
Abstract
The conflict resulting from the expansion of human activities into natural habitats affects the structure and functioning of ecosystems. Similarly, the anthropogenic redistribution of many species all over the world affects the composition of biological communities, possibly altering their capacity to sustain key ecological functions, such as seed dispersal. Urban parks are extreme examples of such novel ecosystems resulting from the anthropogenic redistribution of species in a new ecological framework. Here, we describe the avian seed dispersal network in an urban recreational woodland in central Portugal (c.79 ha). Four quantitative seed dispersal networks were assembled by identifying intact seeds in the droppings of mist-netted birds throughout the year. Overall, 1,244 seeds were identified, representing 33 links between 15 plant species and 11 bird species. Most birds dispersed alien seeds, but these represented a small proportion of the overall network (20 % of the seeds and 13 % of the droppings). Blackcap (Sylvia atricapilla) was the main disperser of native and alien seeds in all seasons, particularly those of the invasive Phytolacca americana. Fleshy fruits were more abundant in summer, but were more consumed in winter, presumably when other foods were difficult to find. Our study suggests that even in a highly managed urban park, seed dispersal networks can be temporally complex and variable and that a network approach can be an important monitoring tool to detect the status of crucial ecosystem functions in rapidly changing habitats such as urban parks.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Natural ecosystems face multiple anthropogenic threats such as agriculture intensification, urbanization and the impact of alien invasive species (Barnosky et al. 2012; Hooper et al. 2012). Urbanization, in particular, had significant effects on natural environments because it severely affects the ecosystem structure, leading to habitat loss, degradation and homogenization (McKinney and Lockwood 1999; Alberti et al. 2003; Alberti 2005). Even when pockets of natural habitats remain in the urban tissue, fragmentation is a serious problem as fragmented areas tend to be more simple and more sensitive to impacts such as biological invasions (Vilà and Ibáñez 2011) and local extinction (Kuussaari et al. 2009), thereby altering the ecosystems’ capacity to perform key ecological functions (Kremen and Hall 2005). In addition, the mobility that characterizes the human society resulted in the redistributing of many species all over the world, leading some species to thrive in places that would not have been naturally colonized (Vitousek et al. 1996). The existence of these two factors—novel species mixes and new environmental conditions—led to the development of a variety of novel ecosystems (sensu: Hobbs et al. 2006), of which urban forests and parks are representative.
Novel ecosystems contain “new combinations of species that arise through human action, environmental change and the impacts of the deliberate and unintentional introduction of species from other parts of the world” (Hobbs et al. 2006). Since natural ecosystems are becoming increasingly impacted by humans and less capable to deliver the ecosystems services upon which we all rely, novel ecosystems such urban parks are becoming more important as reservoirs of native biodiversity (Marzluff et al. 2001) and also by assuming some of those functions, as they represent the remaining near natural habitat (Vitousek et al. 1996). However, there is still very little information regarding the dynamics and management options of these new anthropogenic habitats as the planet becomes more altered by human activities (Hobbs et al. 2009; Seastedt et al. 2008). This information is needed as traditional notions of ecosystem management might prove inadequate at the present, for example, when alien species are impossible to eradicate or when they undertake important roles in the functioning of these ecosystems (Hobbs et al. 2009; Ceia et al. 2011).
Seed dispersal is an important ecosystem function that links successive generations of plants and influences the dynamics of vegetation (Fuentes 2000; Howe and Miriti 2004). It affects vegetation structure and, ultimately, the long-term survival of plant species (Nathan and Muller-Landau 2000; Wang and Smith 2002). By acting as vectors for the dispersal of seeds, frugivorous animals have a crucial role in this process, particularly birds, which are the most important seed dispersers in many temperate ecosystems (Herrera 1995; Sekercioglu 2006; Whelan et al. 2008). Most of the interactions between bird dispersers and plant species usually comprise a generalized dispersal system (Howe and Smallwood 1982). However, seed dispersal has been negatively affected by multiple factors in many regions of the planet (Cordeiro and Howe 2003; Heleno et al. 2013a), which can negatively affect plant recruitment with cascading consequences for the biological communities (Moran et al. 2009; Cordeiro and Howe 2003).
In this study, we evaluate the avian seed dispersal network in an urban recreational woodland (c.79 ha), consisting of a mixture of broad-leaved native and alien tree species in central Portugal. In particular, we aimed to answer the following questions: (1) What is the predominance of alien fruits in the forest? (2) Which are the main seed dispersers? (3) What disperses what, how much and when? (characterization of the seed dispersal network); and (4) What is the relative importance of native and alien seeds for frugivorous birds and vice versa?
Methods
Study site
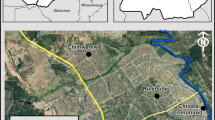
The study was carried out at Mata Nacional do Choupal (hereafter Choupal) (40°13′24″N, 8°27′53″W), a mixed deciduous suburban woodland in central Portugal (Fig. 1). Choupal is a natural reserve established in 1791 near the city of Coimbra in the lower Mondego basin. The climate is predominantly Mediterranean with an Atlantic influence—characterized by warm and dry summers and mild rainy winters (Rebelo 2003; Norte et al. 2010). The mean annual temperature is 15 °C, and the mean annual precipitation is 975 mm, most of it occurring from October to February (data from Geophysical Institute of the University of Coimbra collected from 1971 to 2000). This woodland was originally created to stabilize the banks of the Mondego River, and several forest species were planted along the river bed and marginal land (Silva et al. 2011). The vegetation is composed mostly of the native deciduous trees, mainly black poplar (Populus nigra), plane tree (Platanus sp.), common sallow (Salix atrocinerea), white willow (Salix alba) (Norte et al. 2010) and a rich understory of small native trees and shrubs such as elder (Sambucus nigra), bay laurel (Laurus nobilis) and blackberry (Rubus ulmifolius). Some alien plants of global concern are also present in the forest, particularly silver wattle (Acacia dealbata), Australian blackwood (Acacia melanoxylon), the tree of heaven (Ailanthus altíssima) and American pokeweed (Phytolacca americana). In addition to high plant richness, this suburban park maintains a relatively high animal richness, particularly in native bird species, including one of the largest colonies of Black Kite (Milvus migrans) in Europe and some species associated with aquatic habitats. For the present study, it is important to refer the existence of about 15 bird families of song birds as many of them consume fruits and are potential seed dispersers. The most common passerine species include the families Sylvidae: Blackcap (Sylvia atricapilla), Garden Warbler (Sylvia borin) and Whitethroat (Sylvia communis); Turdidae: Blackbird (Turdus merula), Song Thrush (Turdus philomelos) and Robin (Erithacus rubecula); and Fringillidae: Chaffinch (Fringilla coelebs) and Greenfinch (Chloris chloris), all of these being native to the study area. The woodland is used mainly for recreation, leisure and sports, receiving approx. 200,000 visitors per year.

Geographic location of the study area (Choupal) in central Portugal
Fruit and bird abundance and building the seed dispersal networks
We have built seasonal, quantitative seed dispersal networks for Choupal. The study was divided in three periods between July 2011 and February 2012, which cover the complete fructification period for the study area (authors unpublished data): summer (8 July–30 September), autumn (7 October–15 December) and winter (21 December–23 February). To estimate the seasonal abundance of fleshy fruits, ten linear 50 m transects were randomly defined inside the woodland and performed weekly during the duration of the study. In each transect fruit, production was sampled by counting all ripe standing fruits lying within 1 m of the transect line.
Birds were captured during 28 mist-netting sessions performed weakly during the same period, on days of favourable weather, totalizing 140 trapping hours. Sessions were conducted in a random selection of sites (one per session) spread over the entire woodland. In each session, six 12-m-long ecotone mist nets were opened at sunrise and checked every half hour during five consecutive hours. Trapped birds were kept in individual clean cotton bags for up to 20 min to defecate. All entire seeds found in the droppings were identified under a dissecting microscope by comparison with a reference collection of seeds and fruits collected from the study area.
For network construction, interaction frequency was quantified as the number of droppings from each bird species containing intact seeds of each plant species. This measure includes an assessment of relative bird abundance based on mist-netting captures. Comparisons involving fruit and bird abundances were performed with categorical analyses (likelihood ratio tests) conducted in R (R Development Core Team 2010).
Network structure
We evaluated seasonal changes in the structure of the seed dispersal networks using nine commonly used descriptors: (1) number of bird species (a); (2) number of plant species (p); (3) network size (a × p); (4) number of links (l); (5) connectance—the proportion of links that are realized among the pool of all possible links; (6) mean number of disperser species shared between plants; (7) mean number of dispersers per plant; (8) mean number of plants per disperser; and (9) interaction evenness—the Shannon’s evenness of interaction frequencies (Bersier et al. 2002). Quantitative seed dispersal networks were analysed with package Bipartite 1.16 (Dormann et al. 2008) for R (R Development Core Team 2010) and visualized using specific code written in Mathematica 6.0 (Wolfram Research 1999).
Results
Overall, 22,797 fruits of eight plant species were counted on the linear transects, the native: S. nigra, R. ulmifolius, Smilax aspera, Solanum nigrum, C. australis, L. nobilis and Hedera helix and the alien: P. americana. Fruits were more abundant in summer (15,718 fruits) than in autumn (3,549 fruits) and winter (3,530 fruits) (G test summer/autumn/winter, G = 12029.9; df = 2; P < 0.001; G test autumn/winter G = 0.1; df = 1; P = 0.821). Overall, alien fruits were significantly more abundant than native fruits (G test, G = 157.87; df = 1; P < 0.001), although this tendency was mainly driven by high alien fruit abundance in the summer and autumn, while in winter only native fruits were counted along the sampling transects (all seasonal differences were statistically different; P < 0.001). The majority of fruits were of the alien species P. americana, representing 54 % of all fruit production. The remaining fruits (10,459; 46 %) were from native species, among which the most abundant were S. nigra, H. helix and C. australis.
A total of 1,137 mist-netted birds belonging to 35 species were captured (“Appendix 1”). Overall, 464 droppings produced by 30 bird species were collected, of which 209 (45 %), produced by 11 native bird species, contained intact seeds. A total of 1,244 intact seeds from 15 plant species were retrieved from these droppings, revealing a total of 33 links to dispersers (Fig. 2a). The 15 seed species included eight natives and three aliens: P. americana, A. melanoxylon and Apollonias barbujana. Four seed morphotypes were not possible to identify to the species level, and thus, their origin remained unknown. Overall, 975 native seeds (81.3 %) and 224 alien seeds (18.7 %) were recovered from the droppings.

Visualization of the seed dispersal network of Choupal, a suburban woodland in central Portugal. The overall network (top) results from the fusion of the three seasonal networks for summer, autumn and winter (bottom). Species are represented by boxes, plants on the lower row and birds in the upper row in each network. The width of the boxes at the plant level represent fruit abundance, whereas the width of the boxes representing birds and the interactions are proportional to the presence of intact seeds retrieved from birds droppings. Native seeds are shown in black and alien seeds in light grey. Dotted lines indicate seeds found in the droppings that were not detected in the fruit abundance transect. The two open boxes on the left of the top network represent the scale used for each level
Blackcap was found to be the main seed disperser in all seasons, dispersing 937 seeds of 11 plant species (Fig. 2). While Blackcap is overall, the most abundant species (29 % of all captures), its importance as seed disperser is highly disproportionate to its abundance, being responsible for 74 % of all found seeds and 83 % of the droppings containing seeds (“Appendix 2”). Song Thrush and Blackbird followed, contributing with 9.4 % (117 seeds; 5 species) and 6.3 % (78 seeds; 5 species), respectively. Despite the greater number of captured Blackbirds (81 captures) in comparison with Song Thrush (19 captures) and proportional number of droppings analysed (26 and 10, respectively), Song Thrushes dispersed a significantly greater number of intact seeds (G test: G = 47.4; df = 1; P < 0.001).
In summer, the Blackcap and the Song Thrush were the main seed dispersers. Blackcap had a more diverse diet (6 plant species) dominated by the common alien species P. americana (129 seeds representing 43.4 % of the diet) and the native R. ulmifolius and S. nigra (Fig. 2b). In autumn, the fruits of P. americana started to decrease and C. australis became the most common fruit species dispersed by Blackcaps, followed by S. nigra (Fig. 2c). Finally, in winter, Blackcaps maintained their importance as main dispersers both in diversity and abundance, dispersing the seeds of six plant species, in which H. helix prevailed (208 seeds representing 70 % of the diet) and including the seeds of the highly invasive A. melanoxylon (6 seeds). Blackbird dispersed three plant species, with a particular reference for A. barbujana that is endemic to the Macaronesian islands and was planted in the study area (Fig. 2d). Overall, although fruit availability was higher in summer, fruit consumption by birds was greater in winter (available [consumed] fruits in summer = 15,718[57], in autumn = 3,549[58], and in winter 3,530[94]; G = 160.1; df = 2; P < 0.001).
The overall seed dispersal network indicates that local seed dispersers have integrated alien fruits into their diets, mainly the common P. americana, which was dispersed by five species: Robin, Pied Flycatcher (Ficedula hypoleuca), Blackcap, Garden Warbler and Song Thrush. From these five bird species, the largest interaction was with the Blackcap, which dispersed 85 % of the seeds of P. americana.
Network connectance was higher in autumn and winter than in summer (Table 1). Interaction evenness, on the contrary, was higher in summer, as was the mean number of plants per disperser.
Discussion
Despite the large representation of the alien P. americana, among all available fruits (54 %), the proportion of alien seeds in the network was relatively low (20 % of the seeds retrieved and present in 13 % of the droppings). However, this study shows that native bird species will readily ingest and disperse alien species present in novel ecosystems.
Sampling limitations
The quantification of seed dispersal interactions was based in the analysis of droppings of mist-netted birds. This measure integrates an estimation of relative bird abundance obtained by bird captures, which might be subject to some bias, namely the under sampling of: (1) large, (2) low mobility and (3) canopy specialist birds (Pagen et al. 2002; Wang and Finch 2002; Estades et al. 2006). While estimations of bird diet and seed dispersal interactions can also be obtained via direct observations of feeding records and seed fate with binoculars, this method is largely subject to the same limitations, plus observer-related bias and biases emerging from different detectability rates through the year (e.g. reduced vocalization outside the breeding season) hindering cross-season comparisons (Ralph et al. 1993; Bibby et al. 2000). Given that this study focuses on forestry species which are mostly small and medium sized, that canopy specialists are rare and mostly insectivorous species given the low vertical structure of the habitat and that cross-season comparison was a mail goal of the study, we consider that faecal analysis of mist-netted bird is the best sampling method available to obtain community wide seed dispersal estimates (Wheelwright et al. 1984) and that any bias is not seriously undermining the interpretation of the results. This method has been widely used in the reconstruction of seed dispersal by birds (e.g. Gorchov et al. 1993, 1995; Heleno et al. 2011, 2013b; González-Castro et al. 2012; Spotswood et al. 2012).
Seed dispersal
Blackcap was the main seed disperser in all seasons, followed by Blackbird, Song Thrush and Robin. This is in accordance with studies that demonstrated that species of the genus Sylvia and Turdus and also E. rubecula are the most important seed dispersers in other Mediterranean areas (Jordano 1982, 1994; Herrera 1984, 1995; Fuentes 1994). In particular, the Blackcap is one of the most significant seed dispersers in the Mediterranean region, particularly during autumn and winter, feeding almost exclusively on fruits as their alternative food sources—insects—become harder to find (Jordano and Herrera 1981; Herrera 1998). As a generalist frugivorous bird species, the Blackcap is widely abundant throughout Europe and has a broad niche, ingesting fruits of highly different sizes (Jordano 1982; Herrera 1984, 1995). Therefore, it has a high potential to explore new available resources and increase population and ecological importance in novel forest ecosystems in Europe. Our study supports its disproportionate importance as a main seed disperser suggesting that plants might suffer limited dispersal if its populations are reduced.
Furthermore, emphasis should be given to the relative importance of Song Thrush in these seed dispersal network. In Portugal, this species is much more abundant during the winter due to the income of migrants from the northern European breeding populations. However, their breeding (i.e. summer) population in Portugal expanded in the last two decades (Equipa Atlas 2008; Catry et al. 2010). Species range expansions such as this are likely to change interaction networks such as seed dispersal. Our results suggest that this species has a potential to become one of the main local seed dispersers if the population trend continues growing. Even if the specific outcomes of such increases are difficult to predict, one could hypothesize they would be mostly beneficial, at least initially, by assisting plant dispersal, while negative effects either by competition for food with other birds or by spreading alien seeds could be possible only on extreme scenarios.
There are relevant migratory species that also disperse seeds (although less frequently, mainly because they were only present during a short period), such as the Pied Flycatcher and Garden Warbler which pass through the study area during autumn, on the way to wintering grounds in Africa, and the Melodious Warbler (Hippolais polyglotta) which breeds in the study area (Equipa Atlas 2008). Regardless of their overall low presence in the study site, such migratory birds may play an important role in long-distance dispersal of seeds. In this respect, novel ecosystems can also represent a threat to more pristine habitats as they might serve a source of alien seeds which are then transported to new focus of invasion by migrating birds.
As expected, despite the greater availability of fruits in summer, fruit consumption by birds was greater in winter, which can be explained by the decrease in other food sources such as insects (Herrera 1998; Parrish 2000) and by the arrival of several wintering populations (e.g. Blackcap, Robin and Song Thrush). Our study suggests that even in a highly managed urban park, with a relatively small biodiversity, seed dispersal networks can be highly complex and dynamic, with fruiting plants producing fruits asynchronously and dispersers relying on fruits particularly during the winter and the autumn migration. It is interesting to note that the alien P. americana formed a relevant part of the diet of several birds, particularly in the summer. This alien plant species is not locally dominant, and it is unlikely to represent a key dietary resource for birds. However, it seems undisputable that alien fruits can be important resources for many bird species and that this importance tends to increase with fruit abundance, likely resulting in positive feedbacks between plants and dispersers. We can envisage that when invasions become so widespread that alien species replace most of the native resources for animals, the management of the invasion becomes more complex, as eradication might have serious negative effects (Ceia et al. 2011).
The Blackcap was consistently important as a main disperser of both native and alien seeds throughout the year. This species can be regarded as a key-stone species in the seed dispersal in Choupal. New species combinations are an integral part of interaction networks in novel ecosystems and will influence native biota in direct and indirect ways. Native and alien species will interact leading to new ecological outcomes.
Although most interactions were detected in summer, the season with higher diversity and abundance of fruits (Table 1), the connectance for the autumn and winter networks was higher, indicating that there was a more efficient use of the resources (i.e. higher number of realized links among all possible links). Interaction evenness was higher in summer which means that there was a more homogeneously distribution of the frequency of links in these networks, whereas in autumn and winter, a few highly frequent interactions dominated the network, particularly Blackcap dispersing C. australis in the autumn and H. helix in the winter.
The implementation of a network approach allowed us to look for emergent properties of biological communities by means of standard measurements and to link those patterns to community stability and functioning (Bascompte and Jordano 2007; Thompson et al. 2012). Furthermore, given the inherent complexity of seed dispersal processes, interaction network offers a theoretical framework to look for general patterns without losing information by clumping species into functional groups (Memmott 2009).
Conclusions
Understanding how novel ecosystems work is increasingly important. Because environmental and biotic changes are occurring rapidly, it is fundamental to evaluate the best available options for their management in order to maintain their ecological functions. Although our results revealed a relatively complex seed dispersal network, a single bird species—Blackcap—had a central role in this novel ecosystem, which can make it more vulnerable. Their strong interaction with P. americana suggests that alien plants can attract native dispersers taking advantage of the highly generalist dispersal system.
References
Alberti M (2005) The effects of urban patterns on ecosystem function. Int Reg Sci Rev 28(2):168–192
Alberti M, Marzluff JM, Shulenberger E, Bradley G, Ryan C, Zumbrunnen C (2003) Integrating humans into ecology: opportunities and challenges for studying urban ecosystems. Bioscience 53(12):1169–1179
Bascompte J, Jordano P (2007) Plant–animal mutualistic networks: the architecture of biodiversity. Annu Rev Ecol Evol Syst 38:567–593
Barnosky AD, Hadly EA, Bascompte J, Berlow EL, Brown JH, Fortelius M, Getz WM, Harte J, Hastings A, Marquet PA, Martinez ND, Mooers A, Roopnarine P, Vermeij G, Williams JW, Gillespie R, Kitzes J, Marshall C, Matzke N, Mindell DP, Revilla E, Smith AB (2012) Approaching a state shift in Earth's biosphere. Nature 486(7401):52–58
Bersier LF, Banasek-Richter C, Cattin MF (2002) Quantitative descriptors of food-web matrices. Ecology 83(9):2394–2407
Bibby CJ, Burgess ND, Hill DA, Mustoe S (2000) Bird census techniques, 2nd edn. Academic Press, London
Catry P, Matias R, Elias G, Costa H (2010) Aves de Portugal—ornitologia do território continental. Assírio & Alvim, Lisbon
Ceia RS, Sampaio HL, Parejo SH, Heleno RH, Arosa ML, Ramos JA, Hilton GM (2011) Throwing the baby out with the bathwater: does laurel forest restoration remove a critical winter food supply for the critically endangered Azores bullfinch? Biol Invasions 13:93–104
Cordeiro NJ, Howe HF (2003) Forest fragmentation severs mutualism between seed dispersers and an endemic African tree. Proc Natl Acad Sci USA 100(24):14052–14056
Dormann C, Gruber B, Frund J (2008) Introducing the bipartite package: analysing ecological networks. R News 8(2):8–11
Dunn E, Ralf CJ (2004) Use of mist nets as a tool for bird population monitoring. Stud Avian Biol 29:1–6
Equipa Atlas (2008) Atlas das aves nidificantes em Portugal (1999–2005). Instituto da Conservação da Natureza e da Biodiversidade, Sociedade Portuguesa para o Estudo das Aves, Parque Natural da Madeira, Secretaria Regional do Ambiente e do Mar. Assírio & Alvim, Lisboa
Estades CF, Escobar MH, Tomasevicm JA, Vukasovic MA, Paez M (2006) Mist-nets versus point counts in the estimation of forest bird abundances in south-central Chile. Ornitol Neotrop 17:203–212
Fuentes M (1994) Diets of fruit-eating birds: what are the causes of interspecific differences? Oecologia 97(1):134–142
Fuentes M (2000) Frugivory, seed dispersal and plant community ecology. Trends Ecol Evol 15(12):487–488
González-Castro A, Traveset A, Nogales M (2012) Seed dispersal interactions in the Mediterranean region: contrasting patterns between islands and mainland. J Biogeogr 39(11):1938–1947
Gorchov DL, Cornejo F, Ascorra C, Jaramillo M (1993) The role of seed dispersal in the natural regeneration of rain-forest after strip-cutting in the Peruvian Amazon. Vegetatio 108:339–349
Gorchov DL, Cornejo F, Ascorra C, Jaramillo M (1995) Dietary overlap between frugivorous birds and bats in the Peruvian Amazon. Oikos 74:235–250
Heleno RH, Ross G, Everard A, Ramos JA, Memmott J (2011) On the role of avian seed predators as seed dispersers. Ibis 153:199–203
Heleno RH, Olesen JM, Nogales M, Vargas P, Traveset A (2013a) Seed-dispersal networks in the Galápagos and the consequences of plant invasions. Proc R Soc Lond B Biol Sci 280(1750):20122112
Heleno RH, Ramos JA, Memmott J (2013b) Integration of exotic seeds into an Azorean seed dispersal network. Biol Invasions 15:1143–1154
Herrera CM (1984) A study of avian frugivores, bird-dispersed plants, and their interaction in mediterranean scrublands. Ecol Monogr 54(1):1–23
Herrera CM (1995) Plant-vertebrate seed dispersal systems in the Mediterranean: ecological, evolutionary, and historical determinants. Annu Rev Ecol Syst 26:705–727
Herrera CM (1998) Long-term dynamics of Mediterranean frugivorous birds and fleshy fruits: a 12-year study. Ecol Monogr 68(4):511–538
Hobbs RJ, Arico S, Aronson J, Baron JS, Bridgewater P, Cramer VA, Epstein PR, Ewel JJ, Klink CA, Lugo AE, Norton D, Ojima D, Richardson DM, Sanderson EW, Valladares F, Vilà M, Zamora R, Zobel M (2006) Novel ecosystems: theoretical and management aspects of the new ecological world order. Glob Ecol Biogeogr 15(1):1–7
Hobbs RJ, Higgs E, Harris JA (2009) Novel ecosystems: implications for conservation and restoration. Trends Ecol Evol 24(11):599–605
Hooper DU, Adair EC, Cardinale BJ, Byrnes JEK, Hungate BA, Matulich KL, Gonzalez A, Duffy JE, Gamfeldt L, O'Connor MI (2012) A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486(7401):105–108
Howe HF, Miriti MN (2004) When seed dispersal matters. Bioscience 54(7):651–660
Howe HF, Smallwood J (1982) Ecology of seed dispersal. Annu Rev Ecol Syst 13:201–228
Jordano P (1982) Migrant birds are the main seed dispersers of blackberries in southern Spain. Oikos 38(2):183–193
Jordano P (1994) Spatial and temporal variation in the avian-frugivore assemblage of Prunus mahaleb—patterns and consequences. Oikos 71(3):479–491
Jordano P, Herrera CM (1981) The frugivorous diet of blackcap populations Sylvia atricapilla wintering in southern Spain. Ibis 123(4):502–507
Kremen C, Hall G (2005) Managing ecosystem services: what do we need to know about their ecology? Ecol Lett 8(5):468–479
Kuussaari M, Bommarco R, Heikkinen RK, Helm A, Krauss J, Lindborg R, Ockinger E, Partel M, Pino J, Roda F, Stefanescu C, Teder T, Zobel M, Steffan-Dewenter I (2009) Extinction debt: a challenge for biodiversity conservation. Trends Ecol Evol 24(10):564–571
Marzluff JM, Bowman R, Donnelly R (2001) Avian ecology and conservation in an urbanizing World. Kluwer Academic Publishers, Norwell, Massachusetts, USA
McKinney ML, Lockwood JL (1999) Biotic homogenization: a few winners replacing many losers in the next mass extinction. Trends Ecol Evol 14(11):450–453
Memmott J (2009) Food webs: a ladder for picking strawberries or a practical tool for practical problems? Philos Trans R Soc Lond B Biol Sci 364(1524):1693–1699
Moran C, Catterall CP, Kanowski J (2009) Reduced dispersal of native plant species as a consequence of the reduced abundance of frugivore species in fragmented rainforest. Biol Conserv 142(3):541–552
Nathan R, Muller-Landau HC (2000) Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends Ecol Evol 15(7):278–285
Norte AC, Ramos JA, Sampaio HL, Sousa JP, Sheldon BC (2010) Physiological condition and breeding performance of the Great Tit. Condor 112(1):79–86
Pagen RW, Thompson FR, Burhans DE (2002) A comparison of point-count and mist-net detections of songbirds by habitat and time-of-season. J Field Ornithol 73:53–59
Parrish JD (2000) Behavioral, energetic, and conservation implications of foraging plasticity during migration. Stud Avian Biol 20:53–70
R Development Core Team (2010) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Ralph CJ, Geupel GR, Pyle P, Martin TE, DeSante DF (1993) Handbook of field methods for monitoring landbirds. Pacific Southwest Research Station, Forest Service, U.S. Department of Agriculture, Albany, USA
Rebelo F (2003) Riscos naturais e acção antrópica. Imprensa da Universidade de Coimbra, Coimbra
Seastedt TR, Hobbs RJ, Suding KN (2008) Management of novel ecosystems: are novel approaches required? Front Ecol Environ 6(10):547–553
Sekercioglu CH (2006) Increasing awareness of avian ecological function. Trends Ecol Evol 21(8):464–471
Silva J, Simões A, Nossa L, Campino R, Rosmaninho R, Paiva J, Figueira L, Cruz F, Ramalheira P (2011) Plano de gestão florestal e orientação da utilização pública da Mata Nacional do Choupal. Instituto da Conservação da Natureza e Biodiversidade, Reserva Natural do Paul de Arzila, Arzila
Spotswood EN, Meyer J-Y, Bartolome JW (2012) An invasive tree alters the structure of seed dispersal networks between birds and plants in French Polynesia. J Biogeogr 39(11):2007–2020
Thompson RM, Brose U, Dunne JA, Hall RO, Hladyz S, Kitching RL, Martinez ND, Rantala H, Romanuk TN, Stouffer DB, Tylianakis JM (2012) Food webs: reconciling the structure and function of biodiversity. Trends Ecol Evol 27(12):689–697
Vilà M, Ibáñez I (2011) Plant invasions in the landscape. Landsc Ecol 26(4):461–472
Vitousek PM, D’Antonio CM, Loope LL, Westbrooks R (1996) Biological invasions as global environmental change. Am Sci 84(5):468–478
Wang Y, Finch DM (2002) Consistency of mist netting and point counts in assessing lanbird species richness and relative abundance during migration. Condor 104:59–72
Wang BC, Smith TB (2002) Closing the seed dispersal loop. Trends Ecol Evol 17(8):379–386
Wheelwright NT, Haber WA, Murray KG, Guindon C (1984) Tropical fruit-eating birds and their food plants—a survey of a Costa Rican lower montane forest. Biotropica 16:173–192
Whelan CJ, Wenny DG, Marquise RJ (2008) Ecosystem services provided by birds. In: Year in Ecology and Conservation Biology 2008, vol 1134. Ann N Y Acad Sci pp 25–60
Wolfram Research (1999) Mathematica, version 6.0. Champaign
Acknowledgments
We are grateful to Miguel Araújo for his valuable help during the bird capturing sessions, to Cristina Nabais for commenting early versions of this manuscript an to the Portuguese ringing Scheme (CNA/ICNF) for providing bird rings. RH was supported by the Grant SFRH/BD/77746/2011 of the Fundação para Ciência e Tecnologia (Portugal) and the Marie Curie Career Integration Grant FP7-PEOPLE-2012-CIG- 321794 (European Union).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Weiskittel.
Appendices
Appendix 1
See Table 2.
Appendix 2
See Table 3.
Rights and permissions
About this article
Cite this article
Cruz, J.C., Ramos, J.A., da Silva, L.P. et al. Seed dispersal networks in an urban novel ecosystem. Eur J Forest Res 132, 887–897 (2013). https://doi.org/10.1007/s10342-013-0722-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-013-0722-1