
Abstract
Ungulate overabundance can strongly alter composition and structure of forest communities through selective damage independently of the herbivory history of the recipient system. In the early 1900s, red and fallow deer (Cervus elaphus and Dama dama, respectively) were introduced to northwestern Patagonia. We studied patterns of relative habitat use, damage, and species selection by introduced deer populations in three major plant communities that have the longest history of invasion in the region. We also evaluated community structure and composition characteristics as possible explanatory factors for the observed patterns of browsing, fraying, and bark-stripping. At the species level, exotic deer browsed more than expected on two evergreen species (Austrocedrus chilensis and Schinus patagonicus) and two spiny species (Colletia hystrix and Dasyphyllum diacanthoides), while one dominant shrubland tree (Lomatia hirsuta) was selected for fraying and bark-stripping. Browsing was the more widespread damage in all plant communities, while fraying and bark-stripping occurred at low incidence. Furthermore, species identity was found as the main driver of deer damage over plant community type. Finally, variation in damage and in habitat use was explained by community structure and composition characteristics. Bark-stripping and fraying are best predicted by community composition, whereas browsing and habitat use are best predicted by structural properties including understory cover, cover of fallen logs, and tree density. The patterns of damage and the association with community structure characteristics reported here provide insight for developing management strategies.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Invasive species can severely damage native ecosystems and have consequently become one of the greatest threats to global biodiversity (Mack et al. 2000; Vitousek et al. 1997). Independently of the evolutionary history of herbivory in an invaded system, introduced herbivores at high densities impose a novel type and level of herbivory when they persist in the landscape (Hobbs and Huenneke 1992). Selective browsing often affects vegetation by reducing sapling growth and survival, deforming stems, and inducing chemical changes, all of which can eventually cause regeneration to fail and thus can eliminate some species from the community (Côté et al. 2004; Gill 1992a, b; Rooney and Waller 2003; Vila et al. 2003). For example, in New Zealand, where plant species are poorly adapted to browsing, the introduction of deer more than a hundred years ago has caused dramatic changes in the native flora (Bellingham and Lee 2006; Coomes et al. 2003; Forsyth et al. 2010; Husheer et al. 2003; Nugent et al. 2001) as well as in ecosystem structure and function (Forsyth et al. 2010; Wardle et al. 2001).
In northwestern Patagonia, red and fallow deer (Cervus elaphus and Dama dama, respectively) were introduced a century ago (Jaksic 1998; Jaksic et al. 2002; Novillo and Ojeda 2008; Vazquez 2002). In Argentina, fallow deer are confined to Isla Victoria and the Huemul Peninsula, whereas red deer have become invasive and currently inhabit a wide variety of habitats across Patagonia (Flueck et al. 2003). Detrimental effects of red deer on the structure and composition of some native communities have been demonstrated for temperate forests of southern Argentina (Relva and Veblen 1998; Relva et al. 2009, 2010; Vazquez 2002; Veblen et al. 1989, 1992) and Chile (Ramirez et al. 1981). However, these studies have considered browsing as the sole type of damage by deer, and information about the frequency and magnitude of other deer damage such as fraying and bark-stripping is currently lacking. Fraying is the removal of bark by the action of rubbing antlers up and down the stem, and bark-stripping is the removal of bark with the animal’s teeth (Gill 1992a). Both activities can have negative ecological (Faber and Thorson 1996; Motta 1996) and economic impacts (Faber and Edenius 1998; Gill 1992b; Welch and Scott 2008). Fraying and bark-stripping are highly selective (Gill 1992b; Motta 1996) and result in reduction in growth rates, tree survival, and timber quality, plus increased exposure to fungal infection (Faber and Edenius 1998; Rooney and Waller 2003; Welch and Scott 2008). The incidence and intensity of tree damage depends on the tree species, community structure and composition, and deer density among other variables (Gill 1992a). Together with browsing, fraying and bark-stripping can profoundly affect plant community composition and dynamics (Cotè et al. 2004; Gill 1992b; Rooney and Waller 2003; Russell et al. 2001) as well as biodiversity (Martin et al. 2010; van Wieren and Bakker 2008). Therefore, quantifying the extent and intensity of different types of damage by introduced deer is required to identify the most susceptible communities and to prioritize deer control measures.
Large herbivores interact with forage resources in a heterogeneous environment, selecting for plant species in communities and plant communities in the landscape (Skarpe et al. 2000; Stockton et al. 2005). Diet selection is based in part on palatability, while selection for plant communities is generally a linear function of the relative abundance of the preferred plant species in the community (Senft et al. 1987). The differential consumption of species can be estimated through diet studies that include analysis of feces or stomachs, direct animal observation, or recording traces on vegetation. However, the latter has been suggested to be more relevant to woody species because browsing traces persist on plants and can be accurately evaluated (Boulanger et al. 2009). Previous studies in northwestern Patagonia show that fallow and red deer rely on woody species for their year-round diet, with use of graminoids and herbs increasing during spring and summer (Bahamonde 1984; Galende and Grigera 1998; Ortiz and Bonino 2007; Relva and Caldiz 1998). However, none of these studies reflect species selection because they do not consider diet composition relative to the availability of the species in the communities. Determining whether deer are browsing each species proportionally to its availability and how browsing selection varies among diverse plant communities may also have important implications for browsing management. This information will allow managers to identify susceptible plant species or communities on which management efforts should focus. Additionally, many factors have been related to the occurrence and intensity of damage. Biotic factors such as deer density are positively related to damage (Gill and Morgan 2010; Horsley et al. 2003; Kay 1993), but tree density, canopy closure, stand composition, and understory abundance have also been observed to be closely related to damage (Gill 1992a; Moore et al. 1999). Predicting where severe damage is likely to occur is necessary for risk assessment and also for the development of management polices (Spear and Chown 2009).
In this paper, we seek: (1) to quantify damage by red and fallow deer and to determine selectivity of browsing, fraying, and bark-stripping of plant species and native plant communities, (2) to identify patterns of relative habitat use (via pellet and paths counts), and (3) to determine plant community characteristics that are related to use and damage by deer in order to assess community susceptibility. The long history of establishment by introduced deer on Isla Victoria, Patagonia, Argentina, makes this island an ideal natural experiment for assessing long-term impacts of a well-established and invasive deer population on temperate forests of the southern Andes. We predict that (1) deer damage is species-, size-, and community-specific through deer selection; (2) deer differentially use plant communities as they provide different structure and composition characteristics relevant to foraging and cover, and (3) the structure and composition of the community, understory cover, and accessibility to the site are the main factors related to damage and use by deer.
Methods
Study area
This research was conducted on Isla Victoria, an island of 3710 ha in Lake Nahuel Huapi, Nahuel Huapi National Park, Argentina (40°57′S, 71°33′W). This island is mainly covered by pure southern beech Nothofagus dombeyi forests, pure Austrocedrus chilensis forests, and mixed N. dombeyi–A. chilensis forests. Shrublands are co-dominated by Lomatia hirsuta, Maytenus boaria, and Nothofagus antarctica, while Luma apiculata, Myrceugenia exsucca, and Dasyphillum diacanthoides are subdominant species in these communities. The understory is diverse, with shrubs, ferns, herbs, bamboo, and other graminoids. Between 1917 and 1922 red and fallow deer were introduced to Isla Victoria (Anziano 1962), and currently combined density is estimated through the clearance dung plot method to be as high as 26 deer/km2 (Relva, unpublished data). In addition, in 1999 wild boar (Sus scrofa) arrived on the island and are currently found throughout the island (Barrios-Garcia, unpublished data). The only native ungulate on the island is pudú (Pudu pudu), which have become very rare, likely owing to human activities (Vazquez 2002). Therefore, herbivory and track signs registered during this study correspond only to red and fallow deer. There is no predator on the island, and hunting is performed only by park rangers to control deer populations, but not systematically. Thus, the population is self-regulated and attains very high densities.
Study design
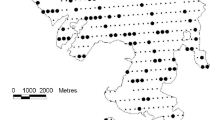
We studied patterns of use and damage by deer in three major community types by surveying 18 randomly selected sites at least 1 km apart (Fig. 1) during the summers of 2004 and 2005. The communities were forests dominated by A. chilensis (hereafter, A. chilensis forests), forests dominated by Nothofagus dombeyi (hereafter, N. dombeyi forests) and shrublands dominated by M. boaria–L. hirsuta (hereafter, shrublands). At each site (six for each plant community) we systematically recorded overstory and understory structure and composition as well as deer damage and use in 20 circular plots with centers spaced 30 m apart. The circular plots had different radii depending on which variables were being measured. We used 150 m2 plots to identify adult tree species and to measure the diameters at breast height (dbh; stems ≥ 4.0 cm dbh) as well as presence, type, and level of damage (fraying and bark-stripping). In addition, we recorded canopy cover with a densiometer and slope with a compass. In 50-m2 circular plots, within the larger 150-m2 plots, we recorded sapling (stems < 4.0 cm dbh) density, height, and accumulated browsing (intensity: 0—no browsing; 1—light, 1–25% of the branches browsed; 2—moderate, 25–75% of the branches browsed; and 3—heavy, 75–100% of the branches browsed). We also recorded relative cover of all vascular species using the Braun–Blanquet modified scale (0%, <1%, 1–5%, 6–25%, 26–50%, 51–75%, 75–100%). Use of this scale is by visual estimation of the proportion of the ground surface occupied by a single species (or group of them) in a plot. We also measured height and recorded accumulated browsing of each of the woody species in the understory. Plant species were identified according to Ezcurra and Brion (2005). The degree of site accessibility to deer was estimated by measuring the maximum height of fallen trees and by estimating their cover as was done in the understory plots. Finally, in order to determine relative habitat use we counted pellet groups in 10 m2 plots and deer paths in 150 m2 plots. We considered a pellet group to be more than six intact pellets voided in the same defecation (Mayle et al. 1999), and all groups were counted regardless of decomposition stage, thus recording cumulative deer presence in the site. All measurements were taken during the same visit to the sites and no significant changes occurred in weather or management across the years of sampling that could affect deer densities or behavior.

Map of Isla Victoria with study sites
Data analysis
A selection index (Motta 1996; Petrides 1975; Skarpe et al. 2000) was calculated for species and plant communities and compared to incidence of all types of damage. We use the term “selection” instead of “preference” because measuring preference requires food types to be equally available (Norbury and Sanson 1992). Selected species are those used at a higher frequency than their availability in the environment. The selection index (SI) is:
where D i is the number of damaged saplings/trees of species i, D t is the total number of damaged saplings/trees of all species, A i is the number of available saplings/trees of species i, and A t is the total number of available saplings/trees of all species. The index centers on 1 as a reference point (the species is neither selected nor avoided; that is, the species is eaten in proportion to its abundance in the field). Species with SI values above 1 are selected and below 1 are avoided. By contrast, incidence refers to the proportion of damaged saplings/trees of species i to the number of available saplings/trees of that species, and intensity refers to the average of the accumulated browsing of particular species (scale 0 to 3). The statistical significance of the selection index was tested by comparing the statistic \( {\text{Savage}} = \frac{{{{({\text{SI}} - 1)}^2}}}{{{\text{SE(SI}}{)^2}}} \) with the critical value of a χ 2 distribution with 1 degree of freedom (Manly et al. 1993). The standard error of the index was calculated by\( {\text{SE(SI}}) = \sqrt {{\frac{{1 - ({{{{A_i}}} \left/ {{{A_{\text{t}}}}} \right.})}}{{Dt*({{{{A_i}}} \left/ {{{A_{\text{t}}}}} \right.})}}}} \). The selection index was also calculated for plant communities; in this case, D i is the number of damaged saplings/trees in plant community i, D t is the number of damaged saplings/trees in all plant communities, A i is the number of available saplings/trees in plant community i, and A t is the total number of available saplings/trees in all plant communities. As with the species index, values above 1 indicate selection of a community and below 1 avoidance; significance was tested with the Savage statistic.
To identify differences in deer use (via pellet and path counts) among plant communities (A. chilensis forests, N. dombeyi forests, and shrublands; n = 6 for each plant community), we performed a one-way analysis of variance (ANOVA) and post-hoc Tukey’s tests. Using a two-way ANOVA and post-hoc Tukey’s tests, we evaluated (1) the effect of species and plant community type on the incidence and selection index of the different deer damages and (2) if selection at species level changes with plant community (interaction effect) (Hjältén et al. 1993). Additionally, we evaluated the strength of the relations between different types of damage and deer use indicators with Spearman correlation coefficients (n = 18). Finally, we used stepwise multiple regressions to identify forest attributes that account for variation in use and damage by deer (n = 18). We used the different types of damage (browsing, bark-stripping, and fraying) and deer use indicators (number of pellets and paths) as response variables and structural and compositional characteristics as predictors. In particular, we used the average of overstory attributes (basal area, tree density, canopy cover), composition (basal area of the dominant tree in each community, i.e., N. dombeyi, A. chilensis, and M. boaria–L. hirsuta), and understory traits (height and density of saplings, cover and height of woody understory and herb cover). Counts (number of pellet groups and paths) and measures (heights and basal area) were log-transformed and proportions (understory cover and incidence of damage) were arcsine square-root transformed to meet normality and homoscedasticity requirements. Because damage cannot be attributed to a particular deer species, results refer to the combined damage by both red and fallow deer.
Results
Introduced deer selectivity at individual and species levels
In all three plant communities, deer used some plant species at a higher frequency than their availability in the environment. The selection index showed that deer browsed more than expected on two evergreen species, A. chilensis and Schinus patagonicus, and two spiny species, Colletia hystrix and Dasyphyllum diacanthoides (Table 1). However, the Savage statistic showed only that deer significantly avoided N. dombeyi (Table 1). Similarly, the selection index showed that fraying occurred selectively on two species, S. patagonicus and L. hirsuta, although deer significantly avoided only M. boaria and N. dombeyi (Table 2). Bark-stripping occurred selectively and significantly on only one species, L. hirsuta, while Aristotelia chilensis and S. patagonicus were significantly avoided (Table 2). Incidence and intensity of all types of damage were highest for the same species that deer selected (Tables 1 and 2). Deer also selected particular tree diameters: small stems (mode of 2.10 cm for saplings and 5.00 cm for adult trees) were used for fraying and relatively larger diameters for bark-stripping (17.00 cm), and the resulting scars varied in length according to the type of damage; fraying scars were longer than the relatively smaller scars produced by bark-stripping (66.80 ± 3.30 and 51.20 ± 2.20 cm, respectively). Overall, 5.50% of adult trees were either frayed or bark-stripped, of which 17.00% presented more than one scar, implying multiple damage events.
Use and damage by deer across plant communities
Pellet and path densities were similar among plant communities (one-way ANOVA (F 2,15 = 0.93; p = 0.42) and (F 2,15 = 0.29; p = 0.75), respectively; Table 3). The two-way ANOVA for both selection index and incidence of deer browsing indicated differences owing to species (two-way ANOVA, F = 4.96, p < 0.01 and F = 4.18, p < 0.01, respectively), but not owing to plant community (two-way ANOVA, F = 0.49, p = 0.48 and F = 0.68, p = 0.41, respectively; Table 3), or to the interaction between them (two-way ANOVA, F = 0.53, p = 0.91 and F = 0.52, p = 0.92, respectively). A post-hoc Tukey’s test indicates that the species significantly browsed are the same as those found as selected by the selection index (i.e., A. chilensis, C. hystrix, D. diacanthoides, L. apiculata, N. dombeyi, and S. patagonicus,). Similarly, the selection index and incidence of fraying varied significantly by species (two-way ANOVA, F = 6.47, p < 0.01 and F = 3.38, p < 0.01, respectively) but did not differ by community (two-way ANOVA, F = 1.21, p = 0.30 and F = 2.20, p = 0.11, respectively; Table 3) or the interaction between them (two-way ANOVA, F = 1.18, p = 0.30 and F = 0.90, p = 0.55, respectively). Likewise, for bark stripping, the selection index and incidence indicated differences owing to species (two-way ANOVA, F = 4.72, p < 0.05 and F = 9.23, p < 0.01, respectively), but not owing to plant community (two-way ANOVA, F = 1.55, p = 0.23 and F = 0.79, p = 0.46, respectively; Table 3), or to the interaction between them (two-way ANOVA, F = 0.96, p = 0.13 and F = 1.81, p = 0.15, respectively). Finally, in agreement with the selection index, the post-hoc Tukey’s test indicated that L. hirsuta is significantly more damaged by both fraying and bark-stripping than the other species.
Community factors related to damage
There were significant and distinct associations among different types of damage and forest characteristics. Areas with higher deer use as shown by pellet densities suffered higher bark-stripping and fraying damage, while areas with higher path density showed higher sapling and understory browsing (p < 0.01 and p < 0.05, respectively; Table 4). In addition, the multiple regression analyses showed that fraying and bark-stripping are related to plant community structural and compositional characteristics, whereas browsing and deer use (pellets and paths) are related only to structural characteristics (Table 5). Seventy-four percent of the variance in fraying is related to tree density (+) and the basal area of N. dombeyi (−) (p < 0.01). Bark-stripping was positively related to the basal area of M. boaria–L. hirsuta and canopy density and negatively to the basal area of uprooted trees and woody cover (r 2 = 0.78, p < 0.01). Fifty-two percent of the variance in browsing was explained by sapling cover (−) and cover by fallen logs (+) (p < 0.01). For deer use indicators, pellet density was positively related to tree density and negatively to woody cover (r 2 = 0.67, p < 0.01), whereas path density was positively related to canopy density and negatively related to sapling density (r 2 = 0.36, p < 0.05; Table 5).
Discussion
In light of the current spread of exotic deer in forests in Patagonia and the expectation that the invasion process will continue in the next decades (Flueck et al. 2003), understanding how deer use and damage plant communities is critical to managing potential deer impacts, as is determining the main factors related to this damage. Our findings indicate that 100 years after introduction to Isla Victoria, deer roam and browse in the three main native plant communities, but they used some plant species more frequently than others as well as selecting for sizes for each type of damage. Additionally, community structure and composition seem to be important factors for the prediction of damage and thus provide further insight for developing management strategies.
Browsing was the most extensive and intensive damage on native plant communities. Deer browsed most of the 30 woody species recorded in this study; however, deer browsed more than expected on two evergreen species (A. chilensis and S. patagonicus) and two spinescent species (C. hystrix and D. diacanthoides). These findings are partially consistent with previous studies in the region, in which A. chilensis and S. patagonicus were described as heavily browsed by deer (Veblen et al. 1989, 1992; Galende and Grigera 1998; Relva and Veblen 1998). In addition, recent studies showed that exotic deer strongly delay sapling growth of one of the dominant trees, A. chilensis, possibly facilitating the invasion of an exotic invasive tree species, Pseudotsuga menziesii (Relva et al. 2010). However, both evergreen and spiny plants are often adduced as having high content of fiber and chemical deterrents to deer (Skarpe and Hester 2008). Selection for highly defended plant species by deer has been previously documented (Cooper and Owen-Smith 1986). The selection of these presumably unpalatable species could be interpreted as indicating depletion of more palatable species owing to high herbivory pressure. Moreover, deer produce salivary tannin-binding proteins that may minimize the detrimental effects of defences present in plants (Austin et al. 1989). This finding might help to explain the fact that the Savage statistic was usually not significant for the selected species. It seems that the selected species are not significantly used but rather that deer forage on the best of the quite poor species available in this impoverished situation. Furthermore, Aristotelia chilensis, a species highly susceptible to browsing (Blackhall et al. 2008; Veblen et al. 1992), was not determined to be a selected species in our study. Presumably, historically heavy deer browsing may have caused Aristotelia chilensis saplings to become rare (Relva et al. 2010; Vazquez 2002; Veblen et al. 1989). Our field observations suggest that the population might by split into two groups, established individuals with heights above the browsing line, and individuals found in “natural refuges” such as amidst fallen trees. This pattern was also observed by Relva et al. (2009), where cover of palatable species was associated with severity of A. chilensis canopy mortality. Additionally, it has been suggested that intense deer browsing can change understory composition, increasing the abundance of browse-resistant species and reducing the browse-sensitive species (Relva et al. 2010; Veblen et al 1989). However, to date, we have no information about the nutritional quality of the selected and avoided plant species; thus further research should investigate the role of plant palatability in the selection process.
Fraying and bark-stripping by introduced deer were also selective in terms of species and individual size. Deer almost exclusively frayed L. hirsuta of smaller diameters and stripped bark from those with larger diameters. This finding may suggest that L. hirsuta has certain physical and chemical characteristics that make it uniquely susceptible to damage. Previous studies elsewhere have shown that physical characteristics such as size appeared to be more important than tree species identity for fraying (Ramos et al. 2006; Vila et al. 2004), while bark-stripping would be driven by nutrient content of the bark (Faber and Edenius 1998) or the ease with which bark can be peeled and the low lignin content of bark (Akashi and Nakashizuka 1999). However, no previous records on these subjects are available for Patagonia, so future research should explore the underlying mechanisms causing this pattern. While more than 50% of recorded saplings were browsed, fraying and bark-stripping occurred to a very limited extent (2.21% and 3.38% of recorded individuals, respectively). These results align with others reporting similar levels of fraying and bark-stripping incidence (Bergqvist et al. 2001; Faber and Thorson 1996). For this region, the low incidence of the damage and the fact that L. hirsuta is a vigorous resprouter suggest that the consequences of fraying and bark-stripping on the structure of these native communities might be negligible.
Deer relative use estimated through pellet group counts and the numbers of paths was similar among the three plant communities. This pattern is surprising in light of the fact that plant communities differ substantially in terms of structure and composition (Table 3) but coincides with the pattern observed for all deer damage. Furthermore, we found that species identity is the main driver of deer damage over plant community and that there is no interaction between species identity and community type. Factors such as type and density of ground cover and size and shape of the plots influence pellets counts (Neff 1968). However, the technique of standing crop pellet accumulation has been successfully used in the assessment of habitat use by deer (Putman 1984). Additionally, the use of the small plots and the same observer throughout the study reduces the chance of overlooking pellet groups (Neff 1968). The persistence of deer pellets is determined by moisture and canopy (Harestad and Bunnell 1987), but studies in the region indicate that the standing crop of pellet groups can be considered to represent deposition from 1 year (Flueck et al. 1999). Therefore, even though we did not consider the possible seasonality of deer activity, our observations account for the cumulative use of the different plant communities throughout the year.
Plant community structure and composition characteristics can be important factors for the prediction of future deer damage. Compositional traits are important determinants of fraying and bark-stripping, while structural characteristics are more important in determining browsing and relative habitat use. Deer strongly avoided N. dombeyi forests and selected sites with high tree density for fraying, while bark-stripping was best predicted by the presence of L. hirsuta and M. boaria. These facts indicate that plant community composition is of greater importance than community structure in predicting fraying and bark-stripping. In contrast, sites with high cover of fallen trees and low sapling cover are threatened by severe deer browsing; whereas sites with high tree density, canopy density, and understory cover were more used (pellets and paths). These community characteristics and the fact that continual impacts of deer could potentially inhibit the development of forest cover or alter successional pathways (Gill 1992b; Relva et al. 2010; Veblen et al. 1989) suggest that shrublands might be most vulnerable and should be a priority in the design of management strategies.
Although this research was conducted on an island, our results reflect the consequences of the oldest introduction in the region and the communities with the highest deer densities. The observed damage is the result of two different deer species with overlapping ranges. Pooling the impact of red and fallow deer could be problematic; however, in this system it is not possible to differentiate them based on the indicators we used. Because bite size is similar for both species and pellet sizes also overlap for most of their lifetime, the identification of browsing patterns by individual deer species is possible only through direct observation, when the two species occupy similar but non-overlapping ranges, or by use of stomach content analyses (Gill 1992a; Neff 1968). It is possible that effects of each herbivore in isolation might be different (Allcock and Hik 2004). However, the similarity found between browsed species in this study and previous studies on red deer in the region (Bahamonde 1984; Galende and Grigera 1998; Ortiz and Bonino 2007; Relva and Caldiz 1998; Relva and Veblen 1998; Relva et al. 2009, 2010; Vázquez 2002; Veblen et al. 1989, 1992) suggest that the two species could behave similarly. Our findings suggest that deer in northwest Patagonia are selective for species and community types for browsing, fraying, and bark-stripping. This study is the first to record all patterns of damage by introduced deer among native communities in the region and should alert managers to the potential impact deer might have on the mainland, encouraging the design of effective management plans.
References
Akashi N, Nakashizuka T (1999) Effects of bark-stripping by Sika deer (Cervus nippon) on population dynamics of a mixed forest in Japan. For Ecol Manag 113:75–82
Allcock KG, Hik DS (2004) Survival, growth, and escape from herbivory are determined by habitat and herbivore species for three Australian woodland plants. Oecologia 138:231–241
Anziano AF (1962) Acción de los animales sobre la flora. Tomo IX, pp 107–112 en Anales de Parques Nacionales, Argentina
Austin PJ, Suchar LA, Robbins CT, Hagerman AE (1989) Tannins binding proteins in saliva of deer and their absence in saliva of sheep and cattle. J Chem Ecol 15:1135–1347
Bahamonde N (1984) Dieta primaveral y estival del guanaco, ciervo colorado y ovino en un área precordillerana patagónica. Departamento de Ecología, S. C. de Bariloche, Universidad Nacional del Comahue: 44
Bellingham P, Lee W (2006) Distinguishing natural processes from impacts of invasive mammalian herbivores. In: Allen RB, Lee WG (eds) Biological invasions in New Zealnd, Series: Ecological studies, vol. 186. Springer, Berlin, pp 323–336
Bergqvist G, Bergström R, Edenius L (2001) Patterns of stem damage by moose (Alces alces) in young Pinus sylvestris stands in Sweden. Scand J For Res 16:363–370
Blackhall M, Raffaele E, Veblen TT (2008) Cattle affect early post-fire regeneration in a Nothofagus dombeyi–Austrocedrus chilensis mixed forest in northern Patagonia, Argentina. Biol Conserv 141:2251–2261
Boulanger V, Baltzinger C, Saïd S, Ballon P, Picard JF, Dupouey JL (2009) Ranking temperate woody species along a gradient of browsing by deer. For Ecol Manag 258:1397–1406
Coomes DA, Allen RB, Forsyth DM, Lee WG (2003) Factors preventing the recovery of New Zealand forests following control of invasive deer. Conserv Biol 17:450–459
Cooper SM, Owen-Smith N (1986) Effects of plant spinescence on large mammalian herbivores. Oecologia 68:446–455
Côté SD, Rooney TP, Tremblay JP, Dussault C, Waller DM (2004) Ecological impacts of deer overabundance. Annu Rev Ecol Evol Syst 35:113–147
Ezcurra C and Brion C (2005) Plantas del Nahuel Huapi. Catalogo de la Flora Vascular del Parque Nacional Nahuel Huapi, Argentina. San Carlos de Bariloche, Universidad Nacional del Comahue y Red Latinoamericana de Botánica
Faber WE, Edenius L (1998) Bark stripping by moose in commercial forests of Fennoscandia- a review. Alces 34:261–268
Faber WE, Thorson EM (1996) Bark stripping of young Pinus sylvestris by Alces alces on the individual, stand, and landscape level in Sweden. Can J For Res 26:2109–2114
Flueck WT, Franken M, Smith-Flueck JAM (1999) Red deer, cattle and horses at high elevations in the Andean precordillera: habitat use and deer density. Mastozool Neotrop 6(2):91–101
Flueck WT, Smith-Flueck JAM, Naumann CM (2003) The current distribution of red deer (Cervus elaphus) in southern Latin America. Z Jagdwiss 49:112–119
Forsyth DM, Wilmshurst JM, Allen RB, Coomes DA (2010) Impacts of introduced deer and extinct moa on New Zealand ecosystems. NZ J Ecol 34:48–65
Galende GI, Grigera D (1998) Trophic relationships of Lagidium viscacia (Rodentia, Chinchillidae) with herbivores introduced in the Nahuel Huapi National Park, Argentina. Iheringia, Ser Zoologia, 3–10
Gill RMA (1992a) A review of damage by mammals in north temperate forests: 1. Deer. Forestry 65:145–169
Gill RMA (1992b) A review of damage by mammals in north temperate forests: 3. Impact on trees and forests. Forestry 65:363–388
Gill RMA, Morgan G (2010) The effects of varying deer density on natural regeneration in woodlands in lowland Britain. Forestry 83(1):53–63
Harestad AS, Bunnell FL (1987) Persistence of black-tailed deer fecal pellets in coastal habitats. J Wildl Manag 51:33–37
Hjältén J, Danell K, Lundberg P (1993) Herbivore avoidance by association: vole and hare utilization of woody plants. Oikos 68:125–131
Hobbs RJ, Huenneke LF (1992) Disturbance, diversity and invasion: implications for conservation. Conserv Biol 6(3):324–337
Horsley SB, Stout SL, DeCalesta DS (2003) White-tailed deer impact on the vegetation dynamics of a northern hardwood forest. Ecol Appl 13:98–118
Husheer SW, Coomes DA, Robertson AW (2003) Long-term influences of introduced deer on the composition and structure of New Zealand Nothofagus forests. For Ecol Manag 181:99–117
Jaksic FM (1998) Vertebrate invaders and their ecological impacts in Chile. Biodivers Conserv 7:1427–1445
Jaksic FM, Iriarte JA, Jiménez JE, Martínez DR (2002) Invaders without frontiers: cross-border invasions of exotic mammals. Biol Invasions 4:157–173
Kay S (1993) Factors affecting severity of deer browsing damage within coppiced woodlands in the south of England. Biol Conserv 63:217–222
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecol Appl 10:689–710
Manly BFJ, McDonald L, Thomas D (1993) Resource selection by animals: Statistical design and analysis for field studies. Chapman and Hall, London
Martin JL, Stockton SA, Allombert S, Gaston AJ (2010) Top-down and bottom-up consequences of unchecked ungulate browsing on plant and animal diversity in temperate forests: lessons from a deer introduction. Biol Invasions 12:353–371
Mayle BA, Peace AJ, Gill RMA (1999) How many deer? A field guide to estimating deer population size. Forestry Commission Field Book 18. Forestry Commission, Edinburgh
Moore NP, Hart JD, Langton SD (1999) Factors influencing browsing by fallow deer Dama dama in young broad-leaved plantations. Biol Conserv 87:255–260
Motta R (1996) Impact of wild ungulates on forest regeneration and tree composition of mountain forests in the Western Italian Alps. For Ecol Manag 88:93–98
Neff DJ (1968) The pellet-group count technique for big game trend, census, and distribution: a review. J Wildl Manag 32:597–614
Norbury GL, Sanson GD (1992) Problems with measuring diet selection of terrestrial, mammalian herbivores. Austral Ecol 17:1–7
Novillo A, Ojeda RA (2008) The exotic mammals of Argentina. Biol Invasions 10:1333–1344
Nugent G, Fraser W, Sweetapple P (2001) Top down or bottom up? Comparing the impacts of introduced arboreal possums and terrestrial ruminants on native forests in New Zealand. Biol Conserv 99:65–79
Ortiz C, Bonino NA (2007) Dieta estacional del ciervo colorado (Cervus elaphus) en el Parque Nacional Nahuel Huapi, Argentina. Ecología Austral 17:281–286
Petrides GA (1975) Principal foods versus preferred foods and their relations to stocking rate and range condition. Biol Conserv 7:161–169
Putman RJ (1984) Facts from faeces. Mamm Rev 14:79–97
Ramirez C, Godoy R, Eldridge W, Pacheco N (1981) Impacto ecológico del ciervo rojo sobre el bosque de olivillo en Osorno, Chile. An Mus Hist Nat Valparaíso 14:197–215
Ramos JA, Bugalho MN, Cortez P, Iason GR (2006) Selection of trees for rubbing by red and roe deer in forest plantations. For Ecol Manag 222:39–45
Relva MA, Caldiz MS (1998) Composición estacional de la dieta de ciervos exóticos en Isla Victoria, P. N. Nahuel Huapi, Argentina. Gayana Zool 62(2):101–108
Relva MA, Veblen TT (1998) Impacts of introduced large herbivores on Austrocedrus chilensis forests in northern Patagonia, Argentina. For Ecol Manag 108:27–40
Relva MA, Westerholm CL, Kitzberger T (2009) Effects of introduced ungulates on forest understory communities in northern Patagonia are modified by timing and severity of stand mortality. Plant Ecol 201:11–22
Relva MA, Nuñez MA, Simberloff D (2010) Introduced deer reduce native plant cover and facilitate invasion of non-native tree species: evidence for invasional meltdown. Biol Invasions 12:303–311
Rooney TP, Waller DM (2003) Direct and indirect effects of white-tailed deer in forest ecosystems. For Ecol Manag 181:165–176
Russell FL, Zippin DB, Fowler NL (2001) Effects of white-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review. Am Midland Nat 146:1–26
Senft RL, Coughenour MB, Bailey DW, Rittenhouse LR, Sala OE, Swift DM (1987) Large herbivore foraging and ecological hierarchies. BioScience 37:789–799
Skarpe C, Hester A (2008) Plant traits, browsing and grazing herbivores, and vegetation dynamics. In: Gordon IJ, Prins HHT (eds) The ecology of browsing and grazing. Springer, Berlin, pp 217–261
Skarpe C, Bergstrom R, Braten AL, Danell K (2000) Browsing in a heterogeneous savanna. Ecography 23:632–640
Spear D, Chown SL (2009) Non-indigenous ungulates as a threat to biodiversity. J Zool 279:1–17
Stockton SA, Allombert S, Gaston AJ, Martin JL (2005) A natural experiment on the effects of high deer densities on the native flora of coastal temperate rain forests. Biol Conserv 126:118–128
van Wieren SE, Bakker JP (2008) The impact of browsing and grazing herbivores on biodiversity. In: Gordon IJ, Prins HHT (eds) The ecology of browsing and grazing. Springer, Berlin, pp 263–292
Vazquez DP (2002) Multiple effects of introduced mammalian herbivores in a temperate forest. Biol Invasions 4:175–191
Veblen TT, Mermoz M, Martin C, Ramilo E (1989) Effects of exotic deer on forest regeneration and composition in northern Patagonia. J Appl Ecol 26:711–724
Veblen TT, Mermoz M, Martin C, Kitzberger T (1992) Ecological impacts of introduced animals in Nahuel Huapi Natianal Park, Argentina. Conserv Biol 6:71–83
Vila B, Guibal F, Torre F, Martin JL (2003) Growth change of young Picea sitchensis in response to deer browsing. For Ecol Manag 180:413–424
Vila B, Guibal F, Torre F, Martin JL (2004) Assessing spatial variation in browsing history by means of fraying scars. J Biogeogr 31:987–995
Vitousek PM, Dantonio CM, Loope LL, Rejmanek M, Westbrooks R (1997) Introduced species: a significant component of human-caused global change. NZ J Ecol 21:1–16
Wardle DA, Barker GM, Yeates GW, Bonner KI, Ghani A (2001) Introduced browsing mammals in New Zealand natural forests: aboveground and belowground consequences. Ecol Monogr 71:587–614
Welch D, Scott D (2008) An estimate of timber degrade in Sitka spruce due to bark stripping by deer in a Scottish plantation. Forestry 81(4):489–497
Acknowledgments
We thank M. Rodriguez-Cabal, Y. Sasal, R Dimarco, S. Moreyra, S. Ballari, G. Barrios, M. Barrios, and C. Quintero for field assistance. Park Rangers Damián “Tata” Mujica and Lidia Serantes provide valuable help on Isla Victoria. The Delegación Técnica and Administración de Parques Nacionales, Nahuel Huapi provided permits. Cau-Cau Espacio SA provided transportation to and from Isla Victoria. Dan Simberloff, Joe Bailey, Nate Sanders, Mariano Rodriguez-Cabal, Martin Nuñez, and two anonymous reviewers provided helpful comments on earlier versions of this manuscript. This research was funded by Administración de Parques Nacionales, Nahuel Huapi, and Sigma Xi.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. Gortázar
Rights and permissions
About this article
Cite this article
Barrios-Garcia, M.N., Relva, M.A. & Kitzberger, T. Patterns of use and damage by exotic deer on native plant communities in northwestern Patagonia. Eur J Wildl Res 58, 137–146 (2012). https://doi.org/10.1007/s10344-011-0554-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10344-011-0554-6