
Abstract
Natural disturbances such as fires, windstorms, floods, and herbivory often act on plant communities, affecting their structure and the abundance and composition of their species. Most research has focused on the effects of single disturbances on plant communities whereas the synergistic effects of several disturbances have received less attention. In this study, we evaluated how timing and severity of tree mortality modified plant use by introduced deer and early post-mortality successional trajectories in northern Patagonian conifer forests. We sampled understory composition and deer use in Austrocedrus chilensis (ciprés de la cordillera) forest stands undergoing varying timing and severity of forest mortality as reconstructed using dendroecological techniques. In addition, we evaluated the effect of fallen logs on plant composition and deer use of plants by monitoring areas of massive dieback where fallen logs had been removed for fire hazard reduction, and nearby control areas not subjected to such removal. Stepwise regression analyses showed that history and severity of tree mortality strongly influence plant composition and deer use of plants. For deer use (with pellet counts and browsing index as response variables), results showed a positive relationship with degree of stand mortality and a negative relationship with cover of fallen logs. Similarly, cover of unpalatable shrub species was explained by canopy mortality history, whereas cover of palatable shrub species was positively associated with severity of canopy mortality. In areas where fallen logs had been removed, pellet counts were six times higher than those in control areas. Though total shrub species cover was similar between log removal and control areas, proportion of unpalatable shrubs increased in areas where fallen logs had been removed. In conclusion, deer use of plants was strongly limited by tall fallen logs, allowing palatable species to establish and grow. Fallen log removal accelerated deer entrance and changed understory composition toward more browse-resistant and unpalatable species. These results underscore the importance of considering the dynamics (timing, severity, and extent) of fallen woody debris influencing understory herbivory and post-disturbance succession. In addition, experimental results underpin the importance of maintaining snags and large woody debris in disturbed landscapes where salvage logging is a routine procedure.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Coarse-scale disturbances such as fires, snow avalanches, windstorms, droughts, and insect defoliation strongly influence the rate and direction of plant succession. These disturbances release limiting resources, triggering vegetation changes that attract herbivores searching the landscape for patches of high-quality forage (Jefferies et al. 1994). On the other hand, the heterogeneous matrix of dead woody debris left after forest disturbances can strongly limit and control herbivore movement (Thomas et al. 1979; Hanley et al. 1989; Nyberg 1990). Thus, plant communities will likely reflect a complex synergism of disturbance characteristics that affect plant performance directly by releasing limiting resources (Pickett and White 1985) and indirectly by modifying herbivore foraging patterns (Stuth 1991).
Although forests are highly dynamic systems subjected to natural disturbances of different scales, relatively few studies have addressed how large herbivores, such as ungulates, differentially use and impact vegetation of sites affected by forest disturbances of varying severity and timing (Wisdom et al. 2006). Ungulates generally exert minor influences on the structure and function of mature forest stands (Russell et al. 2001). However, their effect following a disturbance can determine the trajectory of the system among alternative states (Hobbs 1996; Russell et al. 2001). We hypothesized that depending on the severity and timing of the disturbances, physical and biotic conditions at disturbed sites may alter deer behaviour, thus changing their role in modifying plant succession. We predict that recent sudden, massive forest dieback events such as windstorms may create a mosaic of highly inaccessible microsites composed of a tight network of fallen logs and branches and will be dominated by palatable plants. Older, less severe or more chronic patterns of tree mortality, by contrast, may allow more accessibility, will show signs of higher deer use and will be dominated by unpalatable plants.
Forests of northern Patagonia, particularly on Isla Victoria, are ideal for evaluating forest mortality–herbivory interactions. Here, extensive stands of Austrocedrus chilensis (D. Don.) Pic. Serm. & Bizarri (ciprés de la cordillera) are being affected by “mal del ciprés”, a syndrome caused by a poorly known agent (Filip and Rosso 1999; La Manna and Rajchenberg 2004; Greslebin and Hansen 2006) that causes root death and standing mortality followed by mass canopy collapse owing to root weakening and increased susceptibility to windthrow. At the landscape scale, poor soil drainage controls the occurrence of patches of standing dead trees of diverse sizes plus logs and fallen branches on the forest floor that appear interspersed in a matrix of healthy forest (La Manna et al. 2008; Fig. 1a). Interacting with the understory and tree saplings in these forests, there are also abundant introduced cervids, mostly red deer (Cervus elaphus) and fallow deer (Dama dama) (Simberloff et al. 2003). Austrocedrus forests are heavily used by introduced deer owing to high forage availability and provision of winter cover (Relva and Caldiz 1998; Barrios Garcia Moar 2005). In addition, extensive removal of downed slash and fallen logs along roads for fire hazard mitigation (Fig. 1b) offers a unique large-scale manipulative experimental setting in which to test possible mechanisms involved in this interaction between mortality and herbivory.

Photographs showing massive mortality of Austrocedrus chilensis forests with standing dead trees, logs and fallen branches (a) and adjacent areas where logs and fallen branches were removed (b) on Isla Victoria, northern Patagonia
Here, we present results that combine dendroecological techniques for determining timing and severity of past mortality with standard vegetation and herbivore use assessments that preliminarily underscore the importance of stand decline history on understory vegetation structure and composition. In addition, we experimentally demonstrate the impact of fallen obstacles on herbivory by deer as a key mechanism in modifying the strength of herbivory effects on vegetation.
Methods
Study site
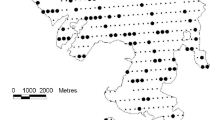
The study was conducted in a 2 × 4 km area of evergreen conifer Austrocedrus forest on northern Isla Victoria, Nahuel Huapi National Park, Argentina (40°57′S; 71°33′W; Fig. 2). Within the study area, we sampled for tree mortality reconstructions, deer use and vegetation censuses in four areas of ca. 1 ha each representing forests with contrasting history and severity of stand mortality (Criollos, Larga, Redonda, Pseudotsuga, Fig. 2, Table 1).

Location of Austrocedrus chilensis forest stands studied on Isla Victoria, Parque Nacional Nahuel Huapi, Argentina. Closed circles denote control areas and open squares denote log removal areas. See Table 1 for stand characteristics
Isla Victoria is an island running NW to SE that comprises 3,710 ha, with a varied topography that includes flat, shallow valleys, and elevations of up to 1,025 m. Mean annual rainfall is 1,700 mm (Barros et al. 1988), mostly occurring during winter (June to September). Soils are allophanic (derived from volcanic ashes), sandy, permeable, and rich in organic matter and acid pH (Koutché 1942). Isla Victoria is covered mainly by southern beech pure Nothofagus dombeyi forests, pure Austrocedrus forests, and mixed N. dombeyi-Austrocedrus forests. Lomatia hirsuta, Maytenus boaria, Nothofagus antarctica, Luma apiculata, Myrceugenia exsucca, and Dasyphyllum diacanthoides are subdominant tree species in these forests. The understory includes palatable shrubs such as Aristotelia chilensis, Maytenus chubutensis, Ribes magellanicum, Schinus patagonicus, and Chusquea culeou as well as unpalatable shrubs such as Berberis spp. and Gaultheria spp. The herbaceous layer includes native species such as Uncinia sp. and exotics such as Cynoglosum creticum and Digitalis purpurea. Species nomenclature follows Ezcurra and Brion (2005).
Historical disturbances consist of extensive fires that occurred during European settlement resulting in 80- to 120-year-old postfire-cohorts (Veblen and Lorenz 1987). These forests have scarce regeneration because the dominant tree species are not shade tolerant, although sporadic regeneration can occur in small tree-fall gaps (Veblen et al. 1989). Since 1948, when the first observation was recorded on Isla Victoria (Havrylenko et al. 1989), and extending over the island and the region with geographically varying intensities, the main present disturbance pattern is mal del ciprés Austrocedrus mortality.
Superimposed on the pattern of disturbance by fires and dieback are the effects of introduced herbivores. In 1916, red deer (Cervus elaphus), axis deer (Axis axis), and fallow deer (Dama dama) were successfully introduced to the island. At present, red deer and fallow deer are extremely abundant, while axis deer is apparently extinct on the island. By 1959, exotic deer densities on the island were estimated to be 40 individuals/km2 (Anziano 1962), and recent estimates indicate densities of 26 individuals/km2 (Relva unpubl.). Average red deer density throughout the present distributional range in Patagonia has been estimated at about 2 individuals/km2 (Flueck et al. 2003); however, these authors also state that in favourable conditions densities may reach 100 deer/km2 (ecotonal habitat) and 40–50 deer/km2 (steppe habitat). Exotic deer have significantly modified the forests on Isla Victoria, reducing cover by palatable species, such as Aristotelia chilensis (Veblen et al. 1989), and delaying the growth of Austrocedrus and Nothofagus dombeyi seedlings and saplings to adult size (Veblen et al. 1989; Relva and Veblen 1998).
Field sampling
Mortality assessments
In each area we used dendroecological techniques (Stokes and Smiley 1968) to reconstruct the timing and duration of tree mortality events. In each area, in fifteen 314 m2 plots we cored the closest live tree to the centre of the plot at ca. 50 cm with increment borers to determine dates of growth release related to mal del ciprés mortality and/or associated windthrow from neighbour trees. Dead standing, wind-snapped, and uprooted trees were sampled by cutting partial cross-sections at the base of each individual to date the year of death. All samples were sanded with successive grades of sandpaper to obtain an optimal view of annual rings. Ring widths in tree cores and cross-sections were measured to the nearest 0.01 mm using a Henson computer-compatible radial increment-measuring device. Disturbance dates were determined on living trees by detecting growth release events. In this study, we define release events as occurring when the tree-ring width of five contiguous years increased more than 150% compared to the preceding 5 years growth (Kitzberger et al. 2000a). The growth release frequencies were quantified in 10-year periods by calculating the number of individuals that underwent growth release in a period relative to total individuals present in that period. Dates of death of dead-standing and downed trees were established using the standard visual skeleton plots method (Stokes and Smiley 1968) in combination with the COFECHA cross-dating program (Holmes 1983). This program statistically analyses the correlation between pieces of undated (floating) tree-ring series and master series dated independently. For cross-dating, Cerro Los Leones (International Tree Ring Data Bank, http://www.ngdc.noaa.gov/paleo/treering.html) was used as the master tree ring chronology.
Vegetation and deer use
In each area we sampled forest structure, understory abundance and composition, and deer use with 15 concentric plots of variable sizes placed systematically every 20 m along three parallel lines that were located in relatively homogeneous areas, each approximately 50 m apart from adjacent lines. Forest structure was sampled in fifteen 314 m2 circular plots, in which we measured diameters of adult trees (>4 cm at breast height) in four categories: living, uprooted dead, standing dead, and snapped dead tree. Understory abundance and composition were surveyed in fifteen 100 m2 circular plots in which we visually estimated cover by individual species of tree saplings (height >10 cm and dbh <4 cm), shrubs, and herbs. In each 100 m2 circular plot, we also counted and measured tree sapling height and assessed seedling abundances (height <10 cm) by counting within four 1 m2 plots randomly distributed throughout the 100 m2 understory plots. We measured the tallest shrub of each species and used a scale according to Allen and McLennan (1983) to assess the degree of browsing on saplings and shrubs. This scale distinguishes: 0, no evidence of browsing; (1) slightly browsed (one or two branches browsed); (2) moderately browsed (more than two branches browsed), and (3) heavily browsed (most branches browsed). Pellet groups were counted using a 10 m2 circular plot placed in each study station. Degree of browsing and pellet group counts were used as an index of animal use (Mayle et al. 1999). The degree of site accessibility to deer was estimated by measuring the maximum height of logs and fallen branches, and by estimating their cover as was done in the understory plots.
Fallen tree-removal experiment
To evaluate the effects of fallen trees on deer–vegetation interactions, we performed a blocked sampling design at control areas (Criollos, Redonda, and Larga) and three nearby (<200 m away) areas from which all downed dead trees had been removed in 1994, 1997, and 1998, respectively (hereafter, removal treatment). There were two different control (non-removal) areas: (1) areas with more than two downed trees (hereafter, non-removal treatment), and (2) naturally open areas between fallen trees (hereafter, non-removal open treatment). Each treatment was sampled in stratified manner using fifteen 20 m2 circular plots. Variables describing forest structure, understory abundance and composition, and animal use were recorded in a similar fashion to those described at the beginning of this section.
Data analyses
We investigated the interaction among forest mortality, deer use, and understory traits through multiple stepwise regression. One set of regression analyses was performed to determine the minimum set of variables related to forest mortality and understory traits that allow us to predict deer use (pellet group counts and degree of browsing as dependent variables). A second regression analysis determined the variables related to forest mortality and deer use that can explain the abundance of palatable and unpalatable shrubs species in the understory. Independent variables related to forest mortality were: (i) history: according to dendroecological data forest stands that were categorized as recent (1, death dates peaking in the 1980s) and old (2, death dates peaking in the 1970s), and (ii) severity: expressed as basal area of live, uprooted, standing dead and snapped trees, and cover of fallen branches. Variables related to understory traits were herb cover, tree sapling cover, and cover of unpalatable and palatable shrubs.
Effects of fallen tree removal on plant community and deer use were evaluated by ANOVA using areas as experimental units and triplets of log removal/log non-removal/non-removal open treatments as blocks. Differences in means between treatments were based on post-hoc tests. In all statistical analyses, counts (numbers of pellet groups) and measures (heights) were log-transformed, and proportions (understory cover) were arcsine-transformed when needed to achieve normality and homoscedasticity.
Results
Timing and severity of tree mortality
Growth release patterns in surviving trees and frequency patterns of death dates suggest differences in timing and severity of mortality occurred within the study area. Larga showed the longest and most uniform history of mortality with death dates and releases starting in the 1950s, peaking in the 1970s, and extending into the 1980s (Fig. 3). Redonda showed evidence of mortality starting mainly in the 1960s and peaking in the 1970s, while at Criollos, mortality started in the 1970s and peaked in the 1980s and 1990s. Similar to Criollos, but with less severity, Pseudotsuga had mortality starting in the 1970s and peaking in the 1980s (Fig. 3). At Criollos, 80% of the dated uprooting occurred in a relatively distinct period during the 1980s and 1990s. By contrast, uprooting during that same period accounted for 40% and 50% of downed trees at Redonda and Larga, respectively, thus suggesting a more gradual process of canopy collapse. During the 1990s, dead trees in massive mortality stands (Criollos, Redonda, and Larga) were mostly uprooted (Fig. 3). Ring width patterns of these uprooted trees indicated the existence of growth release events in a large percentage of trees (50, 75, and 100% at Larga, Criollos, and Redonda, respectively). This fact suggested that wind-induced uprooting occurred after canopy opening owing to mortality of dead standing trees and/or uprooting of neighbouring trees.

Frequency of tree death dates by cause (wide bar) and frequency of live tree releases (narrow bar) at the study sites
Criollos, Redonda, and Larga were on average the stands affected the longest and most severely by mal del ciprés mortality and subsequent windthrow (Table 1). Around 25% of the basal area consisted of live trees, whereas 50–60% of the basal area consisted of downed, uprooted trees. By contrast, Pseudotsuga, which was the youngest stand (Table 1), suffered lower overall levels of mortality and subsequent tree fall with ca. 45% of tree basal area alive and ca. 35% of the basal area on the ground. Percentages of wind-snapped and standing dead trees were relatively uniform among stands (Table 1).
Predictors of deer use and shrub composition
The multiple regression analyses showed that deer use was positively related to the history of stand mortality (stands with older mortality are used more heavily) and negatively related to branch cover. Thirty-five percent of the variance in the number of deer pellets was explained by the history of stand mortality (+, P < 0.01) and fallen branch cover (−, P < 0.01) (model: F = 6.44; df = 4,48; P = 0.0031). Similarly, 32% of the variance in the degree of browsing on plants was explained by the history of stand mortality (+, P < 0.05) and fallen branch cover (−, P < 0.05) (model: F = 2.99; df = 7,45; P = 0.011). By contrast, no single vegetation variable significantly explained deer use.
Composition of understory vegetation was also explained mostly by history and severity of stand mortality. Fifty percent of the variance in cover of unpalatable shrub species was positively related to history of stand mortality (stands with older mortality have higher cover of unpalatable shrubs, P < 0.001), while the degree of browsing was negatively related to cover of unpalatable shrubs (P < 0.05) (model: F = 9.38; df = 5, 47; P = 0.001). Cover of palatable species was related only to basal area of uprooted trees, a measure of mortality severity (P < 0.05) (model: F = 4.46; df = 3, 49; P = 0.00756), explaining 21% of the variance in palatable species cover.
Effects of fallen trees on deer use and vegetation
As expected, uprooted basal area (F = 112.8, df = 2, P < 0.001, Table 2) and branch cover on the ground (F = 37.16, df = 2, P = 0.001) in the fallen tree removal treatment were lower than those found in the non-removal treatment. In the treatment in which fallen trees had been removed and in the naturally open treatment, deer pellet number (F = 75.1, df = 2, P < 0.001) and browsing (F = 23, df = 2, P = 0.002, Table 2) were higher than in the adjacent treatment in which fallen trees had not been removed. Total shrub cover was similar among removal and non-removal treatments (F = 3.99, df = 2, P = 0.079). However, the proportion of palatable shrub species—such as Aristotelia chilensis, Ribes magellanicum, Maytenus boaria—was significantly higher in the non-removal treatment compared with the removal treatment and the naturally open treatment (F = 41.53, df = 2, P = 0.001). Conversely, cover by unpalatable shrubs—such as Berberis spp.—was 15.8% and 12.3% higher in the removal treatment and naturally open treatment, respectively, than in non-removal treatment (F = 38.75, df = 2, P = 0.001, see Appendix). No significant differences were found in total herb cover among the three treatments (F = 1.73, df = 2, P = 0.25, Table 2).
Discussion
Timing and severity of tree mortality
Austrocedrus areas with moderate mortality (ca. 65% of basal area dead) are relatively open, young, and accessible forest with most trees alive or standing but dead. In contrast, where mortality exceeds 75% of basal area, many trees lie on the ground forming an inaccessible tangled mass of logs and branches several meters high. Mortality levels in this study are similar to those found by Loguercio and Rajchenberg (2004) but higher than those found by La Manna et al. (2006) for forests with similar stand structure in southwestern areas of Río Negro and in the nearby province of Chubut.
Two temporal factors are important in the interaction between mortality and herbivores that may affect plant communities: (1) the timing of canopy opening (i.e., increase in light levels to understory plants) and (2) the timing of canopy collapse (i.e., decreasing accessibility to herbivores). These stages do not necessarily coincide. Dendroecological techniques allowed us to differentiate both processes. In our system, most trees were attacked by root fungi, lost foliage, and remained standing until root rot made the trunk unstable and the tree fell. This was evidenced in ring growth patterns of downed trees by a strong suppression before and at the time of death. Additional unattacked trees fell because the lack of surrounding canopy trees made them susceptible to wind-throw. This was evidenced in downed trees by strong radial growth release (suggesting that trees were not infected) before sudden death by snapping or uprooting. In both cases, canopy opening may not result in understory blocking for several years or even one or two decades. This time lag between canopy opening and understory physical blocking may have an impact on understory composition. During early phases of the decline process, the understory receives light but there is also substantial herbivore pressure. Therefore shade-intolerant plants that are resistant to herbivores or are dispersed by them may benefit. In our system, such as species may be Uncinia sp., which dominated recently dead forest, is light-demanding, and is dispersed in deer fur. The initial density of the stand may have been important determinants of how fast the canopy collapsed after mortality began. In our study, in all dense areas (Criollos, Larga, and Redonda) uprooting has been the main cause of mortality process for the past three decades. The death dates in our study are similar to those registered by Cali (1996), who worked in two mainland Austrocedrus stands close to our study sites.
Interactive effects of forest mortality and deer use on plant communities
Our results indicate that fallen logs with a high density of branches strongly limited deer accessibility to certain microsites and created natural exclosures and safe sites for palatable plant establishment and growth. Pulido et al. (2000) found a similar relationship between presence of a native camelid, Lama guanicoe (guanaco), and slash in a managed Nothofagus pumilio forest in Tierra del Fuego (southern Argentina). Rebertus et al. (1997) found that browsing by guanaco was negatively correlated to the blowdown area of N. pumilio forest in Tierra del Fuego. In blowdown areas above 5 hectares, guanaco browsing was restricted to the periphery. Similarly, Cavieres and Fajardo (2005) found in old-growth stands of N. pumilio that guanaco damage was higher in small gaps than in the bigger ones. On the other hand, postfire coarse woody debris has been found to provide Populus tremuloides refugia from red deer browsing in Yellowstone National Park (Ripple and Larsen 2001). On the contrary, Bergquist and Örlander (1998) found that Picea abies browsed by moose did not vary in sites with different amounts of slash on the forest floor. Similarly, Kupferschmid and Bugmann (2005) found that fallen trees do not constitute a barrier to chamois (Rupicapra rupicapra) browsing Picea abies saplings. According to Thomas et al. (1979), a depth of dead and fallen material higher than 0.6 m substantially limits deer use of the area, and when the depth is high enough to make deer jump, the energetic cost of locomotion increases dramatically (Hanley et al. 1989; Nyberg 1990). Another complementary explanation for deer to avoid areas with deep slash is that they would not be able to escape easily if a predator does attack (White et al. 2003).
In our study, the negative relationship between the amount of fallen logs and the deer use was clearly manifested when slash was removed. The number of deer pellet groups found where slash had been removed was six times the number found in control areas. As a result of this heavier use, after only 4 years of the treatment, understory composition changed dramatically toward more unpalatable and browse-resistant species in the slash-removal treatments.
The positive relationship between deer use and time since peak mortality suggests that with time, fallen trees lose decomposing branches, and accessibility increases. In the early stages, shrubs would be not abundant except for Aristotelia chilensis, a shade-intolerant, tall shrub (Muñoz and González 2006) that is highly palatable and consumed by deer (Anziano 1962; Veblen et al. 1989; Relva and Veblen 1998; Relva and Caldiz 1998). In areas with recent or severe mortality, A. chilensis was observed growing between logs and fallen branches. This spatially aggregated distribution in herbivore-free refuges (i.e. safe sites where individuals grow and reproduce successfully, far from the browsing range of the herbivores) located in grazing areas was also observed by Vázquez (2002a), who also found that this type of distribution influenced the mechanisms of pollination of this species. Positive association between certain species of plants with coarse debris has been noted in other forest systems in which windstorms were generally predominant and produced great amounts of dead material on the forest floor (Allan et al. 1997; Peterson and Pickett 2000; de Chantal and Ganström 2007). However, the strong positive relationship between A. chilensis and fallen branches could additionally be a response to improved recruitment conditions, as shown in other species (Schreiner et al. 1996). In areas with the oldest mortality (Redonda and Larga) and in microsites from which logs had been removed, deer use increased, and shrub composition changed toward less palatable species or browse-resistant ones such as Berberis spp. Both B. buxifolia and B. darwinii, which are common in Austrocedrus forests, are dominant in intensely grazed areas (Rebertus et al. 1997; Vázquez 2002b; Gallopin et al. 2005). Berberis spp. and other spiny shrubs may act as nurse plants of other species, by physically protecting more palatable plants from herbivores (De Pietri 1992) and/or improving abiotic conditions to facilitate establishment and growth of tree seedlings (Kitzberger et al. 2000b). In our study site, we have found no saplings of Austrocedrus in recently and severely disturbed forest. This could be because of the high cover of light-demanding herbs, Uncinia sp. and Digitalis purpurea, in early post-disturbance stages that could be negatively affecting tree seedling recruitment or due to low seed production by overmature trees. By contrast, in areas with less severe mortality, Austrocedrus seedlings and saplings are a dominant component of the understory (see Appendix). Because Austrocedrus is a shade-intolerant species, the canopy opening produced by less severe mortality probably explains this abundant tree regeneration despite heavy use of canopy gaps by deer (Veblen et al. 1989; Relva and Veblen 1998).
The spatially and temporally heterogeneous nature of forest mortality interacting with large herbivores may shape complex mosaics of vegetation. Prediction of plant community composition and structure should move forward from approaches that emphasize disturbances modifying abiotic resources for plant regeneration or plant–animal interactions toward spatially explicit approaches that integrate plant performance and animal behaviour within the context of a dynamic forest landscape.
This study underpins the importance of maintaining snags and large woody debris for the role in providing safe sites for tree and understory regeneration, a management policy that should also extend to disturbed landscapes where salvage logging is a routine procedure.
References
Allan C, Stewart G, Allen R (1997) Long term influences of deer browsing on forest health and conservation values of the Kaweka Range. Final Report Lincoln University and WWF, Canterbury
Allen RB, McLennan MJ (1983) Indigenous forest survey manual: two inventory methods. New Zealand For Serv FRI Bulletin no. 48
Anziano AF (1962) Acción de los animales sobre la flora. Anales de Parques Nacionales, Administración de Parques Nacionales. Tomo IX:107–112
Barros VR, Cordon VH, Moyano C, Mendez R, Forquera JC, Pizzio O (1988) Carta de precipitación de la zona oeste de las provincias de Río Negro y Neuquén. Primera contribución (1972–1981). Facultad de Ciencias Agrarias. Universidad Nacional del Comahue, Cinco Saltos
Barrios Garcia Moar MN (2005) Patrones de daño, uso actual y tendencia histórica de la abundancia de ciervos introducidos en bosques nativos de Isla Victoria, Patagonia, Argentina. Licenciate Dissertation. Universidad Nacional del Comahue, Bariloche, Argentina
Bergquist J, Örlander G (1998) Browsing damage by roe deer on Norway spruce seedlings planted on clearcuts of different ages. 1. Effect of slash removal, vegetation development, and roe deer density. For Ecol Manage 105:283–293
Cali SG (1996) Austrocedrus: estudio de los anillos de crecimiento y su relación con la dinámica del “mal del ciprés” en el Parque Nacional Nahuel Huapi, Argentina. Licenciate Dissertation. Universidad Nacional del Comahue, Bariloche, Argentina
Cavieres LA, Fajardo A (2005) Browsing by guanaco (Lama guanicoe) on Nothofagus pumilio forest gaps in Tierra del Fuego, Chile. For Ecol Manage 204:237–248
de Chantal M, Ganström A (2007) Aggregations of dead wood after wildfire act as browsing refugia for seedlings of Populus tremula and Salix caprea. For Ecol Manage 250(1):3–8
De Pietri DE (1992) Alien shrub in a National Park: can they help in the recovery of natural degraded forest? Biol Conserv 62:27–130. doi:10.1016/0006-3207(92)90933-E
Ezcurra C, Brion C (2005) Plantas del Nahuel Huapi. Catálogo de la Flora Vascular del Parque Nacional Nahuel Huapi, Argentina
Filip GM, Rosso PH (1999) Cypress mortality (mal del ciprés) in the Patagonia Andes: comparison with similar forest disease and declines in North America. Eur J Forest Pathol 29:89–96. doi:10.1046/j.1439-0329.1999.00133.x
Flueck WT, Smith-Flueck JA, Naumann CM (2003) The current distribution of red deer (Cervus elaphus) in southern Latin America. Z Jagdwiss 49:112–119. doi:10.1007/BF02190451
Gallopin GC, Martín CE, Mermoz MA (2005) Impacto de la ganadería en la cuenca del río Manso Superior. Parte I: Bosque de Ñire con Laura. Anales de Parques Nacionales, Administración de Parques Nacionales. Tomo XVII:9–36
Greslebin A, Hansen E (2006) Novedades sobre el mal del ciprés. Patagonia For 12(1):11–14
Hanley TAR, Robbins CT, Spalinger DE (1989) Forest habitats and the nutritional ecology of Sitka black-tailed deer: a research synthesis with implications for forest management. USDA For Serv Res Pap PNW-GTR-230
Havrylenko M, Rosso PH, Fontenla S (1989) Austrocedrus chilensis: contribución al estudio de su mortalidad en Argentina. Bosque 10(1):29–36
Hobbs NT (1996) Modification of ecosystems by ungulates. J Wildl Manage 60(4):695–713. doi:10.2307/3802368
Holmes R (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-ring Bull 43:69–75
Jefferies RL, Klein D, Shaver GR (1994) Vertebrate herbivores and northern plant communities: reciprocal influences and responses. Oikos 71:193–206. doi:10.2307/3546267
Kitzberger T, Veblen T, Villalba R (2000a) Métodos dendroecológicos y sus aplicaciones en estudios de dinámica de bosques templados de Sudamérica. In: Roig FA (ed) Dendrocronología en América Latina. EDIUNC, Universidad de Cuyo, Mendoza, Argentina, pp 17–78
Kitzberger T, Steinaker D, Veblen T (2000b) Effects of climatic variability on facilitation of tree establishment in northern Patagonia. Ecology 81(7):1914–1924
Koutché V (1942) Estación Forestal de Puerto Anchorena, Isla Victoria; su organización y trabajos. Ministerio de Agricultura, Dirección de Parques Nacionales, Buenos Aires, Argentina
Kupferschmid A, Bugmann H (2005) Effect of microsites, logs and ungulate browsing on Picea abies regeneration in a mountain forest. For Ecol Manage 205:251–265
La Manna L, Rajchenberg M (2004) The decline of Austrocedrus chilensis in Patagonia, Argentina: soil features as predisposing factors. For Ecol Manage 190:345–357
La Manna L, Bava J, Collantes M, Rajchenberg M (2006) Características estructurales de los bosques de Austrocedrus chilensis afectados por “mal del ciprés” en Patagonia, Argentina. Bosque 27(2):135–145
La Manna L, Matteucci SD, Kitzberger T (2008) Abiotic factors related to the incidence of the Austrocedrus chilensis disease syndrome at a landscape scale. For Ecol Manage 256:1087–1095
Loguercio G, Rajchenberg M (2004) El “mal del ciprés” y la silvicultura del ciprés de la cordillera. In: Arturi MF, Frangi JL, Goya JF (eds) Ecología y Manejo de los Bosques de Argentina. EDULP, Universidad Nacional de La Plata, La Plata, Argentina
Mayle BA, Peace AJ, Gill RMA (1999) How many deer? A field guide to estimating deer population size. Field Book 18. Forestry Commission, Edinburgh
Muñoz A, González M (2006) Aristotelia chilensis (Mol) Stuntz. In: Donoso C (ed) Las especies arbóreas de los bosques templados de Chile y Argentina. Autoecología, Cuneo Ediciones, pp 166–172
Nyberg JB (1990) Interaction of timber management with deer and elk. In: Nyberg JB, Janz DW (eds) Deer and elk habitat as in coastal forests of Southern British Columbia. BC Ministry of Forests and Ministry of Environment, Special Report 5, pp 99–131
Peterson CJ, Pickett STA (2000) Patch type influences on regeneration in a western Pennsylvania, USA, catastrophic windthrow. Oikos 90:489–500. doi:10.1034/j.1600-0706.2000.900307.x
Pickett STA, White PS (1985) The ecology of natural disturbance and patch dynamics. Academic Press, Orlando
Pulido FJ, Díaz B, Martínez Pastur G (2000) Incidencia del ramoneo del guanaco (Lama guanicoe Müller) sobre la regeneración temprana en bosques de lenga (Nothofagus pumilio (Poepp et Endl) Krasser) de Tierra del Fuego, Argentina. Investigación Agraria. Sistemas Recursos Forestales 9(2):381–394
Rebertus AJ, Kitzberger T, Veblen T, Roovers LM (1997) Blowdown history and landscape patterns in the Andes of Tierra del Fuego, Argentina. Ecology 78:678–692
Relva MA, Caldiz M (1998) Composición estacional de la dieta de ciervos exóticos en Isla Victoria, P.N. Nahuel Huapi, Argentina. Gayana (Zool) 62(2):101–108
Relva MA, Veblen T (1998) Impacts of introduced large herbivores on Austrocedrus chilensis forests in northern Patagonia, Argentina. For Ecol Manage 108:27–40
Ripple WJ, Larsen EJ (2001) The role of postfire coarse woody debris in Aspen regeneration. West J Appl For 16(2):61–64
Russell FL, Zippin DB, Fowler NL (2001) Effects of White-tailed deer (Odocoileus virginianus) on plants, plant populations and communities: a review. Am Midl Nat 146:1–26. doi:10.1674/0003-0031(2001)146[0001:EOWTDO]2.0.CO;2
Schreiner EG, Krueger KA, Happe PJ, Houston DB (1996) Understory patch dynamics and ungulate herbivory in old-growth forest of Olympic National Park, Washington. Can J Res 26:255–265. doi:10.1139/x26-029
Simberloff D, Relva MA, Nuñez M (2003) Introduced species and management of a Nothofagus/Austrocedrus forest. Environ Manage 31:263–275. doi:10.1007/s00267-002-2794-4
Stokes MA, Smiley TL (1968) An introduction to tree-ring dating. The University of Chicago Press, Chicago
Stuth JW (1991) Foraging behaviour. In: Heitschmidt RK, Stuth JW (eds) Grazing management. An ecological perspective. Timber Press, Oregon, pp 65–83
Thomas JW, Black H, Scherzinger RJ, Pedersen RJ (1979) Deer and Elk. In: Thomas JW (ed) Wildlife habitats in managed forest. The Blue Mountains of Oregon and Washington. USDA For Serv Agriculture Handbook 553, pp 104–147
Vázquez D (2002a) Interaction among introduced ungulate, plants and pollinators: a field study in the temperate forest of Southern Andes. PhD Dissertation, University of Tennessee, Knoxville, Usa
Vázquez D (2002b) Multiple effects of introduced mammalian herbivores in a temperate forest. Biol Invasions 4:175–191. doi:10.1023/A:1020522923905
Veblen TT, Lorenz DC (1987) Post-fire stand development of Austrocedrus -Nothofagus forest in northern Patagonia. Vegetatio 71:113–126
Veblen T, Mermoz M, Martín C, Ramilo E (1989) Effects of exotic deer on forest regeneration and composition in northern Patagonia. J Appl Ecol 26:711–724. doi:10.2307/2404095
White CA, Feller MC, Bayley S (2003) Predation risk and the functional response of elk-aspen herbivory. For Ecol Manage 181:77–97
Wisdom MJ, Vavra M, Boyd JM, Hemstrom MA, Ager AA, Johnson BK (2006) Understanding ungulate herbivory-episodic disturbances effects on vegetation dynamics: knowledge gaps and management needs. Wildl Soc Bull 34(2):283–292. doi:10.2193/0091-7648(2006)34[283:UUHDEO]2.0.CO;2
Acknowledgments
We wish to thank Diego Vazquez for valuable comments on the manuscript, park rangers of Isla Victoria (Damián Mujica, Lidia Serantes, Domingo Nuñez, and Carina Pedrozo) for helping us in many ways. Delegación Técnica Regional and Intendencia del Parque Nacional Nahuel Huapi assisted us with working permits, and Cau Cau and Mares Sur with transportation. We are especially grateful to Juan Gowda for helping on cross-section tree extractions, and Eduardo Zattara for his field assistance. Daniel Simberloff revised several versions of this manuscript improving the language and clarity. This research was supported by a postdoctoral fellowship to M.A.R from Consejo Nacional de Ciencia y Técnica of Argentina CONICET and by funds from Universidad Nacional del Comahue. Foundation Linnaeus-Palme funded C.L.W scholarship.
Author information
Authors and Affiliations
Corresponding author
Appendix
Appendix
Mean cover (%) and standard error of vascular species recorded in fifteen 100 m2 plots in the study areas
Area | ||||
|---|---|---|---|---|
Criollos | Pseudotsuga | Larga | Redonda | |
Tree species | ||||
Austrocedrus chilensis | 2.83 (1.31) | |||
Lomatia hirsuta | 0.68 (0.29) | 1.69 (1.06) | ||
Luma apiculata | 1.5 (1) | 1.69 (0.99) | 1.87 (0.99) | 16.79 (5.66) |
Maytenus boaria | 0.01 (0.01) | 0.01 (0.01) | 0.01 (0.01) | |
Nothofagus dombeyi | 0.17 (0.17) | 1.69 (0.99) | 1.08 (1.07) | |
Pseudotsuga menziesii a | 1.34 (0.33) | |||
Shrub species | ||||
Aristotelia chilensis | 6.34 (2.68) | 0.35 (0.23) | 3.21 (2.47) | 4.14 (1.60) |
Azara lanceolata | 0.01 (0.01) | 0.01 (0.01) | ||
Berberis darwinii | 2.51 (0.94) | 3.2 (1.27) | 39.5 (7.47) | 20.36 (5.73) |
Budleja globosa | 0.18 (0.17) | 0.01 (0.01) | 0.01 (0.01) | |
Colletia hystrix | 0.18 (0.17) | 0.02 (0.01) | ||
Gaultheria spp. | 2.67 (2.49) | 0.54 (0.28) | ||
Maytenus chubutensis | 0.03 (0.01) | 0.19 (0.18) | ||
Ribes magellanicum | 0.36 (0.22) | 0.21 (0.18) | 0.17 (0.17) | 0.57 (0.28) |
Rosa rubiginosa a | 0.01 (0.01) | |||
Schinus patagonicus | 0.01 (0.01) | 4.36 (1.45) | 5.56 (1.68) | |
Herb species | ||||
Acaena ovalifolia | 0.01 (0.01) | 0.01 (0.01) | ||
Adiantum chilense | 0.01 (0.01) | |||
Alstroemeria aurea | 0.01 (0.01) | 0.01 (0.01) | ||
Blechnum spp. | 0.17 (0.17) | |||
Carex spp. | 0.01 (0.01) | 0.01 (0.01) | ||
Cynanchum diemii | 0.53 (0.26) | 3.67 (1.22) | 0.54 (0.28) | |
Cynoglossum creticum a | 2.51 (1.33) | 1.38 (1.00) | 15.51 (4.32) | 1.27 (1.07) |
Digitalis purpurea a | 30.17 (6.76) | 13.01 (3.67) | 7.16 (2.78) | |
Galium aparine a | 0.34 (0.23) | 0.21 (0.18) | ||
Vicia nigricans | 0.02 (0.01) | 0.01 (0.01) | 0.01 (0.01) | |
Mutisia spp. | 1 (1) | 0.045 (0.01) | 2.87 (2.67) | |
Rumex acetosella a | 0.02 (0.01) | 0.01 (0.01) | 0.2 (0.18) | |
Rumohra adiantiformis | 0.01 (0.01) | |||
Uncinia spp. | 67.83 (6.8) | 1.53 (1.00) | 3.02 (1.29) | 33.93 (6.74) |
Grasses | 0.01 (0.01) | |||
Rights and permissions
About this article
Cite this article
Relva, M.A., Westerholm, C.L. & Kitzberger, T. Effects of introduced ungulates on forest understory communities in northern Patagonia are modified by timing and severity of stand mortality. Plant Ecol 201, 11–22 (2009). https://doi.org/10.1007/s11258-008-9528-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11258-008-9528-5