
Abstract—A field survey of snow leopard (Panthera uncia) habitats was carried out in the southeastern part of the Eastern Sayan Mountains (Okinskii and Tunkinskii districts of the Republic of Buryatia and the Kaa-Khemskii district of Tuva Republic). Seven or eight adult snow leopards were observed as constant inhabitants of the Tunkinskie Gol’tsy, Munku-Sardyk, and Bol’shoi Sayan mountain ridges. The presence of eight snow leopards was confirmed using DNA-based analyses of scats collected in 2014–2016. The main prey species of the snow leopard in Eastern Sayan is the Siberian ibex (Capra sibirica), but its abundance has steadily decreased over the past 20 years. The red deer (Cervus elaphus) and the wild boar (Sus scrofa), which were some of the most numerous ungulates in the survey area, are replacing the Siberian ibex in the snow leopard’s diet. In addition, the mountain hare (Lepus timidus) is also of importance to the snow leopard’s diet.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
INTRODUCTION
The state of populations of some animal species is constantly changing as a result of increasing human impact on nature, so that data on them quickly become obsolete. An indirect assessment of the state of a population or a species as a whole can be achieved through estimation of the stability of the boundaries of ranges, as ranges change and become fragmented, along with the disappearance of some groups, and new groups previously unreported or existing sometime in the past can appear. It should be noted that at the border of a range, the distribution of a species can change for natural reasons as well, due to the occurrence of more extreme conditions than in the main part of the range. For rare species of animals, data on their distribution is relevant and important both from the theoretical point of view and from the practical one, which allows researchers to develop scientifically grounded recommendations for preservation of the species.
A fundamentally new approach to studying the distribution of animals in recent years has become modeling of their potential habitats. The Maxent maximum entropy method has proved particularly successful (Elith et al., 2006, 2011). This method is an algorithm of machine learning and is used to model the distribution of maximum entropy (Maxent). The probability of the presence of a species in a particular pixel of the modeled area is determined from the known points of this species. In this case, the data set is expressed as simple functions of environmental variables, called “features” (Phillips and Dudík, 2008).
The snow leopard (Panthera uncia) is one of the rarest and least studied large predators. Being at the top of the food pyramid, it can serve as an indicator of the state of the entire ecosystem. The Russian part of the snow leopard’s range is the northern outskirts of the species distribution. The mosaic nature of the area, the inaccessibility of its habitats, and its secretive way of life create significant obstacles to studying this species. Given the steady tendency toward a reduction in the number and a decrease in the historically established range of snow leopards, studying it (and in particular, specifying the modern state of individual groupings) appears to be very relevant.
Despite relatively active study of the snow leopard recently, the eastern part of its range, especially the Eastern Sayan, has not been covered by studies using modern methods (camera traps, genetic methods, and modeling). In the Eastern Sayan, the snow leopard’s habitation was noted in the Tunkinskie and Kitoiskie Gol’tsy (Geptner and Sludsky, 1972) and on the Ergak-Targak-Taiga and Udinskii ridges (Nikiforov and Shurygin, 1977; Smirnov et al., 1991). However, the mentions of these authors were based, first of all, on survey information. The first reliable data on the distribution of snow leopards in the Eastern Sayan were obtained by Koshkarev in 1995–1997 (Koshkarev, 1996, 1997). Several special expeditions aimed at finding traces of life of the snow leopard were carried out. Later, the results of Medvedev’s research, which actively used the survey method (Medvedev and Trofimov, 2001; Medvedev, 2003), confirmed these data and partially expanded the range of the snow leopard in Eastern Sayan.
One of the most important signs of the presence of the snow leopard in the study area is the presence of scrapes, as they can persist for quite a long time. It is their presence and the frequency of sightings that are accepted as the most important indicators of the state of the population in the SLIMS method (Jackson and Ahlborn, 1987, unpublished; Hunter and Jackson, 1997). No less important an indicator is also the occurrence of excrement, but during their identification, confusion often occurs: the molecular-genetic analysis we conducted showed that among the previously collected Southwestern Tuva samples of excrement, a large part belonged to the fox (Vulpes vulpes) (Rozhnov et al., 2011). A good indicator of the state of a population can be the occurrence of tracks, but only in the winter period of time, since in the snowless period of the year finding tracks is very difficult, if at all possible.
The purpose of our work was a field survey of the habitats of the Eastern Sayan snow leopard grouping to assess the state of its population and its nutritive base using modern methods, as well as identification of potential habitats of the species by the maximum entropy method.
MATERIALS AND METHODS
Studies were conducted in the eastern, most high-elevation, and fragmented part of the Eastern Sayan: on the territory of the Okinskii and Tunkinskii districts of the Republic of Buryatia and the Kaa-Khemskii district in the Republic of Tuva. Modern methods (camera trapping, molecular diagnostics of excrement) have been used in the Tunkinskie Gol’tsy since 2011, along the Bol’shoi Sayan Ridge, and for most of Munku-Sardyk Ridge in 2014–2015.
Route studies covered Tunkinskie Gol’tsy (583.7 km, 51 registration routes), the Munku-Sardyk Ridge (1360.68 km, 92 routes), the western part of the Bol’shoi Sayan (1528.53 km, 107 routes), and Kropotkinskii Ridge (83.4 km, 18 routes). The census routes were drawn in such a way that the probability of finding traces of life of the snow leopard was maximum: in the summer time, we chose well-marked ridges of mountains and side branches of ridges; in winter time, work was also conducted on river beds. The length of the survey route averaged 13.27 km. In total 268 routes were passed, and their total length was 3556.31 km.
All snow leopard signs found on the routes (scrapes, footprints, urine marks, tear marks on trees) were recorded in the GPS navigator and plotted onto a topographic map using the OziExplorer program. The Garmin GPSMAP 60CSx, Garmin GPSMAP 62S, and Garmin eTrex30 GPS navigators were used. The found feces were sampled, fixed, and sent for subsequent molecular genetic analysis.
In a number of places near traces of life of the snow leopard and on the main routes of its passages, automatic photo and video recorders Reconyx RC60 RapidFire, Reconyx HC600 HyperFire (RECONYX, Inc., United States), Bushnell Trophy Cam HD Max and Bushnell Trophy XLT (Bushnell Outdoor Products, United States), SG560K8mHD were installed. Their location was marked with a GPS navigator (Jackson et al., 2010). In total, 43 photo-traps were installed: 8 in Tunkinskie Gol’tsy, 16 on the Munku-Sardyk Ridge, and 19 on the Bol’shoi Sayan. For photographic registration, the following parameters were used: “image quality: maximum,” “sensitivity: minimum,” other parameters were not changed. Owing to the poor accessibility of the research area, the photo traps were checked 1–2 times a year.
For 47 samples of excrement collected on Tunkinskie Gol’tsy (n = 11), Munku-Sardyk (n = 14), and Bol’shoi Sayan (n = 22), molecular diagnostic methods were used for their identification. At the first stage of the molecular genetic analysis, their species identity was determined by sequencing the fragment of the cytochrome b mtDNA gene. At the second stage, individual identification of samples belonging to the snow leopard was performed, for which each sample was genotyped at eight microsatellite loci, and by two loci localized in the sex chromosomes, the gender of the animals was determined (Rozhnov et al., 2011). As comparative data, the genotypes of snow leopards from the Tsagan-Shibetu mountain range in the Republic of Tuva (n = 17) were used, as well as from Sayano-Shushenskii Nature Reserve in Krasnoyarskii krai (n = 7) and from the territory of the Tsagaan Shiveetiin Nuruu mountain massif in northwestern Mongolia (n = 19) (our data).
Modeling of the potential habitats of the snow leopard was carried out using the maximum entropy method (Maxent) (Elith et al., 2006, 2011). The modeling area covers the region in which we performed intensive field work in 2014–2016, as well as a significant part of the adjacent territory of Mongolia and is located between the following coordinates: northwestern corner 52.470500° N, 98.473600° E and the southeastern corner 50.579300° N, 102.659400° E (i.e., the modeling area included the eastern part of the range of snow leopards, including Buryatia, the adjacent eastern parts of Tuva, and the northern regions Mongolia in the vicinity of Khuvsgul Lake).
The primary analysis included 161 discrete points of snow leopards’ presence points; these points were obtained in 2014–2015. In those pixels where there were no basic discrete points, 800 points were added, obtained during the tracking of the snow leopard. The presence of the snow leopard in all the points is documented (camera trap data, genetically confirmed samples of excrements, scrapes and footprints). To minimize distortions and overlapping of data with environmental indicators during further analysis, WGS 1984 datum coordinates were converted into a conical equal-sized Albers projection for Siberia. All calculations were carried out in the software environment R (version 3.0.1), packages: spatstat (Baddeley and Turner, 2005), rgdal (Bivand et al., 2013), adehabitatHR (Calenge, 2006), maptools (Bivand and LewinKoh, 2013), and SP (Pebesma and Bivand, 2005).
To model the potential habitats, 15 variables characterizing each point related to three different types—climatic, physical-geographical (including types of plant associations), and infrastructure—were used. The type of the climatic variables included (data for different periods): snow cover for the period of 2000–2011 (average monthly and maximum); type of cover (e.g., snow, water, ice or bare ground) for the period of 2006–2011 (monthly); the average monthly temperature and precipitation. The type of the physical-geographical variables (which are considered constant in time) included the height, slope, exposure, index of topographic fragmentation, distance to water sources, structure of the woody vegetation and type of forest. The infrastructure type was represented by the distance from the presence points of snow leopards to roads and settlements.
All the variables were obtained for the whole world or Eurasia in the form of a table in the conical equal-sized Albers projection for Siberia. Before the analysis in Maxent, they were converted into ASC files with a unified cell using the Geospatial Data Abstraction Library (GDAL).
By default, Maxent executes 10 000 runs per pixel of the entire area analyzed (Phillips et al., 2009). We used 5000 runs, which is sufficient to obtain an adequate model (Young et al., 2011). For estimating the predictive power of the models of the snow leopard distribution, we used cross-valuation. The data was divided into 15 equal samples, of which 14 samples were used for training, and one sample, randomly selected as the control, created semi-independent data sets, different for each of the 15 independent launches. Thus, during the replication of Maxent’s work, we used all the data to check the reliability of the prognosis and give us the opportunity to measure the amount of variability in the model (Young et al., 2011). The remaining Maxent parameters were left by default, preconfigured and tested in a wide range of data (Phillips and Dudík, 2008).
The algorithm for the output of the results gives an estimate of the probability of the presence of the species throughout the areas analyzed in the range from 0 to 1. The zones with a probability of 0.5 and higher were considered as the “key” habitats of animals (in which the snow leopard was found with a probability of 50% or higher), and a probability between 0.49–0.25 was attributed to the zone of “transit” habitats. Transit habitats can be considered as ecological corridors connecting the main habitats of the snow leopard.
RESULTS
The current status of the Eastern Sayan grouping of snow leopards. For the entire period of work in 2014–2016 on the registration routes in Eastern Sayan, we recorded 69 scrapes, 50 samples of excrement, 14 tracks, and 12 urine marks of snow leopards (Table 1). In addition, three marking trees were found. Almost all snow leopards signs were confined to the ridges of mountains and side branches of ridges. In winter, traces of snow leopards were also noted in the valleys of rivers on ice.
For the 30 samples of excrement collected, using fragment sequence analysis of the cytochrome b gene of mtDNA, their species identity was determined: 21 specimens belonged to the snow leopard, four samples to the lynx, two samples to the wolf, and one sample each to the Pallas’ cat, wolverine, and fox. Microsatellite analysis made it possible to provide the individual identification of 12 samples of snow leopard feces, of which the genotypes were described for eight individuals: two males from Tunkinskie Gol’tsy, one male and one female from the Munku-Sardyk mountain Ridge, and one male and three females from Bol’shoi Sayan.
The results of field research allow us to divide the range of the snow leopard in the Eastern Sayan conditionally into several sites which the predator inhabits continuously: Tunkinskie Gol’tsy, Munku-Sardyk, and Bol’shoi Sayan. On Kropotkinskii Ridge, we did not observe the snow leopard signs for two years, but the survey data in that place only indicate its periodic visits.
In Tunkinskie Gol’tsy, the snow leopard has been constantly observed from 1999 to the present. Most often traces of its vital activity are recorded in the central and western parts of the ridge, in the upper reaches of the Ekhe-Ugun and Ekhe-Ger (southern macroslope) and Ara-Khongoldoi and Arkhat (northern macroslope) rivers. To the east of the Iltykshin River, the snow leopard is seen less often, and further to the east, according to polling data, only temporary visits are noted. We did not conduct registration routes in Kitoiskie Gol’tsy. Surveys show that the snow leopard used to live here permanently, but at present traces of it are less common than in Tunkinskie Gol’tsy, and its presence is of a temporary nature. In 2016 in the basin of the river, the research team of Medvedev with the help of a photo trap registered for the first time a specimen of the snow leopard, formerly marked on the ridge of Munku-Sardyk (the distance between registration points is about 100 km in a straight line). We, using photo-traps and molecular-genetic analysis of the excrement, revealed the habitat in Tunkinskie Gol’tsy of two adult males of the snow leopard. In addition, in autumn 2015 at the confluence of the Right and Left Shumak rivers, tourists watched a female snow leopard, presumably with cubs.
On Munku-Sardyk Ridge, snow leopards are constantly observed. This mountain range is of great importance for the species: in the eastern part it forms a narrow, of just a few kilometers, migration corridor in Tunkinskie Gol’tsy, where the animals cannot come with other territories. We constantly registered the snow leopard signs in the Munku-Sardyk mountain range, but less often than in the most developed west-central part of the Tunkinskie Gol’tsy Range. In the western part of Munku-Sardyk Ridge, the first time we surveyed in August–September 2015, snow leopards signs of various degrees of recognizability were found. Polls indicate the historical presence of this species in the given territory. At the junction of Tunkinskie Gol’tsy and the Munku-Sardyk mountain massif, in the upper reaches of the Irkut River, during the winter season, there were regular crossings of the snow leopard from one mountain system to another. Currently, such evidence is very rare, which reflects the rarity of the snow leopard. Snow leopard signs were noted by us in the upper reaches of the Muguwek, Boghuvek, Belyi Irkut, Zhokhoi, Butu-Gol, and Horyo rivers. As of the end of 2016 on the ridge, Munku-Sardyk was inhabited by one adult male of snow leopard and one female with two cubs (this is the first reliable information about the presence here of breeding females), which is confirmed by the molecular analysis of collected samples of excrement, and the sex of another adult animal is not established. Thus, on the ridge of Munku-Sardyk, at least three adult snow leopards live.
Bol’shoi Sayan is a vast mountainous area, a node occupying the central part of the range of the East Sayan grouping of the snow leopard. The most common traces of this life were noted at the sources of the Sorkhoi, Kharaganta, and Sagan-Gol rivers. Here, apparently, the nucleus of the Eastern Sayan grouping is located, so this area has an importance for the conservation of snow leopards in the eastern part of its range. In the course of recording the snow leopard in February–March 2016 on the Bol’shoi Sayan Ridge in the headwaters of the Zabit, Sorkhoi, Kharaganta, and Sagan-Gol rivers, we marked in the footprints the presence of 1–2 adult males and one adult female with two cubs. One male and female with cubs were also recorded in camera traps, and molecular-genetic analysis confirmed the dwelling of one male and three females (the cubs, apparently, are females). Thus, 4–5 snow leopards permanently inhabit this area.
An extensive plateau, located east of the Sorkhoi River, is unsuitable for snow leopards, as it is more populated and often visited by people. The terrain lying to the west, in the basin of the Obogol River, is also laid out, and the snow leopard lives here only along the axial ridge of the Bol’shoi Sayan, at the sources the Zun Obogol and Barun Obogol rivers. In the contiguous parts of Tuva, at the sources of the Bilin River, we also found snow leopards signs, but this area remained insufficiently surveyed. The left bank of the Tissa River and the river basin of the Sentsa River, lying to the north, are of little use for a permanent habitat of snow leopards: traces here, according to polling data, are noted only occasionally. On the Okinskoe plateau at the watershed of the Tissa and Sentsa rivers, snow leopards are forced to cross at the transition from the central part of the area of the grouping (right bank of the Tissa) to the western part at the middle reaches of the Zhombolok River (southern extremity of the Kropotkinskii Range): the basins of the Bursag and Hadarus rivers, as well as watersheds of the Barun Khadarus and Zun Hadarus rivers and the mountain ranges adjoining them, are suitable for the habitat of the species. However, during the census of the snow leopard in February–March 2016, we did not find snow leopard signs on the indicated territory. According to the survey data, there are occasional visits of 1–2 individual snow leopards. Further northwest, the presence of snow leopards in the Eastern and Central Sayan Mountains acquires a temporary character; here only occasionally does one find information about the approaches of single animals. It can be associated with a much deeper snow cover, which greatly hinders the movement of predators and forces the migration of ungulates, which form the basis of the snow leopard diet. However, it should be noted that during the last ten years in the Central Sayan no survey was conducted.
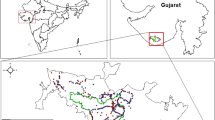
As a result of modeling potential habitats conducted for the whole territory, which is inhabited by the Eastern Sayan grouping of snow leopards and on which we conducted intensive field work during 2014–2016, as well as for a significant part of the adjacent territory of Mongolia, we identified key transit habitats (Fig. 1). The area of key habitats was 13 812 km2, and including Russia, it is 11 625 km2. The area of transit habitats is 62 499 km2 in general for the analyzed territory, including 41 877 km2 in Russia.

The map of potential habitats (key and transit ones) in the eastern part of the snow leopard’s range in Russia.
The central part of the Bol’shoi Sayan in the upper reaches of the Sorkhoi, Kharaganta, and Sagan-Gol rivers are the main stretch of the range of the Eastern Sayan grouping. The second most important site is Munku-Sardyk, the mountain massif itself and the tip adjoining it of the Tunkinskie Gol’tsy and the Bol’shoi Sayan (the upper reaches of Butugol and Zhokhoi). An equally important area is Tunkinskie Gol’tsy. The area of inhabited habitats of snow leopards lying to the north of Kitoiskie Gol’tsy much less than in Tunkinskie Gol’tsy: the presence of the snow leopard is mainly of an accidental, transitory nature, although here also key habitats are identified.
Modeling clearly identified areas with an unusual habitat of the snow leopard, the Sentsa River basin and Bol’shoi Sayan Ridge from the Zabit River up to the Horyo River (a well-watered and heavily waterlogged region separating the grouping of the central part of the Bol’shoi Sayan and the Munku-Sardyk grouping), the Tunkinskaya hollow (Irkut River basin), separating the Tukninskie and Kitoiskie Gol’tsy and the western spurs of the Bol’shoi Sayan, from Sagan-Gol to Tuva and further (Obogol and Bilin river basins, etc.). The areas of Tuva in our analysis were also of little use for the snow leopard.
Thus, the territory of Buryatia, Mongolia, and the adjoining part of Tuva is a habitat for a single transboundary grouping of the snow leopard, in which three main areas connected with each other form a permanent habitat, the Tunkinskie Gol’tsy, Munku-Sardyk, and Bol’shoi Sayan ranges. This grouping is represented by 7–8 adult individuals (4–5 males, 2 females, and one animal the sex of which is not defined) and four young individuals (in 2016 two broods of snow leopard were documented having two cubs in each of them); i.e., its total strength is 11–12 individuals. In addition, according to census data, in Tunkinskie Gol’tsy, another female with cubs may live. Inhabits of several individuals are also possible in the upper reaches of the Bilin and Barun Obogol rivers on the Bol’shoi Sayan and in the Kropotkinskii Ridge, which have not been adequately surveyed. According to our observations, in the past 10–15 years there has been a downward trend in numbers snow leopard, which is confirmed by survey data, known cases of obtaining snow leopard, and the deterioration of the population its feed objects.
The genetic characteristics of the Buryatia group of snow leopards indicate a low level of its variability in comparison with other groups from the northern part of the species range, which manifests itself both in the level of heterozygosity (He and Ho), and in the values of allelic diversity (the average number of alleles per locus) (Table 2). Analysis of the genetic differences based on the FST criterion made it possible to identify a significant genetic differentiation between Buryat animals and other samples (Table 3). The clear pattern of isolation of groups of snow leopard from Buryatia and from the Sayano-Shushenskii Reserve is demonstrated in statistically confirmed genetic differences from all analyzed samples. At the same time, the Buryat and other samples revealed a larger share of the average intergroup variability than between the animals from the Sayano-Shushenskii Reserve and the rest (0.209 and 0.181, respectively), which can speak about the greatest genetic differences of the first.
The state of the nutritive base of the snow leopard. The condition of the food base of the snow leopard in the Eastern Sayan raises serious concerns. This applies to all types of possible diets of snow leopards.
The Eastern Sayan population of the Siberian ibex (Capra sibirica), which is the main object of feeding of the snow leopard (Heptner and Sludsky, 1972), occupies in Buryatia an area of 12 000–15 000 km2. The distribution of ibex has a focal character, which determines the low overall density across the territory. In recent years, the Siberian ibex has greatly decreased in number. This has been noted not only by us on the routes, but by all respondents. In the summer, ibexes in small groups and singly are scattered throughout the study area and are extremely rare.
In the Tunkinskii district of Buryatia for the past ten years, the number of ibex has declined more than twice. Very revealing survey results come from dozens of respondents among local residents, in which the following tendency was traced: in the early 2000s, “little ibex,” and in recent years, “little ibex.” The situation is more acutely reflected in surveys in the Okinskii district of Buryatia: in the early 2000s, “the ibex was a rampart,” and now “there is no ibex.” On Kropotkinskii Ridge, for example, the decline in numbers is characterized from 200–300 to 30–40 individuals. The same situation is observed in Bol’shoi Sayan Ridge, where on the same routes for a month of work in the summer of 2015 we had only four encounters with ibex, and in the summer of 2016, five. On the ridge of Munku-Sardyk for the 100 km routes in the summer of 2015, there were no ibexes noted, only traces, and in the summer of 2016, only two groups. In Tunkinskie Gol’tsy on 100 km of routes in the summer in 2014–2016, only one group of ibex was seen.
On the entire territory of Tunkinskie Gol’tsy and the Bol’shoi Sayan, ibex were previously estimated at 200–500 individuals, with a clear decrease in its numbers. The size of herds fell to a minimum and is an average of 3–5 individuals. Groups of the Siberian ibex of 7–10 individuals are been noted at all.
The total number of ibex in Buryatia still has not been clarified to the proper extent, and its estimates vary greatly. For example, for Tunkinskie Gol’tsy the spread of estimates is more than fivefold: from 280 to 1500 individuals (Medvedev, 2000). In the eastern part Eastern Sayan Ridge there is also observed a spread of 3–5 times in the estimation of the number of ibex, from 500–950 individuals (Krasnaya kniga Respubliki Buryatiya..., 2005) to 2400–3000 individuals (Medvedev, 2000). By our estimate, the total number of the Siberian ibex in Buryatia does not exceed 1000 individuals. It should be noted that this is a very approximate assessment, since high-quality research on the mountain goat over such an extensive territory has never been carried out.
The roe deer (Capreolus pygargus) in the study area is even rarer, so a large value in the snow leopard cannot be fed.
The Siberian musk deer (Moschus moschiferus) inhabits the upper boundary of the forest with the approaches to the pelvic zone and may be the object of food. The snow cover has a significant impact on the conditions of its existence, as well as suitable relief in the zone of the upper boundary of the forest visited by snow leopard. According to our observations, the most favorable for the musk deer is Tunkinskie Gol’tsy, as evidenced by the occurrence of its excrement on the routes, and less favorable is Bol’shoi Sayan as it is more snow-white. An uncontrolled hunting press keeps the density of the musk deer low. In such conditions musk deer in the area of study for snow leopards is accidental prey.
Red deer (Cervus elaphus) is the most numerous of ungulates in the Eastern Sayan, but in accessible and frequently visited areas (Tunkinskie Gol’tsy and the eastern part of Munku-Sardyk) visually here it is practically not observed, as it is exposed to a constant press of hunting. The largest accumulations of red deer are recorded on Bol’shoi Sayan, where this species is likely to make up a significant portion of the snow leopard diet. On Bol’shoi Sayan red deer were noted by us visually almost daily during work, as single animals and as groups of up to 11 individuals. They inhabit not only the river valleys and the taiga zone, but also loaches. We have repeatedly encountered their footprints in the highlands on the ridges and on passes. During work in 2014–2016, we visually observed 36 red deer; they were all encountered only in the upper part of the basins of the Sorkhoi, Kharaganta, Sagan-Gol, and Zun-Obogol rivers. Traceability of tracks on routes varied between 5–24, and the total number of intersections was 131 trails of the deer.
In the Bol’shoi Sayan, boars are also common (Sus scrofa). During the fieldwork boars were visually encountered only five times, however traces of their habitation are fixed on almost all routes.
Of the smaller animals, the mountain hare (Lepus timidus) can play a serious role in nutrition, which was highlighted almost everywhere on all routes. Apparently, it is this species at present that is the most important fodder resource for the snow leopard in Buryatia.
DISCUSSION
The data obtained on the distribution of the snow leopard population in the Eastern Sayan are consistent with previously published data (Koshkarev, 1996, 1997; Poyarkov et al., 2002). For the first time, an estimate was made of the abundance of the East Sayan grouping of the snow leopard with the help of modern methods of research (molecular analysis, camera trapping) and traditional route surveys. Important is the fact that the permanent habitat of the snow leopard has been reliably confirmed in the areas specified, as has the presence of breeding females in the specified territory. In the most favorable places, the occurrence of the snow leopard signs is comparable to that of the snow leopard in Southwestern Tuva, but lower than in the territory Sayano-Shushenskii Nature Reserve in 2011–2013 (Subbotin and Istomov, 2009; Karnaukhov et al., 2011; Poyarkov et al., 2011).
The coincidence of the snow leopard signs fits well-defined forms of relief known for a long time (Heptner and Sludsky, 1972; Jackson and Ahlborn, 1989). We also noted the coincidence of scrapes to notable places, where they are updated periodically, and, apparently, by different animals. In this connection an increase in marking activity (frequency of scrapes) of animals is significant, precisely along the way along linear reference points (Matyushkin and Koshkarev, 1990). The highest frequency of scrapes was noted in the central part of the southern macroslope of Tunkinskie Gol’tsy and on the Bol’shoi Sayan. In the same area we encountered the greatest number of fresh tracks on the ground. However, it should be noted that the occurrence of tracks in the summer is determined by the type of the substrate. Sand and clay patches in the passages of snow leopards are much more common in the area of Sorkhoi River than in other places of study. Traces of the snow leopard were found, first of all, on very extensive ridges with a dense network of lateral spurs. On similar ridges, animals can move from one mountain system to another or from one river basin to another.
The structure of the range of the East Sayan grouping (correlation and interposition of the main inhabited and transit habitats) has an extremely important feature: almost all the inhabited habitats identified by us are connected by transit, and if separated, then the areas of relative inadequacy are small and can easily be overcome by the animals in one diurnal transition. In our opinion, this feature to a large extent determines the possibility of survival of the snow leopard in this severe and in a number of parameters (for example, in the height of the snow cover) extreme region.
The grouping of the snow leopard inhabiting the area of the Bol’shoi Sayan, Munku-Sardyk, and Tunkinskie Gol’tsy ranges in Buryatia bears signs of an edge population. This is evidenced by the smallest the level of heterozygosity among all the samples and the smallest number of alleles. Significant genetic differences from other local samples, apparently, are due to the long-term partial isolation of this grouping from the groupings of the western regions of the species in Russia and Mongolia.
The structures of the potential habitats of the snow leopard in the western and eastern parts of the Russian range different drastically. If the western part is closely related to the key habitats on the territory of Mongolia, then the eastern part is practically devoid of such a connection. We documented that, in the western part of the range in the Tsagan-Shibetu mountain massif (Tsagaan Shivetiin Nuruu on the territory Mongolia), there is a transboundary grouping of snow leopard (Munkhtsog et al., 2015). The Mongolian part of this grouping of snow leopards is more numerous and in better condition than the Russian one. As a result of migration, snow leopards from the Mongolian territory exist and are in a relatively prosperous state of the grouping on Tsagan-Shibetu Ridge and on Chikhachev Ridge, which, in turn, are donors for more northern groupings of Shapshalskii, South Chuyskii, and eastern parts of Kuraiskii ridges. In the eastern part, the area of the snow leopard in the Russian groupings is primary for the existence of its population. This imposes additional responsibility on environmental services in Russia for the conservation of these groupings.
Essentially important for the survival of snow leopard groupings in the eastern part of the range, in our view, is the circumstance that the area of transit habitats is large and well connected to all key groupings. In this regard, the eastern part of the range appears more connected than the western part. Apparently this very important circumstance ensures the conservation of the snow leopard population in Buryatia under extremely harsh environmental conditions.
The main reason for the decline in snow leopard numbers in Buryatia, as well as in Russia as a whole, is poaching. In this case, the main threat is the widespread use of loop trapping in the hunting of musk deer. Direct pursuit of the snow leopard for its pelt is one problem, but leopards can get shot during random encounters with hunters (in Buryatia there was only one confirmed case of shooting the snow leopard when attacking livestock). Reduction in the number of ibex also affects the welfare of the snow leopard population. In connection with the foregoing, significantly strengthening the protection of the territory and persistently preventing the loop trapping of musk deer and hunting of ibexes.
Another limiting factor is the mining in the range of snow leopards (in the upper reaches of the Kitoi River and in the Kropotkinskii Range, there are gold mines, and in Sagan-Sair River, a tributary of the Kitoi River, actively developed jade mining), which leads to a decrease in predator habitats and disturbs the snow leopard during the movement of animals over these sites.
To date, in Buryatia in the habitats of the snow leopard two protected areas have been organized, Tunkinskii National Park and Shumak Nature Park. The borders of the national park completely coincide with the administrative boundaries of the Tunkinskii district of Buryatia Republic, the greater part of which is not suitable habitat for the snow leopard. Special Mode Zone protection, allocated directly to the char in habitats of the snow leopard, is not actually controlled. This is confirmed by regular encounters here with people with weapons and frequent evidence of hunting. The local population freely conducts trapping of musk deer. There are confirmed cases of the deaths of snow leopards in the musk deer traps. In addition, the territory of the national park covers only the southern macroslope of the ridge. In the northern part of the ridge, converted into the valley of Kitoi River and located on the territory of the Okinskskii district of Buryatia Republic, there is no any protected areas, although places here are suitable for the snow leopard and its presence is confirmed by our research. To optimize the conservation of the snow leopard, it is necessary to establish real control over the territory, and also include the northern macroslope of the Tunkinskii Gol’tsy in the national park with the organization of a zone with a strict mode of protection.
Another very effective measure to conserve the snow leopard would be the organization of a reserve on the Bol’shoi Sayan, west of the Munku-Sardyk mountain massif to the border with Tuva. This area is an important ecological corridor, ensuring migration of the snow leopard with the territory of Mongolia and Tuva.
REFERENCES
Baddeley, A. and Turner, R., Spatstat: an R package for analyzing spatial point patterns, J. Stat. Software, 2005, vol. 12, no. 6. http://www.jstatsoft.org/v12/i06/paper.
Bivand, R., Keitt, T., and Rowlingson, B., Rgdal: Bindings for the Geospatial Data Abstraction Library (Version R package version 0.8-10), Institute for Statistics and Mathematics, Vienna, 2013. http://CRAN.Rproject.org/package=rgdal. Accessed January 2013.
Bivand, R. and Lewin-Koh, N., Maptools: Tools for Reading and Handling Spatial Objects (Version R package version 0.8-25), Institute for Statistics and Mathematics, Vienna, Austria, 2013. http://CRAN.R-project.org/package=maptools. Accessed November 2012.
Calenge, C., The package adehabitat for the R software: a tool for the analysis of space and habitat use by animals, Ecol. Model., 2006, vol. 197, pp. 516–519. doi 10.1016/j.ecolmodel.2006.03.017
Elith, J., Graham, C.H., Anderson, R.P., Dudik, M., Ferrier, S., et al., Novel methods improve prediction of species’ distributions from occurrence data, Ecography, 2006, vol. 29, pp. 129–151.
Elith, J., Phillips, S.J., Hastie, T., Dudik, M., Chee, Y.E., and Yates, C.J., A statistical explanation of MaxEnt for ecologists, Divers. Distribut., 2011, vol. 17, no. 1, pp. 43–57.
Geptner, V.G. and Sludskii, A.A., Mlekopitayushchie Sovetskogo Soyuza (Mammals of the Soviet Union), vol. 2, Pt. 2: Khishchnye (gieny i koshki) (Carnivores (Hyenas and Cats)), Moscow: Vysshaya Shkola, 1972.
Hunter, D.O. and Jackson, R., A range-wide model of potential snow leopard habitat, in Proceedings of the 8th International Snow Leopard Symposium, Islamabad, 1997, pp. 51–56.
Jackson, R. and Ahlborn, G., A High-Altitude Wildlife Survey of the Hongu Valley with Special Emphasis on Snow Leopard. Report, 1987.
Jackson, R. and Ahlborn, G., Snow leopards (Panthera uncia) in Nepal—home range and movements, Natl. Geogr. Res., 1989, vol. 5, pp. 161–175.
Jackson, R. Rowe, D., Wangchuk, R., and Hunter, D., Izuchenie gruppirovok snezhnogo barsa s pomoshch’yu fotolovushek (The Study of Snow Leopard Groups Using Camera Traps), Krasnoyarsk: Programma razvitiya OON, 2010.
Karnaukhov, A.S., Poyarkov, A.D., Vanisova, E.A., Korablev, M.P., Aleksandrov, D.Yu., et al., The use of camera traps to study snow leopard group on the Tsagaan-Shibetu Range (South West Tuva), in Nauchnye issledovaniya v zapovednikakh i natsional’nykh parkakh Yuzhnoi Sibiri (Research in Reserves and National Parks of Southern Siberia), Novosibirsk: Sib. Otd. Ross. Akad. Nauk, 2011, pp. 78–88.
Koshkarev, E., The snow leopard in its northeastern range, Cat News, 1996, vol. 25, p. 10.
Koshkarev, E., Has the snow leopard disappeared from Eastern Sayan and Western Hovsgol?, in Proceedings of the 8th International Snow Leopard Symposium, Islamabad, 1997, pp. 96–107.
Krasnaya kniga Respubliki Buryatiya: redkie i ischezayushchie vidy zhivotnykh (The Red Data Book of the Republic of Buryatia: Rare and Endangered Species of Animals), Ulan-Ude: Informpolis, 2005.
Matyushkin, E.N. and Koshkarev, E.P., Footprints of snow leopard, Okhota Okhot. Khoz., 1990, no. 2, pp. 14–17.
Matyushkin, E.N., Footprints and tracking method in the study of large carnivores, Zool. Zh., 2000, vol. 79, no. 4, pp. 412–429.
Medvedev, D.G., Method of counting mountain ungulates in East Siberia (mountain goats and bighorn sheep), in Okhrana i ratsional’noe ispol’zovanie zhivotnykh i rastitel’nykh resursov (Conservation and Sustainable Use of Animal and Plant Resources), Irkutsk: Irkutskaya Gosudarstvennaya Sel’skokhozyaistvennaya Akademiya, 2000, pp. 89–97.
Medvedev, D.G. and Trofimov, P.S., Snow leopard in the mountains surrounding Lake Baikal, in Itogi i perspektivy razvitiya teriologii Sibiri (Results and Prospects of Development of Mammalogy in Siberia), Irkutsk: Irkutskaya Gosudarstvennaya Sel’skokhozyaistvennaya Akademiya, 2001, pp. 265–268.
Medvedev, D.G., Distribution and migration of the snow leopard in the Baikal region, in Teriofauna Rossii i sopredel’nykh territorii (Theriofauna of Russia and Adjacent Countries), Moscow: Tovar. Nauchn. Izd. KMK, 2003, p. 218.
Munkhtsog, B., Poyarkov, А.D., Korablev, M.P., Kuksin, A.N., Aleksandrov, D.Yu., et al., The documentation of transboundary snow leopard (Panthera uncia) population in Mongolia and Russia, in Ecosystems of Central Asia under Current Conditions of Socio-Economic Development: Proceedings of International Conference, Ulaanbaatar (Mongolia), September 8–11, 2015, Ulaanbaatar, 2015, vol. 1, pp. 338–341.
Nikiforov, V.N. and Shurygin, V.V., The current distribution of the snow leopard in Tuva ASSR, in Redkie vidy mlekopitayushchikh i ikh okhrana (Rare Species of Mammals and Their Conservation), Moscow: Nauka, 1977, p. 139.
Pebesma, E.J. and Bivand, R.S., Classes and methods for spatial data in R, R News, 2005, vol. 5, no. 2. http://www.citeulike.org/group/7985/article/3653190.
Phillips, S.J. and Dudík, M., Modeling of species distributions with Maxent: new extensions and a comprehensive evaluation, Ecography, 2008, vol. 31, no. 2, pp. 161–175.
Phillips, S.J., Anderson, R.P., and Schapire, R.E., Maximum entropy modeling of species geographic distributions, Ecol. Model., 2006, vol. 190, nos. 3–4, pp. 231–259.
Phillips, S.J., Dudík, M., Elith, J., Graham, C.H., Lehmann, A., et al., Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data, Ecol. Appl., 2009, vol. 19, no. 1, pp. 181–197.
Poyarkov, A.D., Lukarevskii, V.S., Subbotin, A.E., Zavatskii, B.P., Malkov, N.P., et al., Strategiya sokhraneniya snezhnogo barsa (irbisa) v Rossii (The Strategy of Conservation of the Snow Leopard (Irbis) in Russia), Moscow: Vsemirnyi Fond Dikoi Prirody, 2002.
Poyarkov, A.D., Kuksin, A.N., Karnaukhov, A.S., Korablev, M.P., Aleksandrov, D.Yu., et al., Marking activity of the snow leopard on the Tsagan-Shibetu Ridge in southwestern Tuva, in Teriofauna Rossii i sopredel’nykh territorii (Theriofauna of Russia and Adjacent Countries), Moscow: Tovar. Nauchn. Izd. KMK, 2011, p. 382.
Rozhnov, V.V., Zvychainaya, E.Yu., Kuksin, A.N., and Poyarkov, A.D., Noninvasive molecular genetic analysis in studying the ecology of the snow leopard: problems and prospects, Russ. J. Ecol., 2011, vol. 42, no. 6, pp. 439–444.
Smirnov, M.N., Sokolov, G.A., and Zyryanov, A.N., Distribution and status of the snow leopard population in the south of Siberia, Byull. Mosk. O-va Ispyt. Prir., Otd. Biol., 1991, vol. 96, no. 1, pp. 27–34.
Subbotin, A.E. and Istomov, S.V., The status of the population group of the snow leopard Uncia uncia (Felidae, Carnivora) in the Western Sayan, Dokl. Akad. Nauk, 2009, vol. 425, no. 6, pp. 846–849.
Young, N., Carter, L., and Evangelista, P., A MaxEnt Model v3.3.3e Tutorial (ArcGIS v10). Colorado State University, Fort Collins, Colorado, USA, 2011. http:// ibis.colostate.edu/WebContent/WS/ColoradoView/TutorialsDownloads/A_Maxent_Model_v7.pdf. Accessed November 2012.
ACKNOWLEDGMENTS
This work was conducted by the continuous expedition of the Russian Academy of Sciences on the study of animals by Red Books of the Russian Federation and other important animals of the fauna of Russia in the Program for studying and monitoring the snow leopard population of Siberia with the support of the Russian Geographical Society. Studies conducted in 2016 were financed by the World Wildlife Fund for Nature (WWF Russia).
The authors thank S.V. Istomov, E.P. Kashkarov, D.Yu. Bekhterev, and D.N. Kochetkov for comments and suggestions during preparation of the article.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interests. The authors declare that they have no conflict of interest.
Statement on the welfare of animals. All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Additional information
Translated by N. Smolina
Rights and permissions
About this article

Cite this article
Karnaukhov, A.S., Malykh, S.V., Korablev, M.P. et al. Current Status of the Eastern Sayan Snow Leopard (Panthera uncia) Grouping and Its Nutritive Base. Biol Bull Russ Acad Sci 45, 1106–1115 (2018). https://doi.org/10.1134/S106235901809008X
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1134/S106235901809008X