
Abstract
The European Badger (Meles meles) has been implicated in the epidemiology of bovine tuberculosis in cattle populations in the Republic of Ireland. Badger populations have been subject to a culling regime in areas with chronic histories of bTB cattle herd breakdowns. Removal data from 2004 to 2010 were used to model the impact of culling on populations in areas under capture. Additionally, changes in field signs of badger activity were used as an index of abundance to support, or otherwise, the outcomes of the removal models. Significant reductions in standardised badger captures over time were found across three large study areas (total area, 1,355 km2). Assuming that all inactive setts were vacant, an overall linear trend model suggested that badger captures had decreased by 78 % for setts with 6 years of repeated capturing operations. Given the uncertainty associated with the relationship between sett activity and badger presence, we repeated the linear modelling using two ‘what if’ scenarios. Assuming that individual badgers were missed on 10 % or 20 % of occasions at inactive setts, the estimated decline over 6 years is lowered to 71 % or 64 %, respectively. The decline profile consisted of a steep initial decrease in captures within the first 2 years, followed by a more gradual decrease thereafter. The number of active openings at setts (burrows) declined significantly in all three areas; but the magnitude of this decline varied significantly amongst study areas (41–82 %). There was a significant increase in the probability of setts becoming dormant with time. The removal programme was more intense (mean, 0.45 badgers culled km−2 year−1) than previous experimental badger removals in Ireland but some captures may be attributed to immigrant badgers as no attempt was made to limit inward dispersal from areas not under management. Results from this study suggest that significant reductions in badger density occurred in the areas where management had taken place. Since other non-culled badger populations in Northern Ireland and Britain exhibited stable population trends, we attribute the reduction in relative abundance to the culling regime. Further studies of the dynamics of this reduction are required to quantify how it is counteracted by immigration from populations outside of culled areas.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The European badger (Meles meles) is the main wildlife reservoir of bovine tuberculosis (bTB, Mycobacterium bovis) in the Republic of Ireland and the UK (More and Good 2006; Gortázar et al. 2011). Since 2004, a programme has been implemented within the Republic of Ireland to reduce the density of badgers in areas with chronic problems of bTB in cattle herds (O’Keeffe 2006; Sheridan 2011). This has involved the capturing (with stopped restraints) and removal of badgers in areas up to 2 km from breakdown farms. The assumption underlying such a strategy is that a reduction in density of a disease host reduces the contact and transmission rates both within that host species and between different host species (Woodroffe et al. 2008). The scientific basis for this programme originated from two prior, experimental removals (the Four Area Project (FAP) and the East Offaly Project (EOP)), where extensive culling over large study areas (188–528 km2) was associated with significant decreases in bTB herd breakdowns (O’Mairtin et al. 1998; Griffin et al. 2005).
Despite the national culling programme, the badger is a protected species in the Republic of Ireland under the Irish Wildlife Act and is listed under appendix III of the Bern convention. Badgers also play an important role in temperate ecosystems as they act as ecosystem engineers, seed dispersers and predators (Byrne et al. 2012a; Roper 2010). Trends in badger populations, in areas under capture (AUC), should be assessed from a conservation perspective, in order to evaluate any effects of the removal regime on the badger’s conservation status.
This study is the first attempt at formally assessing the impact of the current culling programme on local badger populations. We examine trends in badger relative abundance over time in capture areas in three counties that were subject to extensive culling. Changes in badger relative abundance were inferred using two indices: badger captures per standardised capture event and changes in signs of activity at setts. Multivariable statistical models were employed to estimate the relative reduction in badger captures over time. This analysis was complemented with a similar investigation of the changes in the frequency of signs of badger activity at setts (badger burrows) and the likelihood of setts becoming dormant over time. A number of studies have found positive relationships between badger numbers and field signs of activity (e.g. Tuyttens et al. 1999; Tuyttens et al. 2001; Wilson et al. 2003; Sadlier and Montgomery 2004; Woodroffe et al. 2008; Szmaragd et al. 2010; Byrne et al. 2012b). We recognise that the predictability of field signs is imperfect, and the strength of the relationships between field signs and badger numbers can be impacted by factors such as season and habitat type (Wilson et al. 2003). Furthermore, the relationship between abundance and field signs of activity may not be linear (Woodroffe et al. 2008), for example a reduction in density may result in changes in sett visiting behaviour by any remaining, or neighbouring badgers, resulting in field signs under representing local abundance. Despite these limitations, field signs of activity have been used previously to infer effects of culling on wildlife relative abundance (e.g. Baker and Harris 2006; Woodroffe et al. 2008). These indirect methods of estimating wildlife numbers can be implemented at large spatial scales and at low cost (in comparison to direct methods, e.g. mark-recapture), and can provide a consistent measure to infer broad abundance trends at these scales (Bonesi and Macdonald 2004; Sadlier et al. 2004; Woodroffe et al. 2008).
Methods
Study areas
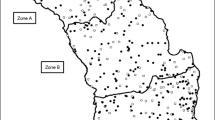
Study areas were chosen within three inland counties within the Republic of Ireland—counties Monaghan, Longford and South Tipperary (see Fig. 1). Large areas of these counties (mean = 31 %) had been under a badger culling regime, which began in 2004. The counties were matched in terms of field staff experience and efficiency. Most setts were located in areas where the dominant land cover type was agricultural grassland, interspersed with woodland or scrub (Fig. 1).

Maps of the study areas of Co. South Tipperary (a), Co. Monaghan (b) and Co. Longford (c). The extent of the badger AUC is delineated by the thick black lines. Preferred badger habitats (mainly dry grasslands, mature woodland and scrub) are represented as white areas using an indicative county habitat map (Fealy et al. 2009). Grey areas are made up of poor or non-badger habitats including open water, wetlands, fens, bogs and rocky complexes. a Much of South-Tipperary is dominated with dry grasslands. Unsurveyed lands in the south and west correspond to uplands with bog, heath and rocky complexes; areas around the northern border are predominantly cutover raised bog lands. b Monaghan is dominated by low, elongated, hills of glacial till (drumlins). Unsurveyed areas in the north-west border are made up of upland blanket bog; further south are areas of reclaimed raised bog. Unsurveyed mid-east areas have lake lands and reclaimed raised bogs. c Co. Longford has the most non-badger habitat area. Large areas of the county in the south west are unsurveyed, corresponding with open water and cutover raised bogs
Sett surveying and badger capturing protocol
Badgers were captured as part of a medium-term national bovine tuberculosis (bTB) control strategy. Detailed descriptions of this programme have been given by O’Keeffe (2006) and Sheridan (2011). Surveys for evidence of badgers on farm land with a bTB breakdown (i.e. new bTB occurrence), and adjacent land (up to 2 km beyond the farm boundary), are instigated as a result of a veterinary epidemiological investigation after a herd breakdown. Presently ∼22 % of all bTB breakdowns nationally lead to badger surveying. Field teams (n = 11–16 people across the three counties) use multiple strategies to locate badger setts within the landscape. Local knowledge (through farmers, local huntsmen, game societies etc.) of sett locations is recorded and the sites checked to validate the record (to ensure that it is a badger sett and not a fox den, for example). Maps and aerial photographs are used to increase the likelihood of finding setts by targeting areas of woodland, scrub, riparian vegetation, ringforts (archaeological remains where badger setts are often found) and well-developed hedgerow networks. Field signs (paths, rooting and latrines) are also used to help locate badger setts.
The capture of badgers involves a standardised block of 11 nights of capturing effort at a sett. These standard blocks are known as capture events. Cable stopped restraints were used to capture badgers (see Anon. 1996 for details). These restraints have been utilised extensively in badger studies in the Republic of Ireland. The majority of badgers captured using this technique have no or minimal injuries (e.g. 98.8 % exhibit either no signs of muscle bruising or slight bruising, only 1.2 % exhibited areas of haemorrhage and tearing of the underlying muscle; Murphy et al. 2009). The restraints were located predominantly at the entrance to active sett openings and along badger paths to maximise the probability of capture (see Sleeman et al. 2009 for details). The number of restraints placed at, or near, each sett was determined by the level of badger activity detected at that time by experienced trained field staff. The mean number of restraints laid per sett was 10.6 (SD 5.6; range = 1–50)). Restraints were checked daily before 1200 hours. If field staff considered that badgers remained (i.e. evaded capture) after a removal event, a new capture event would be initiated immediately. Otherwise, setts were revisited at a minimum intensity of once per year to assess if the local setts showed evidence of activity. If badger activity was apparent, the sett(s) would be re-captured (i.e. a new event would be triggered), using the same protocol as before.
All badger removals were conducted under licence from the Department of Environment, Republic of Ireland. Licences were granted for each county on yearly time periods for the duration of the study (2004–2010; Licence numbers: Longford—25N/2004–25N/2010; Monaghan—29R/2004–29R/2010; South Tipperary—34V/2004–34V/2010). Restraints used conformed to national legislation for humane trapping (Wildlife Act, 1976, Regulations 2003 (S.l. 620 of 2003)). All licensing, capturing and culling adhered to the Irish Wildlife Acts (1976 to 2010–section 23(6)(A)).
Dataset structure
Closely grouped setts were trapped simultaneously to improve efficiency. These groups typically contained 5–10 setts and were called ‘capture blocks’; and each capture block was given an identifier within the dataset. Setts within a capture block were surveyed during each event, though attempts to capture badgers were only made where there was some evidence of badger activity at a sett. Setts that disappeared (e.g. had been abandoned) during the study period were maintained in the dataset, but coded ‘0’ for activity ensuring that data for every sett within a capture block were present for each event. This procedure was implemented in Stata® 11 and affected ∼1 % of the total dataset.
Within the dataset, setts with no signs of badger activity were considered capture events with an outcome of ‘zero capture’. Although not formally assessed, previous experience indicated that the absence of signs of activity has a high specificity for predicting the absence of badgers; the presence of activity has only a moderate sensitivity for predicting the presence of badgers in a specific sett. It should be noted that fieldworkers employ a precautionary principle during capturing attempts, whereby restraints are laid at setts where there is minimal evidence of badger presence (J. O’Keeffe, personal communication). Conversely, restraints are not laid at setts where there is a clear indication that badgers have not been using the sett recently. For example, a typical sign of lack of use would be grass growing within the openings to a sett. In order to meet our population-based objectives, and to reflect the changing activity pattern of all setts over time, we included these uncaptured setts as ‘zero outcome’ data in our models. However, to ensure our analysis is robust in relation to this assumption, two scenarios were implemented whereby we allowed 10 % and 20 % of events at inactive setts to yield a badger (see the ‘The effect of assuming inactive setts were vacant—what-if scenarios’ section below).
Measuring sett activity
Sett activity was used as an additional measure of badger relative abundance. Field signs used to assess activity included: evidence of fresh digging, evidence of movement into or out of an opening, the presence of fresh tracks and the presence of bedding material. The number of openings (entrances) within a single sett that showed any of these signs of activity was recorded. Setts with no field signs of opening activity were recorded as dormant.
Descriptive analysis
In order to estimate sett densities and the intensity of the removal programme, an AUC representing the geographic extent of the removal regime had to be estimated. As initial surveying of badger setts was limited to the area in and around the breakdown farm, the locations of main setts beyond these surveyed areas were unknown. This precludes the use of tessellations in order to estimate the configuration of probable badger territories (e.g. Hammond and McGrath 1998; Halls et al. 2001). As an alternative, the half-mean nearest neighbour distance between setts, from areas where all sett locations were known, was used as a proxy for typical sett spacing in Irish agricultural landscapes. For the present study, we used the distance between setts derived from the FAP and the EOP (Eves 1999; Griffin et al. 2005); thus the mean nearest-neighbour distance for main setts was 916 m, whereas the corresponding mean distance for all setts was 289 m (G. McGrath, personal communication). We conservatively estimated the AUC by applying a buffer of 500 m around all setts, where overlapping circles were dissolved to coalesce into the larger surface of the AUC. This GIS approach has been utilised extensively during bTB programme monitoring and reporting in the Republic of Ireland (O’Keeffe 2006; Healy 2010; Sheridan 2011; G. McGrath, personal communication). Note that this method would tend to marginally underestimate sett densities where known setts are spatially dispersed.
Modelling approach
Count data models were constructed within a Generalised Estimating Equation (GEE) framework, of the number of badgers caught over time, to infer the relative reductions in badger abundance. GEE models are extensions of the Generalised Linear Model method (GLM) to correlated datasets (McCullagh and Nelder 1989), such that valid standard error estimates for model parameters can be drawn (Liang and Zeger 1986). The repeated captures from the same cohort of setts can be thought of as a longitudinal dataset whereby each observation (capture attempt) is not independent. GEE incorporates this non-independence through the inclusion of a correlation matrix amongst the captures from the individual setts. GEE is considered the best approach when the outcome of interest is a population average estimate (Dohoo et al. 2010).
Initially Poisson models were fitted, but since the variance of the response variable was greater than the mean, negative binomial model distributions were subsequently fitted to the datasets. A likelihood-ratio chi-square test was used to formally evaluate if the negative binomial model was a better fit to the data. This tests whether or not the dispersion parameter α is equal to zero (Hilbe 2011).
The default dispersion parameter value (α) for a GEE model with a negative binomial distribution is 1. This effectively ignores the extra variance in the data, so α was estimated from a maximum likelihood GLM model (Hardin and Hilbe 2003). The link function used in the analysis was the log link, and an exchangeable correlation matrix structure with robust standard errors was employed. Robust standard errors are generated empirically from the data, and give valid standard errors even if the assumed correlation structure is incorrect (Dohoo et al. 2010). GEE models are not fitted using maximum likelihood, thus Akaike’s Information Criterion (AIC) for model selection could not be utilised. Quasi-likelihood Information Criterion (QIC) values for the GEE models were used instead to compare competing models (Pan 2001). Both the QIC and QICμ test statistics were utilised during model selection (Pan 2001; Hardin and Hilbe 2003). QICμ approximates QIC when the GEE model is correctly specified. However, QICμ adds a penalty to the quasi-likelihood for additional parameters included, thus, parsimonious models are selected for. The model with the lowest QIC values was considered the model with the best goodness-of-fit to the data; models with ΔQICμ ≤ 2 were considered equivalent, with the preferred model having the fewest parameters. Data manipulation and statistical analyses were completed in Stata® version 11.
Assessing trends
The response variable used was μ sett which is the (population averaged) mean expected number of badgers caught per sett. Setts were recruited to the study at different time points (dates) and interval times between sequential captures varied in accordance with sett activity. Thus, time since recruitment (TIME; scaled to years) into the study was used as the temporal predictor in all analytic models. A dichotomous variable MAIN was included to control for sett type (main setts are larger and more complex—see Sleeman et al. (2009) for details), while the inclusion of MONTH variables controlled for the effects of seasonality (12 levels). The effect of each study area was controlled with the inclusion of an AREA variable. The dependency of the decline in captures on each study site was evaluated with the inclusion of an AREA × TIME interaction term. The clustering variable (i.e. where the repeated measure took place) was the sett identifier.
Linearity between continuous predictors and outcome was tested using the Lowess smoothing regression function within Stata® 11. Where non-linear relationships were found, a piecewise (spline) regression approach was employed (see below). Correlation between predictors was assessed using the Pearson correlation coefficient. Confounding was assessed by inspecting the change in magnitude (or sign direction) of the predictor’s coefficient when an additional predictor was added to the model (Dohoo et al. 2010). The overall significance of categorical variables was tested using Wald tests.
Splines were created within Stata® and a piecewise regression was run in order to model the non-linear relationship between badger captures and time since recruitment. It was necessary to investigate where change points (also called knots or cutpoints (Dohoo et al. 2010)) occurred in order to run the piecewise regression. To achieve this, the relationship between the number of badgers captured and time since recruitment, with time categorised into yearly time points (0, 1, 2, 3, etc.), was modelled. A hierarchical model structure was then employed to assess where significant changes in the relationship occurred (Dohoo et al. 2010). This model tests for the difference between a coefficient estimate from one level and its preceding coefficient estimate (i.e. 1 vs. 0; 2 vs. 1; 3 vs. 2 etc.).
During model construction, the existence of significant interactions between the TIME splines and site were tested (i.e. whether the rate of decline of each (spline) period differed significantly amongst the three sites). An additional ‘average’ trend model was also applied to the data for comparative purposes, where linearity of decline was assumed.
The effect of assuming inactive setts were vacant—what-if scenarios
To investigate the effect of the assumption that inactive setts contained no badgers, two hypothetical scenarios were devised. We allowed single badgers to be caught at (a) 10 % and (b) 20 % of events at inactive setts. The latter would be considered a worst case scenario. We used a pseudo-random number generator to sample 10 % or 20 % of setts during capture events where no restraints were laid and ‘0’ badgers recorded. To ensure that the parameter estimates were not biased by the sample, we iteratively repeated the process ten times. Each iteration produced a new capture dataset (ten datasets, by two scenarios), and the linear trend model was run on each dataset. The maximum and minimum parameter estimates across samples are reported. The decline was calculated from the mean of the parameter estimates; 95 % CI are the maximum and minimum confidence intervals estimated across each scenario.
Analysis of sett activity
Sett activity was analysed in two ways: by the number of openings that were active per sett and by the proportion of dormant setts surveyed. The number of active openings in setts was modelled in a negative binomial regression GEE model (similar in structure to the capture data). The probability of a sett being dormant was modelled using logistic regression within a GEE framework. The model was within the binomial family, with the logit link function and exchangeable correlation structure. The logistic model was evaluated using the goodness-of-fit test for binomial GEE models (Hardin and Hilbe 2003) developed by Horton et al. (1999).
Results
Descriptive analysis
There were 2,516 known badger setts surveyed during the study from 1,355 km2 of agricultural land, giving a mean sett density of 1.9 setts km−2 (range, 1.76–2.04 setts km−2; Table 1). An average of 31 % (range, 25–37 %) of the land area of each county was included in the study area. Approximately a quarter of all setts were considered main setts (23.9 %; range, 21.73–27.51 %; Table 1). In total 57,000 restraints were laid, resulting in 627,000 trap nights of effort. The number of setts captured per year increased during 2004–2005 as more setts were recruited into the cull regime, before stabilising from 2006 onwards (mean, 10,700; SE, 360). A total of 3,861 badgers were removed from the study areas over the study period, giving an overall mean badger removal rate of 2.8 badgers km−2 (range, 2.43–3.06). The average removal intensity was 0.45 badgers km−2 year−1 (range, 0.36–0.50) in the years 2005–2010 (Table 2). Half of all setts did not yield a badger, and of these the majority (88.6 %) were non-main outlier setts (Table 2).
Model of badger captures
During initial GLM model construction all independent variables were significant predictors and so all were offered to the final GEE model. All main effects of all variables presented to the multivariable GEE model were retained in the final model (i.e. p < 0.05; Table 3), with the exception of the interaction terms (TIME × AREA for each spline) which were non-significant (Wald test, p > 0.05). This indicated that the magnitude of the decline, over each spline time period, was not significantly different amongst counties.
The cut-point model indicated that there was a significant change in slope between years 0–1 and 1–2; thus the spline knots were located at these points creating a model with three periods of decline during which the relationship was assumed to be linear (Fig. 2). The piecewise GEE model indicated that there were significant declines in captures during all three time periods (i.e. slope < 0; p < 0.02). The greatest decline in captures was during the first year post recruitment, with an annual rate of decline of 43 % (95 % CI, 36–50 %). During the second year the rate of decline was reduced to an 18 % annual decline (4–30 %), and thereafter the estimated annual rate of decline was 10 % (2–17 %). The model fitted the data well during the first 5 years; however, there was greater variability in capture rates thereafter corresponding to a smaller sample size (Fig. 2; Table 4).

The relationship between badger capture frequency per event and years since recruitment. The solid line represents the predicted capture from a spline model with two knots (cut-points). Cut points are delineated by dashed vertical lines. Circles represent the mean 3-monthly captures, with circle size weighted by the number of badger setts captured during the period. The coefficient of decline for each spline progressively gets smaller over time (β = −0.57; β = −0.20; β = −0.10, respectively)
To establish the average decline in captures, an overall trend model was fitted to the data. The average linear trend model indicated that there was a decline in captures of 21 % (95 % CI, 19–25 %; p < 0.01) per annum. This model indicates that captures from setts over 6 years would decline overall by 78 % (95 % CI, 72–82 %). However, considering the non-linearity between the predictor and outcome variable this estimate needs to be interpreted with caution. The linear model tended to underestimate the initial steep decline and overestimate the percentage decline after 4 years post recruitment.
What-if scenarios
The parameter estimates (β) for TIME across 10 random samples varied from −0.202 to −0.212 for scenario (a), and −0.166 to −0.177 for scenario (b) and were highly significant in all models (p < 0.001). This resulted in the mean estimated linear trend for setts with 6 years of capture being reduced to 71 % (95 % CI, 65–76 %) and 64 % (95 % CI, 59–69 %) under scenario (a) and (b), respectively.
Activity
All predictors offered to the final activity model were retained, including an interaction term AREA × TIME, which indicates a significant difference between the reduction in activity over time amongst the study areas (Table 5). The negative binomial regression model indicated an overall significant reduction in the number of active openings per sett over the 6 years since recruitment (main effect of TIME, p < 0.001; Table 5). There was a significant difference in the number of active openings between main and non-main setts (Table 5). For main and non-main setts, there was a decline in the mean number of active openings of 68 % and 87 %, respectively (Table 6). There was a slight increase in the mean number of active openings between the first survey and the first year of capturing for non-main setts. The greatest estimated decline in activity at sett openings over 6 years was 82 % (annual rate of decline, 25 %; 95 % CI, 20–29 %) in Monaghan, with an intermediate reduction in Longford of 58 % (annual rate, 13 %; 95 % CI, 10–16 %) and the lowest reduction in South Tipperary of 41 % (annual rate, 8 %; 95 % CI, 5–11 %).
The proportion of setts deemed completely dormant on the basis of no field signs at openings increased from 29 % to 64 % for main setts over the study period (Fig. 3). Similarly, there was a general trend of an increasing proportion of non-main setts becoming dormant, with a change from 46 % to 90 % (Fig. 3). For both sett types, there was a slight decrease in the proportion of setts deemed dormant during the first year post recruitment.

The proportion of main (solid black line) and non-main (dashed) setts found during surveys to be dormant (no signs of activity) during each yearly period post-recruitment
The binomial logit GEE model was significantly better than a null model (p < 0.001), and there was no evidence of a lack of fit to the data (χ 2(2 df) = 3.70; p = 0.157). All variables were significant predictors of sett dormancy, as well as the interaction term for YEAR × AREA indicating that the rates of dormancy varied significantly amongst sites over time (Table 7). The probability of a sett becoming dormant significantly increased over time for all three areas (Table 7). Monaghan had a greater probability of an increase in sett dormancy over time then either Longford or South Tipperary. There was no significant difference of the effect of TIME on the probability of sett dormancy between Longford and Tipperary (post hoc Wald test: χ 2(1 df) = 1.58; p = 0.2). Main setts had a lower probability of becoming dormant over time than non-main setts (β = −0.98; p < 0.001).
Discussion
Our analysis shows significant reductions in the number of active openings at setts (40–82 % decline), decreases in the number of active setts (53–59 % decline), and increases in the probability of a sett becoming dormant over time. The reductions in signs of badger activity at sett openings varied significantly across counties. Monaghan had far greater reductions in badger activity and setts were significantly more likely to become dormant over time than in either Longford or South Tipperary.
Part of county Monaghan (368 km2; 28 % of the county) had been involved in the Four Area Project (1998–2002; Griffin et al. 2005), which may account for the reduced activity recorded at setts within these areas during the present study. A model was constructed to test if there was a difference in activity levels between setts found within the removal area and elsewhere within Monaghan (GEE-NB model). There were significantly fewer active openings recorded in setts found within the removal areas than elsewhere (β = −1.06; p < 0.001). We re-modelled the activity data across the three counties without the removal area setts from Monaghan. The interaction terms remained significant, and the parameter estimates did not deviate in a substantial way from the full model (reduced model β 1 = 0.13, β 2 = 0.19, p < 0.001; full model β 1 = 0.14, β 2 = 0.20, p < 0.001). This indicates that the inclusion of setts from the removal area did not have an overall impact upon the estimates drawn from the full three county model. Therefore other factors affected the differences in reduced signs of activity over time amongst counties. It must be kept in mind that the relationship between badger numbers and field signs may not be linear, and may be affected by season, habitat and methodology (Wilson et al. 2003; Sadlier and Montgomery 2004). Fieldworkers in all three counties have been trained to implement the same methodology and were matched in terms of field experience, and it would be fair to assume that seasonal effects are the same for all counties. Despite the AUCs in the counties being similar, there are large scale differences in the landscape composition amongst the three counties; for example South Tipperary has the greatest amount of deep, well-drained soils (49 %) in comparison with Longford (33 %) and Monaghan (25 %) (Fealy et al. 2009). This results in (a) a greater intensity of farming (more improved pasture) and (b) good soil conditions for badgers to dig setts, both of which features have been associated with elevated numbers of badgers in Ireland (e.g. Hammond et al. 2001). If more of Tipperary South has better conditions for badgers, we might expect greater immigration pressure into the removal areas, thus affecting the rate of decline in captures over time. These speculations need to be investigated further.
The present study showed significant declines in badger captures as culling continued, averaging 78 % decline for setts captured over a 6-year period. Recent culling operations in the south-west of England (Randomised badger Culling Trial or RBCT) achieved significant reductions in the density (setts km−2) of active openings (69 %) and active setts (59 %) through proactive removal of badgers (Woodroffe et al. 2008). Proactive culling implemented during these operations involved capturing badgers in cage traps across ten areas of 100 km2 each. A second strategy, during the same study, involved localized reactive culling, where badgers were only removed on land used by a herd that had experienced a bTB breakdown. As expected, this latter removal strategy resulted in lower reductions in sett activity per unit area (e.g. 17 % reduction in active sett density; 26 % reduction in active openings density). A reduction in the numbers of badger captures across successive culls in the RBCT was evident but the magnitude of this trend was not formally evaluated (Woodroffe et al. 2008, Fig 1b). As with the present study, these activity indices and badger capture profiles were used to indicate the success of that culling regime in reducing the relative abundance of badgers. While both studies found evidence of reductions in badger abundance, there are a number of reasons why it would be inappropriate to compare directly the magnitude of these reductions. Badgers were captured using different methods (stopped restraints vs. cages), which may have different efficiencies and biases (O’Connor et al. 2012); however, the relative efficiency or bias in terms of badger capture is currently unknown (but see Muñoz-Igualada et al. 2008 for a study with red fox). Badger densities are greater in south-west England than Ireland generally (Byrne et al. 2012a, b), which probably has an impact on the way badger populations respond to culling. Most fundamentally, the way the areas surveyed were delineated differed between the two studies (the RBCT had explicitly defined the boundaries of their study area, whereas the AUC was estimated in the present study).
As part of the policy of the removal programme, to maximise efficiencies, no attempt was made to capture badgers at setts without signs of recent badger activity (mostly at non-main setts; Table 2). This ensures that effort is focused upon setts with the highest likelihood of capturing badgers. However, it also means that we assume there is a high specificity in the field staffs ability to recognise inactive setts. While it may be difficult to estimate badger numbers from field signs with accuracy (e.g. Wilson et al. 2003), it is a far simpler task for trained experienced field staff to judge presence/absence, especially when the threshold for recording an absence is set high. Despite this, it is likely on rare occasions that badger capturing was not attempted in situations when badgers were actually present. If this is the case, the model would be biased towards giving overestimates in the rate of the decline (estimated β). As badger surveying and capturing is frequently repeated, and as the culling regime continues in these areas, resident badgers that evade capture during initial events have very low likelihoods of survival due to subsequent follow-up culls. To assess the sensitivity of the models to the zero-capture assumption, models were developed under two scenarios where individual badgers were missed on either 10 % or 20 % of occasions. For both scenarios, there remained significant estimated declines of captures over time of a large magnitude (64 % or 71 % over 6 years; p < 0.001). These scenario outcomes, and the broad consistency of our findings across indices, suggest that the inferences made from our models are robust.
The mean badger removal intensity during our study was 0.45 badgers km−2 year−1. This is higher than the mean rate of 0.33 badgers km−2 year−1 (range 0.21–0.48) achieved during the Four Area Project (FAP; 1997–2002; data from Corner et al. 2008) or 0.34 badgers km 2 year−1 for the East Offaly Project (EOP; 1989–1995; Kelly et al. 2008). Kelly and others (2008) reanalysed data from the EOP area with additional removal data up to 2004. Across all years (1989–2004), the average removal intensity was 0.23 badgers km−2 year−1. During these studies, barriers to inward dispersal were implemented. Therefore the higher capture rates recorded during the present study may reflect the capturing of immigrant badgers. Removal intensities were far higher during the RBCT in Britain, with average rates of 1.83 badgers km−2 year−1 (Bourne et al. 2008). This was despite the lower presumed efficiency (due to the use of cage traps; O’Connor et al. 2012) and lower frequency of trapping during the RBCT study (Bourne et al. 2008) compared with our study. This suggests that there was a higher badger population density in the RBCT study areas than in the areas of the present study, prior to trapping and removal (Bourne et al. 2008; Wilson et al. 2011).
We are confident that the declines demonstrated in our analysis result from the badger culling regime and not from other extraneous factors. While there were no explicit controls within the present study (i.e. unculled areas where trends in the population were estimated), a number of lines of evidence suggest that the abundance of unculled badger populations within the British Isles is stable. In Northern Ireland, where badger populations are not culled for bTB management, long-term monitoring of setts has revealed a stable badger population (Feore 1994; Sadlier and Montgomery 2004; Reid et al. 2008, 2011). Feore (1994) completed the first assessment of badger abundance, surveying 129 1 km2 sites for setts and signs of badger activity. No significant changes in the densities of setts were demonstrated amongst a subsample of these sites (20 of 129 1 km2 sites) between 1990/1993 and 1997/1998 (Sadlier and Montgomery 2004). There were significant increases in the proportion of setts deemed active for some non-main sett types, but not for main setts. A repeat survey in 2007/2008 of all sites also found no statistically significant change in the estimated population size in Northern Ireland (Reid et al. 2008, 2011).
There have been two long-term studies of undisturbed high-density badger populations in Britain where population size has been monitored. In Wytham Woods, the trend in the badger population abundance has remained stable during the period of our study (2004–2010 inclusive; Dr. C. Newman, personal communication). Similarly in Woodchester Park, the number of badgers present has remained relatively stable from 2004 through to the most recent population estimate in 2007 (Defra 2011).
Across much of continental Europe increases in badger abundance have been recorded (Holmala and Kauhala 2006; Kranz et al. 2008). A recent analysis of the national German badger populations over a period contemporaneous with the present study (2003–2007) found that badger numbers and reproductive output stayed stable despite hunting pressure (Keuling et al. 2010). An average of 52,817 badgers in Germany have been killed by hunters annually since 2003 (total to 2011, 422,535), equating to a removal intensity of ∼0.14 badgers km−2 year−1 (Keuling et al. 2010; Deutscher 2012). Similarly, in Finland where there is an increasing trend in the badger population ∼10,000 badgers per annum are hunted, which equates to 0.05 badgers km−2 year−1 (assuming that badgers only inhabit 60 % of the country (Kauhala 1995; Kauhala and Auttila 2010; Kauhala and Holmala 2011)). In the context of these positive or stable regional and national trends in badger population abundances, the strongly negative trends described in this paper indicate that the culling regime is having a significant impact on badger abundance in the study areas.
Implications of reduced badger density
From a conservation perspective, our analysis suggests that badger populations have been greatly reduced over large areas of the Irish countryside (31 % of the area of the counties in the present study). Despite this, badgers are continually caught at setts even after recurrent capture attempts over multiple years. This indicates that a likely source-sink dynamic is in place. The medium-term programme in Ireland has a conservation measure built in, whereby no more than 30 % of the agricultural land area nationally can be under capture (Sheridan 2011). As a future conservation measure, it may be important to monitor badger populations in order to prevent extinction of populations at a regional scale and to ensure the maintenance of a viable national population. The most recent estimate of the national badger population size for the Republic of Ireland was 84,000 (95 % CI, 72,000 to 95,000; Sleeman et al. 2009), so the possibility of a national eradication is unlikely. However, this population estimate was made prior to the current removal programme, so this did not incorporate the impact of large-scale badger removals. Future population modelling should incorporate estimates of this removal programme’s effect.
References
Anon (1996) Badger manual. Department of Agriculture, Food and Forestry, Dublin
Baker PJ, Harris S (2006) Does culling reduce fox (Vulpes vulpes) density in commercial forests in Wales, UK? Euro J Wildl Res 52:99–108
Bonesi L, Macdonald DW (2004) Evaluation of sign surveys as a way to estimate the relative abundance of American mink (Mustela vison). J Zool 262:65–72
Bourne J, Donnelly C, Cox D, Gettinby G, McInerney J et al (2008) Badgers and cattle TB: the final report of the independent scientific group on Cattle TB. The Stationery Office, UK
Byrne AW, Sleeman DP, O’Keeffe J, Davenport J (2012a) The ecology of the European badger (Meles meles) in Ireland—a review. Biol Environ—Proc R Irish Acad 112B:105–132
Byrne AW, O’Keeffe J, Sleeman DP, Davenport J, Martin SW (2012b) Factors affecting European badger (Meles meles) capture numbers in one county in Ireland. Prev Vet Med (in press)
Corner LAL, Clegg TA, More SJ, Williams DH, O’Boyle I et al (2008) The effect of varying levels of population control on the prevalence of tuberculosis in badgers in Ireland. Res Vet Sci 85:238–249
Defra (2011) The long term intensive ecological and epidemiological investigation of a badger population naturally infected with M. Bovis—SE3032. randd.defra.gov.uk/Document.aspx?Document = 9871_textfor2011SE3032finalreport.doc. Accessed 4th June 2012
Deutscher Jagdschutz-Verband (2012) Deutscher Jagdschutzverband, Handbuch 2012. http://medienjagd.test.newsroom.de/201011_strecke_dachse2.pdf. Accessed 10th April 2012
Dohoo IR, Martin SW, Stryhn H (2010) Veterinary epidemiologic research, 2nd edn. VER Inc, Canada
Eves J (1999) Impact of badger removal on bovine tuberculosis in east County Offaly. Irish Vet J 52:199–204
Fealy RM, Green S, Loftus M, Meehan R, Radford T et al (2009) Teagasc indicative habitat maps by county. In: Fealy RM, Green S (eds) Teagasc EPA soil and subsoils mapping project-final report. Teagasc, Dublin
Feore SM (1994) The distribution and abundance of the badger Meles meles L. Queens University Belfast, Northern Ireland
Gortázar C, Delahay RJ, McDonald RA, Boadella M, Wilson GJ, Gavier-Widen D, Acevedo P (2011) The status of tuberculosis in European wild mammals. Mamm Rev. doi:10.1111/j.1365-2907.2011.00191.x
Griffin JM, Williams DH, Kelly GE, Clegg TA, O’Boyle I et al (2005) The impact of badger removal on the control of tuberculosis in cattle herds in Ireland. Prev Vet Med 67:237–266
Halls PJ, Bulling M, White PCL, Garland L, Harris S (2001) Dirichlet neighbours: revisiting Dirichlet tessellation for neighbourhood analysis. Comp Environ Urb Syst 25:105–117
Hammond RF, McGrath G (1998) The use of geographical information system (GIS) derived tessellations to relate badger territory to distribution patterns of soils and land use environmental habitat variables within the East Offaly badger research area. ERAD Tuberculosis Investigation Unit, University College Dublin, Selected Papers 1997. pp. 14–25
Hammond RF, McGrath G, Martin SW (2001) Irish soil and land-use classifications as predictors of numbers of badgers and badger setts. Prev Vet Med 51:137–148
Hardin JW, Hilbe JM (2003) Generalized estimating equations. Chapman & Hall/CRC, London
Healy R (2010) TB Programme 2011: Annex 1. Report to the European Commission. http://www.agriculture.gov.ie/media/migration/animalhealthwelfare/diseasecontrols/tuberculosistbandbrucellosis/diseaseeradicationpolicy/2011TBProgramme130111.pdf . Accessed 10th April 2012
Hilbe JM (2011) Negative binomial regression. Cambridge University Press, UK
Holmala K, Kauhala K (2006) Ecology of wildlife rabies in Europe. Mamm Rev 36:17–36
Horton NJ, Bebchuk JD, Jones CL, Lipsitz SR, Catalano PJ et al (1999) Goodness-of-fit for GEE: an example with mental health service utilization. Stat Med 18:213–222
Kauhala K (1995) Changes in distribution of the European badger Meles meles in Finland during the rapid colonization of the raccoon dog. Ann Zool Fenn 32:183–191
Kauhala K, Auttila M (2010) Habitat preferences of the native badger and the invasive raccoon dog in southern Finland. Act Theriol 55:231–240
Kauhala K, Holmala K (2011) Landscape features, home-range size and density of northern badgers (Meles meles). Ann Zool Fenn 48:221–232
Kelly GE, Condon J, More SJ, Dolan L, Higgins I et al (2008) A long-term observational study of the impact of badger removal on herd restrictions due to bovine TB in the Irish midlands during 1989–2004. Epidemiol Inf 136:1362–1373
Keuling O, Greiser G, Grauer A, Strauss E, Bartel-Steinbach M et al (2010) The German wildlife information system (WILD): population densities and den use of red foxes (Vulpes vulpes) and badgers (Meles meles) during 2003–2007 in Germany. Euro J Wildl Res 57:95–105
Kranz A, Tikhonov A, Conroy J, Cavallini P, Herrero J, Stubbe M, Maran T, Fernandes M, Abramov A, Wozencraft C (2008) Meles meles. In: IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2. www.iucnredlist.org. Downloaded on 09 January 2012
Liang KY, Zeger SL (1986) Longitudinal data analysis using generalized linear models. Biometrica 73:13–22
McCullagh P, Nelder JA (1989) Generalized linear models. Chapman & Hall/CRC, London
More SJ, Good M (2006) The tuberculosis eradication programme in Ireland: a review of scientific and policy advances since 1988. Vet Microbiol 112:239–251
Muñoz-Igualada J, Shivik JA, Domínguez FG, Lara J, González LM (2008) Evaluation of cage-traps and cable restraint devices to capture red foxes in Spain. J Wildl Manage 72:830–836
Murphy D, O’Keeffe JJ, Martin SW, Gormley E, Corner LAL (2009) An assessment of injury to European badgers (Meles meles) due to capture in stopped restraints. J Wildl Dis 45:481–490
O’Keeffe JJ (2006) Description of a medium term national strategy toward eradication of Tuberculosis in cattle in Ireland. In: Proceedings of the 11th Symposium of the International Society of Veterinary Epidemiology and Economics, Cairns, Australia. p. 502. http://www.sciquest.org.nz/node/64117. Accessed on 10th April 2012
O’Connor CM, Haydon DT, Kao RR (2012) An ecological and comparative perspective on the control of bovine tuberculosis in Great Britain and the Republic of Ireland. Prev Vet Med 104:185–197
O’Mairtin D, Williams DH, Griffin JM, Dolan LA, Eves JA (1998) The effect of a badger removal programme on the incidence of tuberculosis in an Irish cattle population. Prev Vet Med 34:47–56
Pan W (2001) Akaike’s information criterion in generalized estimating equations. Biometrics 57:120–125
Reid N, Etherington TR, Wilson G, McDonald RA, Montgomery WI (2008) Badger survey of Northern Ireland 2007/08. Quercus and Central Science Laboratory for the Department of Agriculture & Rural Development (DARD), Northern Ireland
Reid N, Etherington TR, Wilson GJ, Montgomery WI, McDonald RA (2011) Monitoring and population estimation of the European badger (Meles meles) in Northern Ireland. Wildl Biol 18(1):46–57
Roper T (2010) Badger. Harpercollins Pub Ltd., UK
Sadlier L, Montgomery WI (2004) The impact of sett disturbance on badger Meles meles numbers; when does protective legislation work? Biol Cons 119:455–462
Sadlier LMJ, Webbon CC, Baker PJ, Harris S (2004) Monitoring foxes (Vulpes vulpes) and badgers (Meles meles): are field signs the answer? Mamm Rev 34:75–98
Sheridan M (2011) Progress in tuberculosis eradication in Ireland. Vet Microbiol 151:160–169
Sleeman DP, Davenport J, More SJ, Clegg TA, Collins JD et al (2009) How many Eurasian badgers Meles meles L. are there in the Republic of Ireland? Euro J Wildl Res 55:333–344
Szmaragd C, Green L, Medley G, Mitchell A, Browne W. (2010) Estimating badger numbers from badger survey signs, with applications to bovine TB prediction. Soc Vet Epidemiol Prev Med (SVEPM): Proc, Nantes, France
Tuyttens FAM, Macdonald DW, Delahay R, Rogers LM, Mallinson PJ et al (1999) Differences in trappability of European badgers Meles meles in three populations in England. J Appl Ecol 36:1051–1062
Tuyttens FAM, Long B, Fawcett T, Skinner A, Brown JA, Cheeseman CL, Roddam AW, Macdonald DW (2001) Estimating group size and population density of Eurasian badgers Meles meles by quantifying latrine use. J Appl Ecol 38:1114–1121
Wilson GJ, Delahay RJ, de Leeuw ANS, Spyvee PD, Handoll D (2003) Quantification of badger (Meles meles) sett activity as a method of predicting badger numbers. J Zool 259:49–56
Wilson GJ, Carter SP, Delahay RJ (2011) Advances and prospects for management of TB transmission between badgers and cattle. Vet Microbiol 151:43–50
Woodroffe R, Gilks P, Johnston WT, Le Fevre AM, Cox DR et al (2008) Effects of culling on badger abundance: implications for tuberculosis control. J Zool 274:28–37
Acknowledgments
AWB was supported by a Walsh Fellowship from Teagasc—The Irish Agriculture and Food Development Authority (www.teagasc.ie). All data were generated during licensed bovine TB control activities funded through the Department of Agriculture, Fisheries and Food (www.agriculture.gov.ie) as part of the medium term national strategy toward eradication of bovine TB. The authors wish to thank Stuart Green (Teagasc), Dr. Liam Lysaght and the staff of the National Biodiversity Data Centre for their support during the writing of this paper. Gratitude is extended to all the field staff in Longford, Monaghan and South Tipperary and administrative staff within the Wildlife Unit of the Department of Agriculture, which contributed to data collection during this project. We also wish to thank all land owners for cooperating during the fieldwork of this study. The manuscript was greatly improved by comments from two anonymous reviewers.
Competing interests
The authors have declared that no competing interests exist.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gortázar
Rights and permissions
About this article
Cite this article
Byrne, A.W., O’Keeffe, J., Sleeman, D.P. et al. Impact of culling on relative abundance of the European badger (Meles meles) in Ireland. Eur J Wildl Res 59, 25–37 (2013). https://doi.org/10.1007/s10344-012-0643-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-012-0643-1