
Abstract
Migratory birds use compass systems derived from the geomagnetic field, the stars, the sun and polarized light patterns. We tested whether birds use a single underlying reference system for calibration of these compasses and, specifically, whether sunset and sunrise polarized light cues from the region of the sky near the horizon are used to calibrate the magnetic compass. We carried out orientation experiments with Savannah sparrows, Passerculus sandwichensis, in Alaska during autumn migration 2005, and compared the magnetic orientations of individual birds before and after exposure to conflicting information between magnetic and celestial cues. Birds exposed to an artificially shifted polarization pattern (±90° shift relative to the natural condition) for 1 h at local sunrise or sunset recalibrated their magnetic compass, but only when given access to the artificial polarization pattern near the horizon. Birds exposed to a 90° clockwise-shifted magnetic field for 1 h at solar noon did not recalibrate their magnetic compass. These results indicate that migratory birds calibrate their magnetic compass using the skylight polarization pattern vertically intersecting the horizon at sunrise and sunset. In conjunction with earlier work showing that sun and star compass calibrations are secondarily derived from magnetic and polarized light cues, our findings suggest that polarized light cues near the horizon at sunrise and sunset provide the primary calibration reference for the compass systems of migratory songbirds.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Migratory songbirds use multiple compasses for orientation during migration, including magnetic, star, solar and polarized light compasses (e.g., Emlen 1970; Able 1982; Moore 1987; Schmidt-Koenig 1990; Munro and Wiltschko 1995; Wiltschko and Wiltschko 1995). Depending on which compass cues birds rely on during migration, they are led along different migration routes (Alerstam and Gudmundsson 1999; Alerstam 2001; Alerstam et al. 2001; Bingman et al. 2003; Muheim et al. 2003; Åkesson and Hedenström 2007). The relationships between the different compass routes change along the migratory journey due to changes in declination, resulting from differences in the relative positions of the magnetic and geographic poles. Since cue availability changes with weather conditions, season, time of day, and latitude, birds must calibrate the different compasses with respect to a common reference both before and during migration to avoid navigational errors. The common reference can be one of the compass systems that has primacy over the others, or an independent source of directional information. Despite three decades of intensive research, the reference system birds use, how they integrate information from the different compass systems, and how often the different compasses are calibrated against each other are all still poorly understood.
Research on the integration of magnetic and celestial compass cues has produced variable and contradictory findings (for reviews see Able 1993; Åkesson 1994; Wiltschko et al. 1997, 1998; Wiltschko and Wiltschko 1999; Muheim et al. 2006a). During the pre-migratory season, most studies show that daytime/twilight celestial cues are given saliency over magnetic cues; i.e., both juvenile and adult birds recalibrate their magnetic compasses when exposed to conflicting information between magnetic and daytime/twilight celestial cues (Able and Able 1990b, 1993; Prinz and Wiltschko 1992; Weindler and Liepa 1999). In addition, no recalibration of celestial compasses is evident when birds exposed to a cue conflict are tested without access to information from the magnetic field (Bingman 1984; Able and Able 1997). The magnetic compass in turn seems to be the calibration reference for the star compass, at least in juvenile birds, since exposure to conflicting stellar and magnetic cues resulted in recalibration of the star, but not the magnetic compass (Wiltschko and Wiltschko 1976; Beason 1987; Weindler et al. 1998, but see Able and Able 1990a). Also, the star compass by itself only seems to contain information about the geographic north–south axis, requiring magnetic compass information to establish the population-specific migratory direction (Weindler et al. 1996, 1997). During migration, a majority of experiments suggest that birds use magnetic cues as their primary reference, since they appear to follow a magnetic field shift when exposed to a conflict between magnetic and celestial cues in orientation funnels (for references see Tables 2, 3 in Muheim et al. 2006a).
In a recent review of the cue conflict literature, we proposed that the contradictory findings obtained in different studies can be ascribed to differences in cue availability during exposure to the cue conflict (Muheim et al. 2006a). In particular, recalibration of the magnetic compass occurred during both the premigratory and migratory periods when birds exposed to conflicting information between magnetic and celestial light cues were able to see polarized light cues from the region of the sky near the horizon at sunrise or sunset. This hypothesis is consistent with two studies that reported recalibration of the magnetic compass during migration (Able and Able 1995a; Cochran et al. 2004). In both studies, the birds were exposed to the cue conflict in open cages, where they had access to a full view of the horizon, as typically done in experiments during the premigratory season. Other studies carried out during migration that did not obtain evidence for recalibration of the magnetic compass exposed the birds to the cue conflicts in orientation cages/funnels that blocked the view of the sky near the horizon. Thus, access to sunset cues, and possibly also sunrise cues, near the horizon may provide a better explanation of the pattern found in the literature than migration season, and such cues seem to be crucial for magnetic compass calibration (Muheim et al. 2006a).
The aim of the present study was to investigate whether migratory songbirds tested during migration recalibrate their magnetic compass after exposure to conflicting magnetic and polarized light cues under the following conditions: (a) polarization pattern shifted ± 90° relative to the natural magnetic field at sunrise, with view of the horizon; (b) polarization pattern shifted ±90° relative to the natural magnetic field at sunset, with view of the horizon; (c) polarization pattern shifted ±90° relative to the natural magnetic field at sunset, without view of the horizon, and; (d) magnetic field shifted +90° relative to the natural polarization pattern at solar noon.
Methods
We carried out orientation experiments with Savannah sparrows, Passerculus sandwichensis, at Kanaryarmiut Field Station (N61°22′, W165°08′) in the Yukon Delta National Wildlife Refuge, Alaska, USA, during autumn migration (11 August–21 September 2005). The Savannah sparrows of the nominate subspecies P. s. sandwichensis caught in the vicinity of the field station were both juvenile and adult birds from the local breeding population and migrants on passage. This subspecies winters in coastal southwestern British Columbia to western, central California (Pyle et al. 1987) and is thus expected to depart from the Yukon Delta in easterly to southeasterly migratory directions (see the Supporting Online Material in Muheim et al. 2006b).
The birds were kept indoors with access to the natural daylight scheme, but no view of celestial cues. Magnetic orientation was tested in orientation funnels covered with milky Plexiglas sheets and lined with typewriter correction (Tipp-Ex) paper in a translucent tent (Hansen Weatherport, Inc., Gunnison, CO, USA). All orientation experiments were carried out at sunset and lasted for 1 h. During that time the birds left scratch marks on the Tipp-Ex paper, which were analyzed by a visual estimation method (see below). Individual birds orienting towards the expected migratory direction (SE ± 90°) without access to celestial cues were exposed to experimental conditions (see “Experimental exposures” below) providing conflicting information between magnetic and daytime/twilight celestial cues. After exposure, the birds were tested again for magnetic orientation. Birds not significantly directed or inactive during an experiment were tested on subsequent evenings until they showed a valid direction (see below for criteria), for a maximum of four times. After completion of an experiment, the birds were set outside for several hours around sunset and/or sunrise to recalibrate the compasses back to the natural alignment (see exception under “Polarization pattern shifted ±90° at sunset without view of horizon” below). After recalibration, the birds were again tested for magnetic compass orientation until they showed a seasonally appropriate initial direction, before being exposed to the next experimental condition. Between exposures and experiments, the birds were kept indoors and were not allowed to see any outdoor cues, so that they could not recalibrate any compasses. All experiments were approved by the US Fish and Wildlife Service (permit #05-YDNWR-02).
Experimental exposures
Polarization pattern shifted ±90° at sunrise and/or sunset with view of horizon
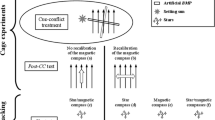
To simulate the polarization pattern at sunrise and sunset, when the band of maximum polarization perpendicularly intersects with the horizon, the birds were put into a cardboard box (30 × 32 × 32 cm) with four windows covered with netting. This box was placed in the center of a larger cardboard box (45 × 62 × 39 cm) with each window covered with a pseudo-depolarizing (outside) and a polarizing filter (inside). In the two windows on the short axis of the box the e-vector was aligned vertically, while the e-vector in the two windows on the long axis was horizontally aligned (for details see the Supporting Online Material in Muheim et al. 2006b). By depolarizing the light before it passed through the polarizing filters, we avoided any effects on light intensity caused by the interaction of the polarizing filters with the natural skylight polarization. The axis with the two vertically aligned polarizing filters was directed towards the sunrise/sunset position, so that the birds perceived the band of maximum polarization shifted ±90° relative to the natural condition at the time of exposure (Fig. 1a,b; Column 2). All exposures took place between 30 min before and 30 min after sunrise or sunset, in an open area about 100 m to the north of the field station. Birds exposed to the experimental condition at sunrise were tested the same evening around sunset, while birds exposed to an experimental condition during sunset were tested during the following evening.

Magnetic responses of Savannah sparrows exposed to conflicting information between the magnetic field and polarized light patterns. a Polarization pattern shifted ±90° at sunrise, with view of the horizon. b Polarization pattern shifted ±90° at sunset, with view of the horizon. c Polarization pattern shifted ±90° at sunset, without view of the horizon. d Magnetic field shifted +90° at solar noon. Column 1: initial magnetic orientation before exposure to cue conflict. Column 2: cue conflict condition during experimental exposure. Column 3: magnetic orientations after cue conflict exposure. Column 4: deviations from initial orientation [orientation after exposure (Column 3) − initial orientation (Column 1) = initial orientation set to 0°]. Each data point in the circular graphs gives the magnetic orientation of an individual bird (Columns 1 and 3) or the deviation from the initial response (Column 4). The arrows give the mean orientation of the group of birds and the length of each arrow is a measure of the concentration (r) of the group drawn relative to the radius of the circle = 1. Double-arrows indicate axially distributed samples. Dashed lines give the 95% confidence interval for significantly directed samples according to the Rayleigh test. Diagrams in Column 2 show sun positions and bands of maximum polarization at sunrise (a), sunset (b and c) and solar noon (d); dark gray circles in the centers of a and b and the outer ring in c indicate sections of the sky not visible to the birds during exposure. Large triangles outside of the circles in the graphs in column 4 give the expected response (light gray sunrise, medium gray sunset). For details see Table 1
Polarization pattern shifted ±90° at sunset without view of horizon
A small number of birds were exposed to a ±90° shifted artificial polarization pattern at sunset, with the horizon blocked from view, simulating the polarization cues perceived by birds tested in orientation funnels (Fig. 1c, Column 2). These exposures immediately followed a sunrise or sunset exposure with view of the horizon (a or b), without intermittent recalibration, which is an exception to the general experimental design. The birds were placed into a cardboard box (31 × 45 × 33 cm) with a single, round polarizing filter at the top (diameter 31 cm). They were able to see a ∼105° view of the overhead sky when sitting on one of the perches at the bottom of the box and ∼142° when jumping or flying up towards the ceiling of the box (in conventional orientation funnels, views of the sky vary between 90 and 160°; for a review see Muheim et al. 2006a). The e-vector of the polarizing filter was aligned axially 90° relative to the sun’s position along the natural alignment of the band of maximum polarization. Thus, birds that had recalibrated the magnetic compass in the earlier cue-conflict exposure (Fig. 1a or b) would be expected to calibrate the magnetic compass back to the natural calibration. Exposures lasted from 30 min before to 30 min after sunset and the orientation of the birds was tested during the following evening (for details see the Supporting Online Material in Muheim et al. 2006b).
Magnetic field shifted ±90° at solar noon with view of horizon
Since the skylight polarization pattern is more complex at solar noon than at sunrise and sunset (the band of maximum polarization intersects the horizon at an angle ≠90°), we created a cue conflict using a Helmholtz coil (80 × 80 cm) to shift the magnetic field 90° clockwise relative to the natural celestial cues at solar noon (Fig. 1d, Column 2). The coil was placed in an open area about 100 m to the north of the field station (same position as sunrise/sunset polarization exposures). An 18 V car battery was used as power source. The birds were placed into a cardboard box (35 × 37 × 36 cm) with a window on each side and centered so that the horizontal sitting perches were approximately in the middle of the coil. Exposures took place 30 min before to 30 min after 14:55 local time (solar noon: sun azimuth = 180°, sun elevation = 30°) during partly overcast skies (cloud cover 4/8). Birds exposed to the experimental exposure at solar noon were tested the same evening around sunset.
Data evaluation and statistics
We used a visual method to determine the direction chosen by each bird (Mouritsen 1998). A person blind to the experimental condition and the position of the north mark visually estimated the median of the circular distribution of scratches on the Tipp-Ex paper. Each paper was given a score for activity from 0 (0–50 scratches) to 4 (>2,000 scratches) and a score for estimation accuracy from 0 (>45° estimation accuracy) to 4 (0–5° estimation accuracy; for details see the Supporting Online Material in Muheim et al. 2006b). We excluded experiments with activity = 0 or concentration = 0 or total score (activity + concentration) <3, thus including only experiments with well-directed migratory orientation (see also Mouritsen 1998). This estimation method has been shown to be highly repeatable and in close agreement with the more conventional method of counting scratches in different sectors across the Tipp-Ex paper (Mouritsen and Larsen 1998; M. Tarka, R. Muheim, S. Åkesson, unpublished data).
We applied circular statistics to test whether the directional responses of the birds were significantly directed (Rayleigh test, Batschelet 1981) and the method of doubling of angles to test whether a group was axially distributed (r axial > r unimodal). Since we were interested in the response of each individual bird to the exposure, we calculated the difference between each individual’s initial orientation and the direction chosen after the exposure. We used the 95% confidence intervals to examine whether the observed difference agreed with the expected shift. Differences between groups were tested with the nonparametric circular Watson U 2 test (Batschelet 1981).
Results
Savannah sparrows exposed to an artificially shifted polarization pattern (±90° shift relative to the natural condition) for 1 h around sunrise or sunset, with full view of the surroundings, including the horizon, recalibrated their magnetic compass. The difference between the individual magnetic orientation after exposure (Column 1 of Fig. 1a,b) and the orientation before exposure (Column 3 of Fig. 1a,b) was axially distributed along the 90–270° axis (Table 1). The 95% confidence intervals included the ±90° shifts expected when the magnetic compass is recalibrated at sunrise or sunset relative to the natural polarization pattern at the same time of day (Column 4 of Fig. 1a,b). The responses of birds that recalibrated their magnetic compass after being exposed to a shifted polarization pattern at sunrise and then at sunset, or vice versa (but exposed to the natural conditions in order to calibrate their magnetic compass back in-between the two exposures) were not statistically different to birds exposed to only one of the exposure conditions (sunrise: U 2 = 0.03, P > 0.5; sunset: U 2 = 0.03, P > 0.5; see Table 1).
The sample of six Savannah sparrows exposed to a ±90° shifted artificial polarization pattern at sunset, with the region of sky immediately above the horizon blocked from view, showed no shift in response, but instead kept orienting towards the previously chosen magnetic direction (Column 4 of Fig. 1c, Table 1). Similarly, birds exposed to a +90° shifted magnetic field for 1 h at solar noon did not show a 90° shift in orientation; instead they either did not shift at all or they shifted by 180° (Column 4 of Fig. 1d, Table 1).
Discussion
Our results show that Savannah sparrows recalibrate their magnetic compass repeatedly with respect to polarized light cues at both sunrise and sunset, but not at solar noon. Twilight celestial cues close to the horizon provide the crucial information necessary for magnetic compass calibration. In conjunction with other studies, our results confirm that recalibration occurs both prior to and during migration, and in both juvenile and adult birds.
In the majority of published cue conflict experiments carried out during migration, birds did not recalibrate their magnetic compasses, but instead followed the shift of the magnetic field and recalibrated their celestial compasses (for references see Muheim et al. 2006a). A review of the cue-conflict literature suggested that birds in many of these studies did not have access to all crucial celestial cues during the cue-conflict exposures (Muheim et al. 2006a). Birds exposed to the cue conflict in orientation funnels, which therefore had their view of the polarization pattern near the horizon blocked (funnels allow an average view of the sky around the zenith of 90–160°), typically followed the shift of the magnetic field. They, therefore, did not recalibrate the magnetic compass. In contrast, in all but one experiment (Able and Able 1990b) where the birds were exposed to the cue conflict in an open cage, allowing them to see the sky down to the horizon, recalibration of the magnetic compass was observed (Bingman 1983; Able and Able 1990b, 1995a; Prinz and Wiltschko 1992; Weindler and Liepa 1999; Cochran et al. 2004). When our birds were exposed to an artificial polarization pattern that did not include the region of sky near the horizon, we observed no recalibration of the magnetic compass, confirming the importance of sunrise/sunset polarization cues near the horizon as the crucial calibration reference.Footnote 1 Thus, the failure to observe magnetic compass recalibration in many studies carried out during migration (for a review see Muheim et al. 2006a) is likely the result of restricted access to the celestial polarized light cues close to the horizon at sunrise or sunset.
Our results indicate that the polarized light pattern at sunrise or sunset is the primary calibration reference used by Savannah sparrows to recalibrate their magnetic compasses. Recalibration occurred after exposure to a shifted polarization pattern for as little as 1 h at both sunrise and sunset, but not at solar noon. This result agrees with evidence from earlier studies that celestial cues at times other than sunrise and sunset do not provide the cues needed to recalibrate the magnetic compass (Able and Able 1990b; Åkesson et al. 2002). In previous studies with Savannah sparrows reporting recalibration of the magnetic compass, the birds were exposed to the cue conflict for several days to weeks (Bingman 1983; Able and Able 1990b, 1993, 1995a, 1995b). Access to a number of celestial cues such as sunrise, sun and sunset during exposure to the cue conflict made it difficult for conclusive predictions about the times of day that recalibration takes place and about the celestial cues used for recalibration. Cue calibration studies with other species have been equally inconclusive (Prinz and Wiltschko 1992; Weindler and Liepa 1999), with the exception of Cochran et al. (2004), who showed that two species of thrush, the grey-cheeked thrush, Catharus minimus, and Swainson’s thrush, Catharus ustulatus, recalibrated their magnetic compasses after exposure to a shifted magnetic field for variable periods of up to ∼3 h, including sunset.
Our Savannah sparrows did not use the position of the sun itself as a recalibration reference, since the natural position of the sun was unchanged and at least partially visible to some of the birds. This finding is consistent with the results of an earlier study, in which Savannah sparrows exposed to a shifted magnetic field under a depolarizing filter that eliminated polarized light cues, but allowed detection of the sun and star patterns, did not recalibrate their magnetic compass, while control birds exposed to the same treatment, but without depolarizing filters, recalibrated the magnetic compass (Able and Able 1993). However, these birds were given multiple exposures lasting for 3–4 h throughout the day, thus the time(s) of day the recalibration occurred could not be determined. In a later experiment (Able and Able 1995b), birds exposed to artificially shifted polarization patterns for 1 h at sunrise and sunset were also found to recalibrate their magnetic compass, indicating that the polarization pattern at sunrise and/or sunset provided the calibration reference, but did not exclude the possibility that the same cues could also lead to a recalibration at other times of day (Able and Able 1995b). Our results exclude the use of polarized light cues at other times of day aside from sunrise and sunset; the reasoning for this is discussed below.
Why do birds use polarized light cues from the sunrise or sunset sky and not from other times of day? Only at sunrise and sunset, the band of maximum polarization (BMP) and the e-vector pass directly through the zenith and are aligned vertically on the horizon. The intersections of the BMP with the horizon are independent of topography and horizon height, and their use as calibration reference would not require a time-compensation mechanism (Fig. 2). This is not true for the positions at which the sun disk appears above the horizon at sunrise and disappears below the horizon at sunset, which vary depending on the topography and vegetation height, making it difficult for birds to determine the exact timing and position of sunset. A problem with using sunrise/sunset cues is that sunrise/sunset positions shift with season and latitude, which can result in curving migratory paths that are only adaptive under specific circumstances (see below). Based on earlier work with pigeons (Phillips and Waldvogel 1988), we therefore propose the theory that birds average the information from successive sunrises and sunsets. Averaging the intersections of the band of maximum polarization with the horizon during a successive sunrise and sunset from the same location would enable migratory birds to derive a true geographic reference system that is “fixed” with respect to the North–South meridian at any location on Earth and that is independent of latitude and time of year (Fig. 3). Periodic updating of the relationship between the polarization patterns at sunrise and sunset (their angular “split” on either side of the meridian) would make it possible, for example when bad weather makes averaging impossible, to use either sunrise or sunset cues alone to estimate the reference direction and to calibrate other compass systems. Birds exposed to only one exposure to a cue conflict, as done in most cue-conflict experiments, show a full shift and seem to average only when exposed to the cue conflict on a consecutive sunrise/sunset from the same location.

Three-dimensional illustration of the band of maximum polarization (BMP) at sunrise (a) and sunset (b), intersecting the horizon vertically. Course of setting sun in locations with different horizon features (c), illustrating that the position/timing of the local sunset depends on topography

Two-dimensional representations of the band of maximum polarization (BMP) at sunrise (a) and sunset (b). The deviations of the band of maximum polarization from the geographic North–South axis at sunrise and sunset are equal in magnitude, but opposite in direction (c). Therefore, by averaging the alignments of the BMP at these two times of day from the same location (regardless of time of year and latitude) it is possible to derive the geographic North–South axis (Phillips and Waldvogel 1988). Once the relative alignments of the BMPs at sunrise and sunset have been learned, the alignment at sunrise (d) or at sunset (e) alone can be used to derive the reference direction (e.g., gN) to calibrate other compass systems. However, periodic updating of the calibration reference, requiring access to polarized light cues at both sunrise and sunset, would be necessary to correct for seasonal and latitudinal changes in the alignments of the BMP (see Fig. 4)
Why should birds be averaging instead of using only sunrise or only sunset cues as independent calibration references? Changes in latitude and time of year produce opposite shifts in the alignments of the BMP at sunrise and sunset. For example, during autumn migration, sun position and the alignment of the BMP shift clockwise at sunrise and counter-clockwise at sunset. The use of only sunrise or only sunset as the independent calibration reference may be adaptive in species that use the polarization pattern at either sunrise or sunset alone, without averaging. Such a strategy can result in gradually curving migration routes, but it is only adaptive when only one cue and always the same (polarization pattern at only sunrise or only sunset) is used. In species like the Savannah sparrows that use both sunrise and sunset cues to recalibrate their magnetic compasses, failure to average the information from both times of the day would produce a “zig-zagging” migratory path, depending on whether the clear skies necessary to see the polarization pattern occurred most recently at sunrise or at sunset (Fig. 4). In contrast, averaging the alignments of the BMP at sunrise and sunset, as sky conditions permit, would eliminate seasonal and latitudinal variation and enable the birds to correct for this variation when forced to rely on either sunrise or sunset cues alone to update the calibrations of their other compass systems. The opportunistic use of sunrise and sunset cues is therefore not adaptive for species like Savannah sparrows that recalibrate their magnetic compass at both times of day.

Hypothetical migratory paths followed by a Savannah sparrow with a starting point at the approximate center of the breeding distribution for the subspecies P. s. sandwichensis in Alaska (64°50′N 147°50′W) on 20 August with an initial heading of 140°, using one of four strategies to calibrate its other compass systems. The hypothetical paths were determined assuming that the compass systems were recalibrated daily, weather conditions permitting, with a travel rate of 100 km/day for 40 days; changes in the alignments of the BMP at sunrise and sunset were calculated for the new date and location, and then used to calculate the new migratory direction. Between each step (dot/square) the birds were assumed to follow rhumb lines, i.e., constant geographic routes, which in this example do not significantly differ from a constant magnetic compass course. Light gray line of dots: the route followed by a bird using the horizon polarization pattern at sunrise only as a calibration reference. Dark gray line of squares: the route followed by a bird using the horizon polarization pattern at sunset only as a calibration reference. Line of mixed dots and squares: migratory route followed by a bird that opportunistically recalibrates the magnetic compass at sunrise and sunset, depending on cloud cover, without averaging the alignments of the BMP at these two times of day. Black dotted line: route followed by a migratory bird using polarized light cues at both sunrise and sunset for magnetic compass calibration and averaging the information from the two times of day, as suggested by our theory (from Muheim et al. 2006b)
Conclusions
Our findings suggest that polarized light cues near the horizon at sunrise and sunset provide the primary calibration reference for the compass systems of Savannah sparrows. Earlier work showed that sun and star compass calibrations are secondarily derived from magnetic and polarized light cues (Beason 1987; Bingman and Wiltschko 1988; Phillips and Moore 1992). This primary calibration reference would provide birds with an absolute, geographic directional system, which is independent of season and latitude. If our theory is correct, curving migratory routes and abrupt changes in migratory direction to circumfly major topographic features, i.e., oceans and mountain ranges, that regularly occur along natural migratory routes (e.g., Alerstam 2001; Alerstam et al. 2003), are likely to involve secondary adaptations rather than properties of the underlying calibration system in species like the Savannah sparrows that use both sunrise and sunset for recalibration of their magnetic compass.
References
Able KP (1982) Skylight polarization patterns at dusk influence migratory orientation in birds. Nature 299:550–551
Able KP (1993) Orientation cues used by migratory birds: a review of cue-conflict experiments. TREE 8:367–371
Able KP, Able MA (1990a) Calibration of the magnetic compass of a migratory bird by celestial rotation. Nature 347:378–380
Able KP, Able MA (1990b) Ontogeny of migratory orientation in the Savannah sparrow, Passerculus sandwichensis: calibration of the magnetic compass. Anim Behav 39:905–913
Able KP, Able MA (1993) Daytime calibration of magnetic orientation in a migratory bird requires a view of skylight polarization. Nature 364:523–525
Able KP, Able MA (1995a) Interactions in the flexible orientation system of a migratory bird. Nature 375:230–232
Able KP, Able MA (1995b) Manipulations of polarized skylight calibrate magnetic orientation in a migratory bird. J Comp Physiol A 177:351–356
Able KP, Able MA (1997) Development of sunset orientation cues in a migratory bird: no calibration by the magnetic field. Anim Behav 53:363–368
Åkesson S (1994) Comparative orientation experiments with different species of passerine long-distance migrants: effect of magnetic field manipulation. Anim Behav 48:1379–1393
Åkesson S, Hedenström A (2007) How migrants get there: migratory performance and orientation. BioScience 57:123–133
Åkesson S, Morin J, Muheim R, Ottosson U (2002) Avian orientation: effects of cue-conflict experiments with young migratory songbirds in the high Arctic. Anim Behav 64:469–475
Alerstam T (2001) Detours in bird migration. J Theor Biol 209:319–331
Alerstam T, Gudmundsson GA (1999) Bird orientation at high latitudes: flight routes between Siberia and North America across the Arctic Ocean. Proc R Soc Lond B 266:2499–2505
Alerstam T, Gudmundsson GA, Green M, Hedenström A (2001) Migration along orthodromic sun compass routes by arctic birds. Science 291:300–303
Alerstam T, Hedenström A, Åkesson S (2003) Long-distance migration: evolution and determinants. Oikos 103:247–260
Batschelet E (1981) Circular statistics in biology. Academic, London
Beason RC (1987) Interaction of visual and non-visual cues during migratory orientation by the bobolink, Dolichonyx oryzivorus. J Ornithol 128:317–324
Bingman VP (1983) Magnetic field orientation of migratory Savannah sparrows with different first summer experience. Behaviour 87:43–51
Bingman VP (1984) Night sky orientation of migratory pied flycatchers raised in different magnetic fields. Behav Ecol Sociobiol 15:77–80
Bingman VP, Wiltschko W (1988) Orientation of dunnocks, Prunella modularis, at sunset. Ethology 77:1–9
Bingman VP, Budzynski CA, Voggenhuber A (2003) Migratory systems as adaptive responses to spatial and temporal variability in orientation stimuli. In: Berthold P, Gwinner E, Sonnenschein E (eds) Avian migration. Springer, Berlin, pp 567–469
Cochran WW, Mouritsen H, Wikelski M (2004) Migrating songbirds recalibrate their magnetic compass daily from twilight cues. Science 304:405–408
Emlen ST (1970) Celestial rotation: its importance in the development of migratory orientation. Science 170:1198–1201
Hegedüs R, Åkesson S, Wehner R, Horváth G (2007a) Could Vikings have navigated under foggy and cloudy conditions by skylight polarization? On the atmospheric optical prerequisites of polarimetric Viking navigation under foggy and cloudy skies. Proc R Soc Lond A 463:1081–1095
Hegedüs R, Åkesson S, Horváth G (2007b) Polarization of overcast skies and implications for animal orientation: radiative transfer of the clear-sky E-vector pattern through thick ice and water clouds. J Opt Soc Am A (in press)
Moore FR (1987) Sunset and the orientation behaviour of migrating birds. Biol Rev 62:65–86
Mouritsen H (1998) Redstarts, Phoenicurus phoenicurus, can orient in a true-zero magnetic field. Anim Behav 55:1311–1324
Mouritsen H, Larsen ON (1998) Evaluating alternative methods for the analysis of Emlen funnel data. In: Mouritsen H (ed) Compasses and orientational strategies of night migrating passerine birds. Center of Sound Communication, Institute of Biology, Odense University, Odense, pp 115–124
Muheim R, Åkesson S, Alerstam T (2003) Compass orientation and possible migration routes of passerine birds at high arctic latitudes. Oikos 103:341–349
Muheim R, Moore FR, Phillips JB (2006a) Calibration of magnetic and celestial compass cues in migratory birds––a review of cue-conflict experiments. J Exp Biol 209:2–17
Muheim R, Phillips JB, Åkesson S (2006b) Polarized light cues underlie compass calibration in migratory songbirds. Science 313:837–839
Munro U, Wiltschko R (1995) The role of skylight polarization in the orientation of a day-migrating bird species. J Comp Physiol A 177:357–362
Phillips JB, Moore FR (1992) Calibration of the sun compass by sunset polarized light patterns in a migratory bird. Behav Ecol Sociobiol 31:189–193
Phillips JB, Waldvogel JA (1988) Celestial polarized patterns as a calibration reference for sun compass of homing pigeons. J Theor Biol 131:55–67
Prinz K, Wiltschko W (1992) Migratory orientation of pied flycatchers: interaction of stellar and magnetic information during ontogeny. Anim Behav 44:539–545
Pyle P, Howell SNG, Yunick RP, DeSante DF (1987) Identification guide to North American passerines. Slate Creek, Bolinas, CA
Schmidt-Koenig K (1990) The sun compass. Experientia 46:336–342
Weindler P, Liepa V (1999) The influence of premigratory experience on the migratory orientation of birds. In: Adams NJ, Slotow RH (eds) Proceedings of the 22nd International Ornithological Congress. Birdlife South Africa, Johannesburg, South Africa, pp 979–987
Weindler P, Wiltschko R, Wiltschko W (1996) Magnetic information affects the stellar orientation of young bird migrants. Nature 383:158–160
Weindler P, Baumetz M, Wiltschko W (1997) The direction of celestial rotation influences the development of stellar orientation in young garden warblers, Sylvia borin. J Exp Biol 200:2107–2113
Weindler P, Liepa V, Wiltschko W (1998) The direction of celestial rotation affects the development of migratory orientation in pied flycatcher, Ficedula hypoleuca. Ethology 104:905–915
Wiltschko W, Wiltschko R (1976) Interrelation of magnetic compass and star orientation in night-migrating birds. J Comp Physiol A 109:91–99
Wiltschko R, Wiltschko W (1995) Magnetic orientation in animals. Springer, Berlin
Wiltschko R, Wiltschko W (1999) Celestial and magnetic cues in experimental conflict. In: Adams NJ, Slotow RH (eds) Proceedings of the 22nd International Ornithological Congress. Birdlife South Africa, Johannesburg, South Africa, pp 988–1004
Wiltschko R, Wiltschko W, Munro U (1997) Migratory orientation in birds: the effects and after-effects of exposure to conflicting celestial and magnetic cues. In: Orientation and navigation––birds, humans and other animals. Royal Institute of Navigation, Oxford, pp 6.1–6.14
Wiltschko W, Weindler P, Wiltschko R (1998) Interaction of magnetic and celestial cues in the migratory orientation of passerines. J Avian Biol 29:606–617
Acknowledgments
We thank the Swedish Polar Research Secretariat, Brian J. McCaffery, Mike Rearden and the staff at the US Fish and Wildlife Service, Yukon Delta National Wildlife Refuge, for financial and logistic support. We are grateful to Thomas Alerstam for valuable comments on the manuscript. This work was financed by the Swedish Science Research Council and the Swedish Polar Research Secretariat (to SÅ). RM was funded by the Swiss National Science Foundation (postdoctoral fellowship) and JBP by the US National Science Foundation (IBN04–25712 and IBN02–16957). All experiments were approved by the US Fish and Wildlife Service (permit #05-YDNWR-02).
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H. Mouritsen.
Rights and permissions
About this article
Cite this article
Muheim, R., Åkesson, S. & Phillips, J.B. Magnetic compass of migratory Savannah sparrows is calibrated by skylight polarization at sunrise and sunset. J Ornithol 148 (Suppl 2), 485–494 (2007). https://doi.org/10.1007/s10336-007-0187-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-007-0187-4