
Abstract
The snow leopard Panthera uncia coexists with the wolf Canis lupus throughout most of its distribution range. We analysed the food habits of snow leopards and wolves in their sympatric range in the Karakoram mountains of Pakistan. A total of 131 genotyped scats (N = 74, snow leopard; N = 57, Tibetan wolf) were collected during the cold periods (i.e. winter and spring) of 2011 and 2012 in the Hushey valley. Large mammals, i.e. livestock and ibex, accounted for 84.8 and 83.1% of the diet (relative frequency) of the snow leopard and the wolf, respectively. Domestic prey was the staple of the diet of both snow leopards (66.6%) and wolves (75.1%). Ibex Capra ibex, the only wild ungulate in our study area, contributed 18.2 and 16.9% of relative frequencies in the diets of the snow leopard and the wolf, respectively. In winter, the snow leopard heavily relied on domestic sheep (43.3%) for food, whereas the wolf preyed mainly on domestic goats (43.4%). Differently from other study areas, both snow leopards and wolves showed no apparent prey preference (Jacobs index: snow leopard min. − 0.098, max. 0.102; Tibetan wolf min. − 0.120, max. 0.03). In human depauperate areas, with livestock and only a few wild prey, should competitive interactions arise, two main scenarios could be expected, with either predator as a winner. In both cases, the best solution could primarily impinge on habitat restoration, so that a balance could be found between these predators, who have already coexisted for thousands of years.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Large carnivore species can occur in simpatry throughout the world because of prey partitioning, body size differences, different activity rhythms or habitat use (Palomares et al. 1996; Lovari et al. 2015; Sugimoto et al. 2016), which limit interspecific competition (Palomares and Caro 1999; Donadio and Buskirk 2006). When they compete, it is apparently through interference, where usually the smaller species experiences negative effects on its population dynamics (for reviews on competition among carnivores: Palomares and Caro 1999; Donadio and Buskirk 2006). Although there is a wealth of studies showing interference competition among carnivores, exploitative competition has not been documented so far in this order (but see Watts and Holekamp 2008). Information is limited on the role of the snow leopard Panthera uncia within its ecosystem, as well as its ecological relationships with the carnivore guild of wolf Canis lupus, brown bear Ursus arctos lynx Lynx lynx and common leopard Panthera pardus (Schaller 2016). These large carnivores may use the same prey species, and competition for resource exploitation could arise between them, although relevant evidence of effects at the population level has been lacking so far. The potential for competition is greater where the local resource spectrum is poor (Stephens and Krebs 1986; Peiman and Robinson 2010, for a review); thus, studies in such areas should be especially important for conservation purposes.
The snow leopard and the Tibetan grey wolf Canis lupus filchneri are key predators of the Himalayan rangelands (Fox et al. 1991; Mishra 1997). The former is globally listed as ‘vulnerable’ in the IUCN Red List of Threatened Species (IUCN Red List: http://www.iucnredlist.org/details/22732/0 - Accessed on 26th October 2017), whereas the latter is common throughout Central Asia (IUCN Red List: “least concern”: http://www.iucnredlist.org/details/3746/0 - Accessed on 10th April 2017), even locally considered as a pest (Lescureux 2006). As to Pakistan, the Tibetan grey wolf has been listed as ‘endangered’ in the red list of Pakistan Mammals (Sheikh and Molur 2004). Information on food habits is a crucial first step to understand ecological features and to predict predator influence on prey communities (Shrestha 2008; Anwar et al. 2011).
Wild ungulates are the main prey of the snow leopard (Lovari et al. 2013a; Lyngdoh et al. 2014), e.g. Siberian ibex Capra ibex sibirica (Anwar et al. 2011), bharal Pseudois nayaur and Himalayan tahr Hemitragus jemlahicus (Oli et al. 1993; Lovari et al. 2013b). Similarly, wild ungulates build up also the largest proportion of the wolves’ diet in Central Asia (Schaller 1977; Blumestein 1993; Newsome et al. 2016). Where these two predators coexist, if the wild ungulate community is rich, their diet differentiates, with wolves preying on plain dwellers (e.g. gazelle Gazella spp., kiang Equus kiang and Marco Polo sheep Ovis ammon polii) and snow leopards selecting cliff-dwelling species (e.g. bharal: Chetri et al. 2017). By contrast, where species richness is low, diet overlap may increase (e.g. Kyrgyzstan: Jumabay-Uulu et al. 2013; NW China: Wang et al. 2014). In areas with high livestock densities, the percentage of domestic ungulates in the snow leopard diet may increase up to 70% (Bagchi and Mishra 2006; Anwar et al. 2011). Livestock is also frequently found in the wolf diet, ranging from 38 to 65% in northern Pakistan (Schaller 1977; Khan 2012). In one area of Nepal (Jackson 1996) and in one of northern Pakistan (Khan 2012; Khan et al. 2017), where both snow leopards and wolves live, most herders suspected snow leopards as responsible for livestock depredations, whereas in Ladakh, India, the wolf was considered responsible for most killings (Namgail et al. 2007). The high mountains of northern Pakistan host a declining population of snow leopards (Ud Din et al. 2016). In this region, wolf packs are also present, although the density of wild ungulates is very low (Schaller 1977; Blumestein 1993; Khan 2012). Thus, a high dietary overlap between these large carnivores would be expected. Should resource competition arise between these carnivores, the more generalist species (i.e. the wolf, Newsome et al. 2016) could outcompete the more specialised one (i.e. the snow leopard, Sharma et al. 2015; Lovari and Mishra 2016).
Our study objectives were to (i) investigate the food habits of the snow leopard and the Tibetan wolf in an area with a poor wild prey spectrum, in Northern Pakistan; (ii) estimate niche breadth, prey selection and diet overlap of these predators and (iii) assess the relative importance of wild and domestic prey in the diet of both predators.
Materials and methods
Study area
Our study was conducted in the largest protected area in Pakistan, the Central Karakoram National Park (hereafter, CKNP; about 10,557 km2, with the annexed buffer zone). This national park contains some of the world’s largest glaciers and the greatest concentration of high mountains on earth, ranging from 2000 to over 8000 m.a.s.l., including K2. As to its climate, the CKNP area can be considered a transition zone between the arid climates of western Asia and the semi-humid sub-tropics of southern Asia. The whole region is characterised by relatively dry summers, with an average annual precipitation of less than 300 mm in low-lying areas (Clemens and Nusser 1997), because of mountains limiting monsoon movements to the west and heavy winter snowfalls at the highest altitudes (WWF-Pakistan 2008). Five altitudinal ecological layers are recognised (Champion et al. 1965): Alpine dry steppe (2600–3000 m.a.s.l.), dominated by mugwort Artemisia spp. and juniper Juniperus spp.; sub-alpine scrub zone (3000–3500 m.a.s.l.), with birch forests Betula utilis, juniper Juniperus spp., wild rose Rosa canina and Hippophae rhamnoides; Alpine grasslands (3500–4200 m.a.s.l.); sub-nival (4200–5000 m.a.s.l.) and nival zones (over 5000 m a.s.l.), where only Salix denticulata and Mertensia tibetica may be found.
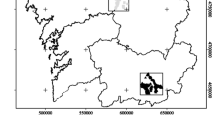
Our study area was located in the easternmost valley of the park, the Hushey valley (76 20′ E to 35° 27′ N), covering over 4000 km2 (map surface, actual size being over three times greater because of altitudinal development; Fig. 1), from 3000 to 5000 m.a.s.l., with a main N-S orientation. The valley is split into four main sub-valleys: Aling, Musherbrum, Ghandogoro and K6 (WWF-Pakistan 2008; Khan et al. 2014). The persistence of snow (December–May, i.e. winter and spring, hereafter ‘cold period’) and the frequent landslides on the only connecting road isolate the area for long periods. Livestock and subsistence agriculture are the main resources of the local population (c. 1200 humans, in 150 households). Livestock is composed of goats (500–600 individuals), sheep (c. 700), donkeys (c. 20), cattle (c. 200) and domestic yaks (c. 30). The Siberian ibex is the only wild ungulate in this area. Official counts of ibex contemporary to our study indicated about 370 individuals (Khan et al. 2014). Besides the snow leopard and the Tibetan wolf, the red fox Vulpes vulpes and the corsac fox Vulpes corsac are also present.

Location of the study area (WWF-Pakistan 2008)
The Himalayan snowcock Tetraogallus himalayensis, the chukar partridge Alectoris chukar and the Royle’s pika Ochotona roylei are also relatively abundant.
Data collection and diet analysis
Carnivore scats were collected monthly, along fixed trails evenly distributed through the four valleys of our study area (Fig. 1), for a total of nearly 35 km, during spring (March–May) and winter (December–February) 2011–2012. A thorough, conservative selection of scats was made in the field on the basis of different features (e.g. smell, position, size, contents and presence of tracks) to reduce the risk of collecting scats of other species (cf. Lovari et al. 2009). Furthermore, approximately 1 cm3 of each fresh scat was preserved in a vial with silica gel for later DNA analyses. Upon genetic validation (Jan E. Janečka, Duquesne University, USA, unpublished), we confirmed 90.5% of scats as belonging to the wolf (N = 63), while 93.7% were confirmed as belonging to the snow leopard (N = 79). We proceeded with diet analyses (131 scats: snow leopard, SL: N = 74; wolf, W: N = 57). Scats were sun-dried and then preserved in polyethylene bags, labelled with date, GPS coordinates and altitude. All the plastic bags were frozen (− 30 ° C) until the start of the diet analysis (Reynolds and Aebischer 1991).
Scats were washed with tap water in a fine-mesh sieve (0.5 mm), and indigestible remains of the hair, teeth, hooves, bones, feathers and claws were oven dried at 60 °C for 24 h. Fragments of plastic or other man-made materials were eliminated from our analyses. Prey species were identified by comparison with a reference collection of slides and photographs of the cuticle (cuticula) and the medulla of the hair of potential prey species (Teerink 1991; Oli 1993), at × 100–× 400 magnifications. Hairs were washed in warm water with detergent, rinsed in distilled water, dehydrated in 70% ethanol and dried on filter paper. Twenty individual hairs were collected from each scat for identification. The hair was placed on a glass slide, on a layer of transparent nail polish, to observe the cuticle structure. After drying up the slide, the hair was removed and the cast was observed at the microscope (see above). The hair was longitudinally sectioned and wetted with cedar oil at the section level to observe the medullar pattern.
Data were tabulated as absolute (AF: number of occurrences of each food, when present/total number of scats × 100) and relative (RF: number of occurrences of each food, when present/total number of occurrences of all food items × 100) frequencies of occurrence of each prey species (Lucherini and Crema 1995). Seasonal differences in the absolute occurrences of main prey items in faecal samples were assessed through the G test. For each sample, the estimated relative volume of food ingested (estimated volume of each category/total number of scats × 100) was scored on a seven-point scale (absent, < 5, 6–25, 26–50, 51–75, 76–95, > 96%: Kruuk and Parish 1981). Relative frequencies and volumes were then plotted in a diagram (Kruuk 1989). The Levins’ standardised index (B STA) was used to assess the trophic niche breadth (Krebs 1999), B sta = (B − 1)/(B max − 1), where B is the Levins’ index (Levins 1968) and B max is the total number of prey categories. B sta varies between 0 (minimum breadth) and 1 (maximum breadth). The Pianka index (O jk) was used to assess the trophic niche overlap (Pianka 1974). Eight food categories were identified: domestic sheep, domestic goat, Siberian ibex (hereafter, ibex), cattle, yak (and hybrids with cattle), small mammals, birds, plant material and other. Occasionally, canids—but not felids—may use fruits as food, whereas grass blades are used as roughage, but they go undigested through the stomach and the intestines of both dogs and cats (cf. Bothma 1966; Hoppe-Dominik 1988). Accordingly, we have used the category ‘plant material’ only for the analysis of absolute frequencies, where the addition of this category does not affect the quantitative evaluation of the others. Conversely, we eliminated this indigestible category from relative frequency, volume and trophic niche overlap analyses. As to fruits, there was none during the cold period, in our study area. We have preferred not to estimate the biomass consumed, because of the substantial biases which may affect this calculation (cf. Chakrabarti et al. 2016; Lumetsberger et al. 2017) with further uncertainties, which make biomass estimation misleading, i.e. it is usually impossible to know (i) whether a young/sub-adult/male/female has been preyed upon (body mass is normally quite different in different age classes and sexes, especially of polygynous ungulates); (ii) whether a predator scavenged from a carcass already partly eaten by other carnivores or from its own fresh kill and (iii) whether it fed alone on it or with conspecifics. The selection of ungulate species as prey of snow leopards and Tibetan wolves was estimated through the comparison of the percentage of occurrence of each ungulate in the scats (i.e. use) with the availability of the same ungulate species in the study area. Prey (ungulate) selection was assessed through the Jacobs index (Jacobs 1974; Lovari et al. 2015).
Results
The Brillouin diversity index (Hass 2009; Lovari et al. 2015) suggested that our sample was large enough to assess the seasonal diet of both species in both seasons, as no new food category was found after 21 and 12 scats (winter) and 18 and 21 scats (spring), respectively, for the snow leopard and the wolf (Fig. 2).

Brillouin diversity index for the snow leopard (above) and the Tibetan wolf (below), for winter (left) and spring (right)
Diets of snow leopards and wolves differed significantly (SL: N = 74; W: N = 57; G = 26.87, degrees of freedom = 8, P < 0.01; Fig. 3), especially in winter (SL: N = 37; W: N = 17; G = 34.22, degrees of freedom = 6, P ≪ 0.01; Table 1).

Absolute frequencies of food categories in snow leopard and wolf diets during the cold period
Domestic sheep comprised the staple in the diet of the snow leopard, whereas goats and sheep were eaten by wolves in comparable numbers (Table 1; Fig. 3). Plant material was present in 69.9% of faecal samples of the snow leopard (mainly Myricaria rosea), as well as in 48.5% of the wolves (Table 1; Fig. 3). The trophic niche showed the same breadth for both species, in both seasons (SL: winter = 0.50, spring = 0.49; W: winter = 0.47, spring = 0.50). The Pianka index suggested a great overlap of trophic niche between snow leopards and wolves during the whole period (O jk = 0.74), as well as in winter (O jk = 0.67) and in spring (O jk = 0.77) respectively.
Ungulates (livestock and ibex) made up 85.8 and 82.8% in relative volume of the snow leopards and the wolves, respectively (Table 1; Fig. 4). Small mammals and birds were not present in the wolf’s diet and only rarely in the snow leopard’s (Table 1; Fig. 4). In winter, sheep was the category with the greatest total volume in the diet for the snow leopard, while goats had the highest volume in that of the wolf (Fig. 4). In spring, sheep showed the greatest volume in the diet for both carnivores, even if much more was consumed by wolves (Fig. 4).

Diet of the snow leopard and of the Tibetan wolf in winter (left) and spring (right). Isopleths connect points of equal relative volume
During the cold period, ungulates were eaten proportionally to their local availability by both snow leopards and Tibetan wolves (Table 2).
Discussion
An unexpected result of our study was the apparent lack of any selection, positive or negative, in the use of ungulate prey species by both predators. This is in contrast with the data provided by Jumabay-Uulu et al. (2013) and Chetri et al. (2017) who found a different selectivity, with the wolf concentrating on ungulates of undulating terrain and the snow leopard selecting cliff-dwelling ones. This difference could be due to the different composition of local prey spectra, livestock included. The ungulate assemblies in the studies of Jumabay-Uulu et al. (2013) and Chetri et al. (2017) were comparatively rich (8 prey species, in the former; 12 prey species, in the latter) compared to our study area (5 prey species). In our study, options for selection were less, due to the depauperate prey community because of human actions, which could explain why domestic ungulates were heavily used, while they were avoided in the other studies. This may support the expectation that predation on livestock increases when the wild ungulate community is depleted (e.g. Meriggi and Lovari 1996, for the wolf in South Europe, whereas results are unclear as to the snow leopard: Suryawanshi et al. 2013; Johansson et al. 2015; Sharma et al. 2015; cf. Lovari et al. 2013a, for a review). Predation by snow leopards and wolves on livestock was high in our study area compared to reports from other studies (Chundawat and Rawat 1994; Wang et al. 2014; Chetri et al. 2017), possibly because of the low availability of wild ungulates and the absence of alternative prey, e.g. marmots Marmota himalayana and Marmota baibacina (Khatoon 2010; Anwar et al. 2011; Lovari et al. 2013a; Lyngdoh et al. 2014). In Hushey valley, in the cold period, domestic livestock comprised 53.4 and 65.6% of the volume in the diet of the snow leopard and the wolf, respectively. The higher frequency of ibex in the diet of the former, with respect to that of the latter, could be due to the fact that ibex and leopards share the same terrain, whereas wolves seldom move into the steep terrain inhabited by ibex (Jumabay-Uulu et al. 2013).
In the cold period, both the snow leopard (Nawaz 2009) and the wolf (Mech 1970) descend to lower altitudes (< 3000 m.a.s.l.) where they may meet with livestock, an easy prey compared to wild mountain ungulates (Anwar et al. 2011). A puzzling result of our work is the differential predation of these carnivorous species on livestock (sheep/goats), during the cold period. On winter–early spring, especially in case of poor snow cover, goats tend to graze/browse at higher altitudes or on ridges where snow has been swept away by blowing winds, whereas usually, sheep remain close to the village. One might assume that the group-living wolf tends to prey on livestock where the probability of being detected is the lowest (i.e. far from the villages, should the opportunity arise), whereas the solitary snow leopard could rely more on its stealthy behaviour not to be detected, even if close to human habitations. With snow melting, on spring, broken terrain tends to be more available and goats attend it more than in winter, thus escaping predation by the wolf, which could explain this predator’s shift to sheep.
Alternative prey options such as small mammals and birds were absent in the diet of the wolf, whereas this category occurred in the snow leopard diet albeit rarely. In fact, smaller mammals may be used substantially by snow leopards when major prey species are not readily available (Chundawat and Rawat 1994). Plant material was more frequent in faecal samples of snow leopards than in those of wolves. As to the snow leopard, this category was mainly based on leaves and stems of the rosy false tamarisk Myricaria rosea (Dycotiledonae: Tamaricaceae), most likely as a way to clean the digestive tract, rather than as a food source (cf. Macdonald 1992; Lovari et al. 2013b).
The trophic niche breadths of these predators were comparable. A large diet overlap (up to 90%) has also been observed in another area, the Sarychat-Ertash, in Kyrgyzstan (Jumabay-Uulu et al. 2013), but not in the central Himalaya of Nepal (44%) where the ungulate community was particularly rich (Chetri et al. 2017). In the Hushey valley, the narrow niche breadth, the extensive overlap in diet and the apparent lack of prey selection would be expected to lead to food competition between these large carnivores. In fact, our data are limited to the colder months, when shortage of food is more likely to occur (Khan et al. 2014), thus enhancing potential competition.
Schaller (1977) outlined the vertical distribution of the main mammalian species in the north-west Himalaya, Karakoram and Hindu Kush. The snow leopard and the wolf have a widely overlapping altitudinal range, with the former between c. 1800 and 5000 m.a.s.l. and the latter from at least 1000 to over 5000 m.a.s.l. The greater biological flexibility of the wolf is confirmed, thus, both as to habitat type and to the use of local food resources (Mech 1970; Newsome et al. 2016). Conversely, the snow leopard appears to be much more of a ‘specialist’ species, strongly preferring open habitats and medium-sized wild herbivores (Sharma et al. 2015; Lovari and Mishra 2016). When an adaptable carnivore and a specialised one compete, most likely, the former will outcompete the latter (e.g. the red fox replacing the Arctic fox Vulpes lagopus: Tannerfeldt et al. 2002; for reviews on carnivore competition: Palomares and Caro 1999; Donadio and Buskirk 2006). A higher level of prey selection, with a minor risk of competition, should occur where the prey spectrum is rich (Butler et al. 2004; Wang et al. 2014). Conversely, where few prey species are observed and little or no prey selection occurs, interspecific competition is more likely to develop (cf. Stephens and Krebs 1986). In our study area, the outcome of a potential competition between the snow leopard and the wolf is difficult to predict. As the major food category is livestock, conflict with humans would drive consequences for both species more strongly than competition, as the availability of domestic prey is not limited in the same way as wild prey. Hence, most likely, competition for livestock would not escalate. Furthermore, in SW Asia, the coexistence of snow leopards and wolves is linked mainly to habitat type, with the leopard preferring cliffs and the wolf using less rugged, undulating terrain (Chetri et al. 2017). If herders start guarding livestock in a better way, the snow leopard could still prey on Siberian ibex, a cliff-dwelling species, whereas the wolf would be disadvantaged.
Overall, by contrast, current climatic changes have been suggested as detrimental to the survival of the snow leopard (Forrest et al. 2012; Lovari et al. 2013a; Aryal et al. 2016). In case of the development of competitive interactions, one could expect that this ‘vulnerable’ cold-adapted cat will be outcompeted by the ‘least-concern’ ecologically flexible wolf, an unfortunate event which would make its conservation even more precarious.
In both cases, the best way out could be primarily local habitat restoration, i.e. reforestation, control over domestic sheep/goat distribution, reintroduction of local prey species and poaching control, so that a balance could be found between these predators, who have coexisted for at least 1.2–1.4 Ma (Turner and Anton 1997) thus showing their ability to coexist in the same area.
References
Anwar MB, Jackson R, Nadeem MS, Janečka JE, Hussain S, Beg M, Muhammad G, Qayyum M (2011) Food habits of the snow leopard Panthera uncia (Schreber, 1775) in Baltistan, Northern Pakistan. Eur J Wildl Res 57(5):1077–1083. https://doi.org/10.1007/s10344-011-0521-2
Aryal A, Babu Shrestha U, Ji W, Ale SB, Shrestha S, Ingty T, Maraseni T, Cockfield G, Raubenheimer D (2016) Predicting the distributions of predator (snow leopard) and prey (blue sheep) under climate change in the Himalaya. Ecol Evol 6(12):4065–4075. https://doi.org/10.1002/ece3.2196
Bagchi S, Mishra C (2006) Living with large carnivores: predation on livestock by the snow leopard (Uncia uncia). J Zool (Lond) 268(3):217–224. https://doi.org/10.1111/j.1469-7998.2005.00030.x
Blumestein DT (1993) Dhee Sar—portrait of a high alpine meadow. WWF-Pakistan. Natura 18:4–7
Bothma JDP (1966) Food of the silver fox Vulpes chama. Zool Afr 2(2):205–210. https://doi.org/10.1080/00445096.1966.11447343
Butler JRA, du Toit JT, Bingham J (2004) Free-ranging domestic dogs (Canis familiaris) as predators and prey in rural Zimbabwe: threat of competition and disease to large wild carnivores. Biol Conserv 115(3):369–378. https://doi.org/10.1016/S0006-3207(03)00152-6
Chakrabarti S, Jhala YV, Dutta S, Qureshi Q, Kadivar RF, Rana VJ (2016) Adding constraints to predation through allometric relation of scats to consumption. J Anim Ecol 85(3):660–670. https://doi.org/10.1111/1365-2656.12508
Champion HG, Seth SK, Khattak GM (1965) Forest types of Pakistan. Pakistan Forest Institute, Peshawar
Chetri M, Odden M, Wegge P (2017) Snow leopards and Himalayan wolf: food habits and prey selection in the Central Himalayas, Nepal. Plos ONE 12(2):e0170549. https://doi.org/10.1371/journal.pone.0170549
Chundawat RS, Rawat GS (1994) Food habits of the snow leopard in Ladakh. In: Fox JL, Jizeng D (eds) International snow leopard trust and northwest plateau. Institute of Biology, Seattle, pp 127–132
Clemens J, Nusser M (1997) Resource management in Rupal Valley, Northern Pakistan: the utilization of forests and pastures in the Nanga Parbat area. In: Stellrecht I, Winiger M (eds) Perspectives on history and change in the Karakoram, Hindukush and Himalaya, vol 3. Culture Area Karakoram Scientific Studies, Cologne, pp 1–17
Donadio E, Buskirk SW (2006) Diet, morphology and interspecific killing in Carnivora. Am Nat 167(4):524–536. https://doi.org/10.1086/501033
Forrest JL, Wikramanayake E, Shrestha R, Areendran G, Gyeltshen K, Maheshwari A, Mazumdar S, Naidoo R, Thapa K (2012) Conservation and climate change: assessing the vulnerability of snow leopard habitat to treeline shift in the Himalaya. Biol Conserv 150(1):129–135. https://doi.org/10.1016/j.biocon.2012.03.001
Fox JL, Sinha SP, Chundawat RS, Das PK (1991) Status of snow leopard (Panthera uncia) in northwest India. Biol Conserv 55(3):283–298. https://doi.org/10.1016/0006-3207(91)90033-6
Hass CC (2009) Competition and coexistence in sympatric bobcats and pumas. J Zool (Lond) 278(3):174–180. https://doi.org/10.1111/j.1469-7998.2009.00565.x
Hoppe-Dominik B (1988) Grass-eating leopards: wolves turned into sheep? Naturwissenschaften 75(1):49–50. https://doi.org/10.1007/BF00367444
Jackson RM (1996) Home range, movements and habitat use of snow leopard (Uncia uncia) in Nepal. (Ph.D. Thesis) University of London (External Programme), London
Jacobs J (1974) Quantitative measurement of food selection-a modification of the forage ratio and Ivlev’s electivity index. Oecologia 14(4):413–417. https://doi.org/10.1007/BF00384581
Johansson O, McCarthy T, Samelius G, Andren H, Tumursukh L, Mishra C (2015) Snow leopard predation in a livestock dominated landscape in Mongolia. Biol Conserv 184:251–258. https://doi.org/10.1016/j.biocon.2015.02.003
Jumabay-Uulu K, Wegge P, Mishra C, Sharma K (2013) Large carnivores and low diversity of optimal prey: a comparison of the diet of snow leopard Panthera uncia and wolf Canis lupus in Sarychat-Ertash Reserve in Kyrgyzstan. Oryx 48:529–535
Khan B (2012) Pastoralism-Wildlife Conservation Conflict in Climate Change Context—a study of climatic factors influencing fragile mountain ecosystems and pastoral livelihoods in the Karakoram Pamir trans-border area between China and Pakistan. Ph.D thesis, Graduate University of the Chinese Academy of Sciences, China
Khan MZ, Awan MS, Bocci A, Khan B, Abbas SY, Khan G, Abbas S (2014) Population structure and grouping tendency of Asiatic ibex Capra sibirica in the Central Karakoram National Park, Pakistan. J Bio Env Sci 5:542–554
Khan MZ, Khan B, Awan MS, Begum F (2017) Livestock depredation by large predators and its implications for conservation and livelihoods in the Karakoram Mountains of Pakistan. Oryx 1–7. https://doi.org/10.1017/S0030605316001095
Khatoon R (2010) Diet selection of snow leopard Uncia uncia in Chitral Area. Master Thesis. Department of Wildlife Management, Faculty of Forestry, Range Management and Wildlife, Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi
Krebs CJ (1999) Ecological methodology, 2nd edn. Addison Wesley Longman, Menlo Park
Kruuk H (1989) The social badger: ecology and behaviour of group-living carnivore (Meles meles). Oxford University Press, Oxford
Kruuk H, Parish T (1981) Feeding specialization of the European badger Meles meles in Scotland. J. Anim Ecol 50:773–788
Lescureux N (2006) Towards the necessity of a new interactive approach integrating ethnology, ecology, and ethology in the study of the relationship between Kyrgyz stockbreeders and wolves. Soc Sci Inform 45:463–478
Levins R (1968) Evolution in changing environments. Princeton University Press, Princeton
Lovari S, Mishra C (2016) Living on the edge: depletion of wild prey and survival of the snow leopard. In: McCarthy T, Mallon D (eds) Snow leopards. Elsevier Academic Press, Amsterdam, pp 69–76
Lovari S, Boesi R, Minder I, Mucci N, Randi E, Dematteis A, Ale SB (2009) Restoring a keystone predator may endanger a prey species in a human-altered ecosystem: the return of the snow leopard to Sagarmatha National Park. Anim Conserv 12(6):559–570. https://doi.org/10.1111/j.1469-1795.2009.00285.x
Lovari S, Ventimiglia M, Minder I (2013a) Food habits of two leopard species, competition, climate change and upperline: a way to the decrease of an endangered species? Ethol Ecol Evol 25(4):305–318. https://doi.org/10.1080/03949370.2013.806362
Lovari S, Minder I, Ferretti F, Mucci N, Randi E, Pellizi B (2013b) Common and snow leopards share prey, but not habitat: competition avoidance by large predators? J Zool (Lond) 291(2):127–135. https://doi.org/10.1111/jzo.12053
Lovari S, Pokheral CP, Jnavali SR, Fusani L, Ferretti F (2015) Coexistence of the tiger and the common leopard in a prey-rich area: the role of prey partitioning. J Zool (Lond) 295(2):122–131. https://doi.org/10.1111/jzo.12192
Lucherini M, Crema G (1995) Seasonal variation in the food habits of badgers in an Alpine valley. Hystrix 7:165–171
Lumetsberger T, Ghoddousi A, Appel A, Khorozyan I, Waltert M, Kiffner C (2017) Re-evaluating models for estimating prey consumption by leopards. J Zool (Lond) 302(3):201–210. https://doi.org/10.1111/jzo.12449
Lyngdoh S, Shrotriya S, Goyal SP, Clements H, Hayward MW, Habib B (2014) Prey preferences of the snow leopard (Panthera uncia): regional diet specificity holds global significance for conservation. PLoS One 9(2):e88349. https://doi.org/10.1371/journal.pone.0088349
Macdonald DW (1992) The velvet claw: a natural history of carnivores. BBC Book, London
Mech LD (1970) The wolf: the ecology and behaviour of an endangered species. Natural History Press, New York
Meriggi A, Lovari S (1996) A review of wolf predation in southern Europe: does the wolf prefer wild prey to livestock? J Appl Ecol 33(6):1561–1571. https://doi.org/10.2307/2404794
Mishra C (1997) Livestock depredation by large carnivores in the Indian trans-Himalaya: conflict perceptions and conservation prospects. Environ Conserv 24(4):338–343. https://doi.org/10.1017/S0376892997000441
Namgail T, Fox JL, Bhatnagar YV (2007) Carnivore-caused livestock mortality in trans-Himalaya. Environ Manag 39(4):490–496. https://doi.org/10.1007/s00267-005-0178-2
Nawaz MA (2009) Draft management plan for CKCC; sub-plan Species Management. IUCN, Karachi 24pp
Newsome TM, Boitani L, Chapron G, Ciucci P, Dickman CR, Dellinger JA, Lòpez-Bao JV, Peterson RO, Shores CR, Wirsing AJ, Ripple WJ (2016) Food habits of the world’s grey wolves. Mammal Rev 46(4):255–269. https://doi.org/10.1111/mam.12067
Oli MK (1993) A key for the identification of hair of mammals of a snow leopard (Panthera uncia) habitat in Nepal. J Zool (Lond) 231(1):71–93. https://doi.org/10.1111/j.1469-7998.1993.tb05354.x
Oli MK, Taylor IR, Rogers ME (1993) The diet of snow leopard (Panthera uncia) in the Annapurna Conservation Area, Nepal. J Zool (Lond) 231(3):365–370. https://doi.org/10.1111/j.1469-7998.1993.tb01924.x
Palomares F, Caro TM (1999) Interspecific killing among mammalian carnivores. Am Nat 153(5):492–508. https://doi.org/10.1086/303189
Palomares F, Ferreras P, Fedriani JM, Delibes M (1996) Spatial relationships between the Iberian lynx and other carnivores in an area of south-western Spain. J Appl Ecol 33(1):5–13. https://doi.org/10.2307/2405010
Peiman K, Robinson B (2010) Ecology and evolution of resource-related heterospecific aggression. Q Rev Biol 85(2):133–158. https://doi.org/10.1086/652374
Pianka ER (1974) Niche overlap and diffuse competition. Proc Nat Ac Sci 71(5):2141–2145. https://doi.org/10.1073/pnas.71.5.2141
Reynolds J, Aebischer NJ (1991) Comparison and quantification of carnivore diet by faecal analysis: a critique, with recommendations, based on a study of red fox Vulpes vulpes. Mammal Rev 21(3):97–121. https://doi.org/10.1111/j.1365-2907.1991.tb00113.x
Schaller GB (1977) Mountain monarchs. Wild sheep and goats of the Himalaya. The University of Chicago Press, Chicago
Schaller GB (2016) Foreword. In: McCarthy T, Mallon D (eds) Snow leopards. Elsevier Academic Press, Amsterdam, pp 23–27
Sharma RK, Bhatnagar YV, Mishra C (2015) Does livestock benefit or harm snow leopards? Biol Conserv 190:8–13. https://doi.org/10.1016/j.biocon.2015.04.026
Sheikh KM, Molur S (2004) Status and red list of Pakistan mammals. Conservation assessment and management plan. IUCN Pakistan, Karachi
Shrestha B (2008) Prey abundance and prey selection by snow leopard (Uncia uncia) in Sagarmatha (Mt. Everest) National Park, Nepal. Report for International Snow Leopard Trust, Snow Leopard Conservancy and snow leopard network, Forum of Natural Resource Managers, Nepal, 34pp
Stephens DW, Krebs JR (1986) Foraging theory. Princeton University Press, Princeton
Sugimoto T, Aramilev VV, Nagata J, McCullough DR (2016) Winter food habits of sympatric carnivores, Amur tigers and Far East leopards, in the Russian Far East. Mammal Biol 81(2):214–218. https://doi.org/10.1016/j.mambio.2015.12.002
Suryawanshi KR, Bhatnagar YV, Redpath S, Mishra C (2013) People, predators and perceptions: patterns of livestock depredation by snow leopards and wolves. J Appl Ecol 50(3):550–560. https://doi.org/10.1111/1365-2664.12061
Tannerfeldt M, Elmhagen B, Angerbjörn A (2002) Exclusion by interference competition? The relationship between red and arctic foxes. Oecologia 132(2):213–220. https://doi.org/10.1007/s00442-002-0967-8
Teerink BJ (1991) Hair of West-European mammals. Cambridge University Press, Cambridge
Turner A, Anton M (1997) The big cats and their fossil relatives: an illustrated guide to their evolution and natural history. Columbia University Press, New York
Ud Din J, Ali H, Ali Nawaz M (2016) The current state of snow leopard conservation in Pakistan. In: McCarthy T, Mallon D (eds) Snow leopards. Elsevier Academic Press, Amsterdam, pp 486–492
Wang J, Laguardia A, Damerell PJ, Riordan P, Shi K (2014) Dietary overlap of snow leopard and other carnivores in the Pamirs of Nortwestern China. Chin Sci Bull 59(25):3162–3168. https://doi.org/10.1007/s11434-014-0370-y
Watts HE, Holekamp KE (2008) Interspecific competition influences reproduction in spotted hyenas. J Zool (Lond) 276(4):402–410. https://doi.org/10.1111/j.1469-7998.2008.00506.x
WWF-Pakistan (2008) Land cover mapping of the Central Karakoram National Park. WWF Pakistan, Lahore
Acknowledgements
We are greatly indebted to Agostino Da Polenza and all the staff of Ev-K2-CNR for financial support and help with logistics. The personnel of CKNP, as well as Musa Alì and especially the Wildlife Watcher Sakhawat Alì, helped in fieldwork. We are also grateful to Erica Ercoli, who analysed a part of the scats for diet assessment, as well as to Francesco Ferretti and Luca Rossi for reading critically a previous draft of this manuscript. David M. Shackleton kindly revised our English and provided suggestions. Two anonymous referees improved our final draft with useful comments.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Bocci, A., Lovari, S., Khan, M.Z. et al. Sympatric snow leopards and Tibetan wolves: coexistence of large carnivores with human-driven potential competition. Eur J Wildl Res 63, 92 (2017). https://doi.org/10.1007/s10344-017-1151-0
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-017-1151-0