croatian coast as far south as Tunisia to cover a latitudinal gradient in terms of temperature range and seasonal variability (Fig. 1). Corallites were cleaned of epibionts and organic matter and the polyps were removed by immersion in hydroperoxide (30%). To calculate yearly growth rates, X-rayed positive corallite images were analyzed through Coral XDS image-analyzing software version 3.0, available on the Web site of the National Coral Reef Institute of Florida, USA (http://www.nova.edu/ocean/coralxds/index.html).
For climate comparison, annual SST from 1985 to 2004 were calculated from monthly data downloaded from NASA Physical Oceanography Distributed Active Archive Centre (PODAAC) through the Ocean ESIP Tool (POET) (http://poet.jpl.nasa.gov//). Data on monthly air temperatures (AirT) were downloaded from the site of the Italian Agency of the Ministry of Environment (APAT) that collects and distributes the environmental data through the SINAnet–SCIA (http://www.scia.sinanet.apat.it/).
Results
Fossil samples
The two ST fossil samples showed clear, alternating banding patterns as seen in recent corallites in agreement with the findings of Peirano et al. (2004). The distance between high-density peaks allowed the estimation of the mean annual growth rate of 4.1 ± 2 mm year−1.
The Santa Teresiola X-rayed C. caespitosa allowed the calculation of two individual lifespans with 67 (ST1) and 95 (ST2) years, respectively (Fig. 3). The analysis of 3-year smoothed average trend curves showed a great similarity of the two growth curves and a good superimposition of peaks and valleys was found with a negative shift of the ST1 curve of nearly 10 years (R = 0.44; n = 65; P < 0.001). Trend analysis evidenced also oscillations in growth rates of Santa Teresiola samples over 10 years (Fig. 3).

Growth rates of Cladocora caespitosa from Santa Teresiola Late Pleistocene reef samples ST1 and ST2. ST1 curve is shifted of 10 years to evidence the similarity of the two growth curves. Years are counted from the top of the colonies (from right to left). In bold the 3-year smoothed average trends. The dashed lines are the sinusoidal multiannual trends generated by a sixth-order exponential smoothing average
A general positive trend in ST2 growth curve is determined from the sudden positive increase of the colony growth in the last 7–10 years.
Recent samples
The oldest living colony from Fiascherino was the only one that allowed the long-term comparison between the coral growth rates and the air temperatures (Table 1, Fig. 4). The comparison showed decadal oscillations similar to those observed in fossil samples but with a lesser amplitude (mean growth rates: 3.3 ± 0.4 mm year−1). The 3-year smoothed averages showed growth rates correlated to air temperature (R = 0.32; n = 43; P < 0.05) that, in turn, was highly correlated to SST (R = 0.88; n = 12; P < 0.001).

Cladocora caespitosa: annual trends and 3-year smoothed average trends (in bold) of air temperature, SST and mean growth rates of the colony of Fiascherino
Taking into account the temperature gradients, recent colonies living in coastal waters showed annual growth rates (Table 1) negatively correlated (R = −0.60; n = 11; P < 0.05) with mean annual SST (Table 2, Fig. 5a). The maximum growth rates of Cladocora caespitosa is attained with mean annual SST of 17–18.5°C recorded in the northern part of the Mediterranean (Table 1, Fig. 5a).

Mean annual growth rates of Cladocora caespitosa versus mean annual SSTs: a Black quadrats represent the living colonies analyzed in the present work and the pointed line is the calculated growth relationship; black triangles represent the fossil colonies of Santa Teresiola. Hatched lines are standard deviations of back-calculated SSTs of fossil samples. b Growth curves of coral banks: white triangles are the living banks of Croatia (data from Kružić and Benković 2008) and black triangles are from Santa Teresiola
Discussion
The analysis of multidecadal time series of living colonies of Cladocora caespitosa showed that greater growth rates were associated with coldest mean annual SST. This growth pattern, found also in the scleractinian Balanophyllia europaea by Goffredo et al. (2008), seems to be characteristic of the Mediterranean species of zooxanthellate corals. However, to form a mature coral bank special environmental conditions are necessary, as outlined by Kružić and Benković (2008).
To infer past climate conditions, the linear relationship that correlates the SST with annual growth rates of C. caespitosa was used to back calculate the mean SST temperatures of fossil colonies. The results showed that the fossil bank grew in one environment characterized by mean annual SST of 17.6 ± 2.2°C, similar to those recorded in the Northern Adriatic (Table 3, Fig. 5a). Although data on mean annual SST for the C. caespitosa living banks (Kružić and Benković 2008, unpublished data) are limited to the year 2004 (Table 3), the comparison between growth data of single colonies and those of the banks leads to the assumption that the linear relationship between SST and growth rates may change into a curve described by a significant quadratic relationship (R = 0.92; n = 5; P < 0.05) (Fig. 5b). When mean annual SST is below 15–16°C and monthly temperatures at depth last for months below 14°C, as in the case of Pag and Prvic coral banks at the Croatian coast (Kružić and Benković 2008), C. caespitosa shows lower mean growth rates due to a reduction in growth or no growth (see also Montagna et al. 2007).
The fossil Santa Teresiola coral bank probably developed under similar SST regimes which are comparable to those recorded today in the coldest regions of Eastern Adriatic Sea, where cold winds or inputs of freshwater may lower the SST temperature on the coral banks (Benović et al. 2000; Novosel et al. 2004; Montagna et al. 2007; Kružić and Benković 2008).
Mastronuzzi et al. (2003) and Mastronuzzi (2006) concluded that the fossil C. caespitosa bank of Santa Teresiola existed in a lagoon affected by inputs of freshwater. Such a palaeoenvironmental setting matches the situation of the living coral bank of Mljet, Croatia. The bank grows in a semi-enclosed coastal lagoon that maintains water exchange with the open Adriatic Sea through a 4.5 m deep channel. In this lagoon, SST vary between 9 and 29°C and salinity shifts from 36.3 to 39 PSU (Vaniček et al. 2000; Kružić and Požar-Domac 2003). Although the Santa Teresiola reef refers to a warm climatic phase of the Pleistocene, the differences between the temperature of the lagoon and the warmer open-sea temperatures of MISs 5.5 could be explained by freshwater coming from underwater, karstic springs that are common today both in Croatia and in the Taranto area (Pagliarulo and Bruno 1990; Kružić and Benković 2008). These assumptions also agree with palaeontological records of other fossil reefs found in alluvial deposits dating back as far as to the Late Pliocene (Aguirre and Jiménez 1998; Fornos et al. 1996) where ‘beds’ of the bivalve Ostrea spp., a suspension feeder characteristic of environments with cold seawater, lower salinity and high turbidity, forms the basement or is intercalated in coral banks (Bernasconi et al. 1997; Aguirre and Jiménez 1998; Dornbos and Wilson 1999).
The comparison of the growth characteristics of fossil Santa Teresiola samples and long-living colonies evidenced larger oscillations in growth rates that are in agreement with Mediterranean climate reconstruction of Felis et al. (2004) and with palaeoclimate reconstruction of Martrat et al. (2007) from the Iberian margin. The alternation of dry and wet phases were probably more accentuated than today (Felis et al. 2004) and could have influenced karstic spring inputs, thus causing the growth rate oscillations observed in the Santa Teresiola bank.
The Santa Teresiola coral bank sedimentary sequence is covered with a discontinuous layer of reddish silty sand rich in Arca sp., Ostrea sp., Glycymeris sp., Cerastoderma sp. and riverine mollusks (Caldara and Laviano 1980). The general characters of the Santa Teresiola section and the palaeoecological studies performed on the reddish level suggest that the C. caespitosa bank lived in correspondence of maximum transgression or sea-level highstand. A massive upload of sediment due to alluvial inputs is the most probable cause of the decline of the Santa Teresiola bank, as indicated by the presence of the reefs as a unique entity in ‘vital’ position. Maximum transgression sea-level scenarios are also assumed for Pleistocene C. caespitosa banks on the Tyrrhenian side of Calabria, Italy (Bernasconi et al. 1997), whilst C. caespitosa colonies described from Late Pleistocene (MISs 5e-d) coastal facies on Rhodes, Greece, developed at the beginning of a regressive sequence (see Titschack et al. 2008).
Another possible cause of death of the fossil coral bank could be a prolonged increase in temperature. In recent colonies, the rise in summer temperature recorded throughout the Mediterranean enhanced corallite growth (Peirano et al. 2005) and a positive trend in coral growth rates was evidenced in the Mljet bank (Peirano and Kružic 2004). However, if the temperature level climbs up to 26–28°C, and this lasts for more than one month, then the death of coral tissue is likely to happen (Rodolfo-Metalpa et al. 2006), as was observed both in Fiascherino (Rodolfo-Metalpa et al. 2000, 2005) and in the Mljet reef (Kružić and Požar-Domac 2002). The rise in temperature also favors the spreading of the invasive tropical green alga Caulerpa racemosa var. cylindracea in the Mediterranean, that presently is overgrowing the bank of Mljet (Kružić et al. 2008).
Conclusions
The analysis on fossil samples from Santa Teresiola near Taranto confirmed the great importance of the coral Cladocora caespitosa as a proxy for the study of past climate. The Taranto bank gives the opportunity to reconstruct growth rates of a coastal, long-living species at a yearly resolution, a rare, if not unique, opportunity for the Mediterranean. Other fossil/subfossil reefs of C. caespitosa are known in the Mediterranean (Peirano et al. 1998, 2004), however, these remain poorly studied due to the fact that many of them are underwater and therefore difficult to sample, or the aragonite corallites suffer from intense diagenetic alteration.
Sclerochronological analysis of the Santa Teresiola reef allows studying a particular coastal environment elucidating some details on the littoral ecosystems of the Quaternary. This approach may help palaeontologists to differentiate fossil deposits that are otherwise difficult to date (Goy et al. 2003), or could be useful in geochemical analysis to improve the reconstruction of past temperature and carbonate deposition of C. caespitosa (Silenzi et al. 2005; Montagna et al. 2007) at seasonal scale.
References
Aguirre J, Jiménez AP (1998) Fossil analogues of the present-day ahermatypic Cladocora caespitosa coral banks: sedimentary setting, dwelling community and taphonomy (Late Pliocene, W Mediterranean). Coral Reefs 17:203–213. doi:10.1007/s003380050119
Belluomini G, Caldara M, Casini C, Cerasoli M, Manfra L, Mastronuzzi G, Palmentola G, Sanso P, Tuccimei P, Vesica PL (2002) Age of Late Pleistocene shorelines, morphological evolution and tectonic history of Taranto area, southern Italy. Quat Sci Rev 21:427–454. doi:10.1016/S0277-3791(01)00097-X
Benović A, Lučić D, Onofri V, Peharda M, Caric M, Jasprica N, Sobanovic S, Olic C (2000) Ecological characteristics of the Mljet Island seawater lakes (South Adriatic Sea) with special reference to their resident population of Medusae. Sci Mar 64:197–206. doi:10.3989/scimar.2000.64s1197
Bernasconi MP, Corselli C, Carobene L (1997) A bank of the scleractinian coral Cladocora caespitosa in the Pleistocene of the Crati valley (Calabria, southern Italy): growth versus environmental conditions. Boll Soc Paleontol Ital 36:53–61
Caldara M, Laviano A (1980) Osservazioni paleontologiche e paleoecologiche su un deposito quaternario affiorante a Punta delle Penne (Mar Piccolo—Taranto). Annali Università di Ferrara. Sez IX(VI):57–71
Dai Pra G, Stearns CE (1977) Sul Tirreniano di Taranto. Datazioni su coralli con il metodo del 230Th/234U. Geol Rom 16:231–242
Dornbos SQ, Wilson MA (1999) Paleoecology of a Pliocene coral reef in Cyprus: recovery of a marine community from the Messinian salinity crisis. N Jahrb Geol Palaeontol Abh 21:103–118
Felis T, Lohmann G, Kuhnert H, Lorenz SJ, Scholz D, Pätzold J, Al-Rousan SA, Al-Moghrabi SM (2004) Increased seasonality in Middle East temperatures during the Last Interglacial Period. Nature 429:164–168. doi:10.1038/nature02546
Fornos JJ, Baron A, Pons GX (1996) Arrecifes de corales hermatípicos (Cladocora caespitosa) en el relleno holoceno de la zona de Es Grau (Menorca, Mediterraneo Occidental). Geogaceta 20:303–306
Goffredo S, Caroselli E, Mattioli G, Pignotti E, Zaccanti F (2008) Relationships between growth, population structure and sea-surface temperature in the temperate solitary coral Balanophyllia europaea (Scleractinia, Dendrophylliidae). Coral Reefs 27:623–632. doi:10.1007/s00338-008-0362-y
Goy JL, Hillaire-Marcel C, Zazo C, Ghaleb B, Dabrio CJ, González A, Bardají T, Civis J (2003) U-series ages of coral-bearing littoral deposits with Strombus bubonius of OIS 7 from La Marina (Alicante, SE Spain). A reappraisal of the Thyrrenian chronostratigraphy in the Mediterranean Sea. In: Mastronuzzi G, Sansò P (eds) Project IGCP 437—Coastal environmental change during sea-level highstands: a global synthesis with implication for management of future coastal change. Puglia 2003 final conference, Quaternary coastal morphology and sea-level changes GI2S coast, Research publication 4, pp 109–111
Hearty PJ, Dai Pra G (1992) The age and stratigraphy of Quaternary coastal deposits along the Gulf of Taranto (south Italy). J Coast Res 8:882–905
Issel A (1914) Lembi fossiliferi quaternari e recenti nella Sardegna meridionale. Accad Nazionale Lincei Ser 5:759–770
Kružić P, Benković L (2008) Bioconstructional features of the coral Cladocora caespitosa (Anthozoa, Scleractinia) in the Adriatic Sea (Croatia). Mar Ecol (Berl) 29:125–139. doi:10.1111/j.1439-0485.2008.00220.x
Kružić P, Požar-Domac A (2002) Skeleton growth rates of coral bank of Cladocora caespitosa (Anthozoa, Scleractinia) in Veliko Jezero (Mljet National Park). Period Biol 104:123–129
Kružić P, Požar-Domac A (2003) Banks of the coral Cladocora caespitosa (Anthozoa, Scleractinia) in the Adriatic Sea. Coral Reefs 22:536. doi:10.1007/s00338-003-0345-y
Kružić P, Zuljević A, Nikolić V (2008) The highly invasive alga Caulerpa racemosa var. cylindracea poses a new threat to the banks of the coral Cladocora caespitosa in the Adriatic Sea. Coral Reefs 27:441. doi:10.1007/s00338-008-0358-7
Lambeck K, Chappell J (2001) Sea-level change during the last glacial cycle. Science 292:679–686. doi:10.1126/science.1059549
Martrat B, Grimalt JO, Shackleton NJ, de Abreu L, Hutterli MA, Stocker TF (2007) Four climate cycles of recurring deep and surface water destabilizations on the Iberian margin. Science 517:502–507. doi:10.1126/science.1139994
Mastronuzzi G, Sansò P, Tuccimei P, Vesica P (2003) Stop 3.3.1 - The Cladocora caespitosa bank of Santa Teresiola locality. In: Mastronuzzi G, Sansò P (eds) Project IGCP 437—Coastal Environmental change during sea-level highstands: a global synthesis with implication for management of future coastal change. Puglia 2003 Final conference, Quaternary coastal morphology and sea-level changes—Field guide GI2S Coast, Research Publication 5, pp 85–88
Mastronuzzi G (2006) Evoluzione dell’orografia antica della città di Taranto. Atti del Seminario “Dal Kastron bizantino al Castello aragonese”, 17 novembre 2004, Taranto, Castello Aragonese, Scorpione Editrice, Taranto, pp 123–140
Montagna P, McCulloch M, Mazzoli C, Silenzi S, Odorico R (2007) The non-tropical coral Cladocora caespitosa as the new climate archive for the Mediterranean: high-resolution (weekly) trace element systematics. Quat Sci Rev 26:441–462. doi:10.1016/j.quascirev.2006.09.008
Novosel M, Požar-Domac A, Pasarić M (2004) Diversity and distribution of the bryozoa along underwater cliffs in the Adriatic Sea with special reference to thermal regime. PSZNI Mar Ecol 25:155–170. doi:10.1111/j.1439-0485.2004.00022.x
Pagliarulo R, Bruno G (1990) Implicazioni tettonico—strutturali nella circolazione idrica profonda nell’area del Mar Piccolo di Taranto (Puglia). Boll Soc Geol It 109:307–312
Peirano A, Morri C, Mastronuzzi G, Bianchi CN (1998) The coral Cladocora caespitosa (Anthozoa, Scleractinia) as a bioherm builder in the Mediterranean Sea. Mem Descr Carta Geol It 52:59–74
Peirano A, Morri C, Bianchi CN (1999) Skeleton growth and density pattern of the zooxanthellate scleractinian Cladocora caespitosa (L.) from the Ligurian Sea (NW Mediterranean). Mar Ecol Prog Ser 185:195–201. doi:10.3354/meps185195
Peirano A, Morri C, Bianchi CN, Rodolfo-Metalpa R (2001) Biomass, carbonate standing stock and production of the Mediterranean coral Cladocora caespitosa (L.). Facies 44:75–80. doi:10.1007/BF02668168
Peirano A, Kružic P (2004) Growth comparison between Ligurian and Adriatic samples of the coral Cladocora caespitosa: first results. Biol Mar Medit 11:166–168
Peirano A, Morri C, Bianchi CN, Aguirre J, Antonioli F, Calzetta G, Carobene L, mastronuzzi G, Orrù P (2004) The Mediterranean coral Cladocora caespitosa: a proxy for past climate fluctuations? Glob Planet Change 40:195–200. doi:10.1016/S0921-8181(03)00110-3
Peirano A, Abbate M, Cerrati G, Difesca V, Peroni C, Rodolfo-Metalpa R (2005) Monthly variations in calix growth, polyp tissue and density banding of the Mediterranean scleractinian Cladocora caespitosa (L.). Coral Reefs 24:404–409. doi:10.1007/s00338-005-0020-6
Rodolfo-Metalpa R, Bianchi CN, Peirano A, Morri C (2000) Coral mortality in NW Mediterranean. Coral Reefs 19:24. doi:10.1007/s003380050221
Rodolfo-Metalpa R, Bianchi CN, Peirano A, Morri C (2005) Tissue necrosis and mortality of the temperate coral Cladocora caespitosa. Ital J Zool (Modena) 72:271–276
Rodolfo-Metalpa R, Richard C, Allemand D, Ferrier-Pages C (2006) Growth and photosynthesis of two Mediterranean corals, Cladocora caespitosa and Oculina patagonica, under normal and elevated temperatures. J Exp Biol 209:4546–4556. doi:10.1242/jeb.02550
Shackleton NJ, Sanchez-Goni MF, Pailler D, Lancelot Y (2003) Marine isotope substage 5e and the Eemian Interglacial. Glob Planet Change 36:151–155. doi:10.1016/S0921-8181(02)00181-9
Silenzi S, Bard E, Montagna P, Antonioli F (2005) Isotopic and elemental records in a non-tropical coral (Cladocora caespitosa): discovery of a new high-resolution climate archive for the Mediterranean Sea. Glob Planet Change 49:94–120. doi:10.1016/j.gloplacha.2005.05.005
Titschack J, Nelson NC, Beck T, Freiwald A, Radtke U (2008) Sedimentary evolution of a Late Pleistocene temperate red algal reef (Coralligène) on Rhodes, Greece: correlation with global sea-level fluctuations. Sedimentology 55:1747–1776. doi:10.1111/j.1365-3091.2008.00966.x
Vaniček V, Juračić M, Bajraktarević Z, Cosović V (2000) Benthic foraminiferal assemblages in a restricted environment—an example from the Mljet lakes (Adriatic Sea, Croatia). Geol Croat 53:269–279
Zibrowius H (1980) Les Scléractiniaires de la Méditerranée et de l’Atlantique nord-oriental. Mém Inst océanogr 11:1–284
Acknowledgments
We thank A. Freiwald who encouraged the submission of the manuscript, J. Titschack and two anonymous reviewers who greatly improved our work. We are grateful to S. Cocito, C. Lombardi and S. Sgorbini for their assistance during diving and sampling operations. Special thanks also go to P. Ritrovato for his courtesy and support during the sampling operations at Scoglitti (Sicily) and to the directors and staffs of the Marine Protected Areas of Capo Rizzuto (Italy—Calabria) and of Mljet (Croatia). This research was supported by the Italian project VECTOR and by a contribution to the IGCP project no. 495 “Quaternary Land–Ocean Interactions: Driving Mechanisms and Coastal Responses” dell’UNESCO–IUGS (Project leaders: Dr. A. Long, University of Durham, UK; Dr. S. Islam, University of Chittangong, Bangladesh).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Peirano, A., Kružić, P. & Mastronuzzi, G. Growth of Mediterranean reef of Cladocora caespitosa (L.) in the Late Quaternary and climate inferences. Facies 55, 325–333 (2009). https://doi.org/10.1007/s10347-008-0177-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10347-008-0177-x