
Abstract
Birds have evolved a mobile lifestyle in which vision is of major importance when controlling movements, avoiding predators, finding food and selecting mates. Birds have extraordinary colour vision and have been suggested to perceive the linear polarisation of light. Behavioural experiments support this idea, but still the exact physiological mechanism involved is not known. The twilight period, when the sun is near the horizon at sunrise and sunset, is of crucial importance for migrating birds. At this time millions of songbirds initiate migration when the degree of skylight polarisation is the highest and all compass cues are visible in a short range of time. The biological compasses are based on information from the stars, the sun and the related pattern of skylight polarisation, as well as the geomagnetic field, and may be recalibrated relative to each other. The celestial polarisation pattern near the horizon has been shown to be used in the recalibration of the magnetic compass, but conflicting results have been obtained in experiments with different bird species. For the future we should understand the physiological mechanisms of avian polarisation vision and investigate the interrelationship and calibrations between the different compasses, including the one based on the pattern of skylight polarisation. A conditioning paradigm may be fruitful, but the risk of introducing optical artefacts needs to be minimised in behavioural experiments, as well as in cage experiments with migratory birds.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others

Keywords
These keywords were added by machine and not by the authors. This process is experimental and the keywords may be updated as the learning algorithm improves.
1 Introduction
Birds are highly aerial animals and in their everyday life they are largely dependent on their vision. Some birds may use their vision while diving several hundreds of metres in cold oceanic waters, while others are climbing to 3,000 m altitude and navigate across continents by relying on celestial and magnetic compasses. Others inhabit dense tropical forests where they display to find a mate. On the outermost branches in tall trees, tiny warblers feed on insects and spiders, optimising their foraging efforts and locating their prey by vision. Some birds are only active at night or hide in caves. The width of avian visual adaptations to cope with these varying ecological situations is large (Goldsmith 1991). Bird eyes show an allometric relationship with brain and body size (Burton 2008). At the same time they constitute a major part of the head, and correspondingly, the optical lobes are a substantial part of the brain (Martin 1985; de Brooke et al. 1999). Birds have especially high temporal resolution measured as flicker fusion frequency (FFF) extending up to 100 Hz in domestic fowl (Lisney et al. 2011), and varying with light levels (average FFF: 20–87 Hz), underlining the visual adaptation to a highly mobile life, where fast aerial movements through forested habitats and detection of predators have been important selective forces involved in shaping the form and function of the birds’ visual system.
Birds have been shown to possess extraordinary colour vision (Bowmaker 1980; Goldsmith 1991; Finger and Burkhardt 1994), involving at least four classes of visual cone pigments with specific maximum sensitivity to different parts of the spectrum and an additional long-wavelength-sensitive double cone (Bowmaker et al. 1997; Bowmaker 1991; Hart et al. 1998). Associated with the single and double cones in birds, a number of oil droplets have been described. Absorbing in different parts of the 300–700 nm range, they function as cut-off filters (e.g. Bowmaker 1977; Lythgoe 1979; Partridge 1989; Hart et al. 1998, 1999, 2000; Beason and Loew 2008). The colourless oil droplets, on the other hand, seem to have a light-gathering function (Baylor and Fettiplace 1975). It has been found that a certain type of oil droplets is associated with one type of cone pigments (Bowmaker 1991, 2008), and most of them contain carotenoid pigments (Wald and Zussman 1937; Goldsmith et al. 1984). The dimensions of oil droplets for pigeons are typically 2–4 μm (Muntz 1972). The combination of cones and oil droplets provides an arsenal of different types of optical sensors which can be important in colour and hue discrimination as well as possibly in polarisation vision (Fig. 12.1). The colour vision in birds is used in a number of tasks including mate choice (e.g. Andersson and Amundsen 1997; Hunt et al. 1998), foraging (e.g. Church et al. 1998) and orientation (Able 1982, 1989), where detection of the pattern of skylight polarisation and linearly polarised light may be one of these tasks. Still we need to understand the mechanism behind polarisation vision in birds.

Schematic diagram of photoreceptors found in the avian retina of diurnal songbirds. Below the schematic photoreceptors, the λmax of the visual pigments of the four spectral classes of single cone (SWS1, SWS2, RH2, LWS), double cones (LWS) and rods (RH1) is shown. In the UV/UVS cone class (SWS1) the maximal sensitivity is either in the UV close to 370 nm or in the violet between 400 and 420 nm. Both members of the double cones contain the same LWS (long-wavelength-sensitive) pigment as the R-type single cones. The oil droplets associated with the photoreceptors are pale (principal: P) in double cones and Red (R), Yellow (Y), Clear (C) and Transparent (T) in single cones. Given below each cone and double cone are the approximate relative percentages of the different cone types in the bird’s retina [from Bowmaker (2008)]. Visualised are “red” LWS, “green” RH2, “blue” SWS2 (short-wavelength-sensitive) and “violet or ultraviolet” SWS1 single cones. Double cones contain a red-sensitive LWS cone pigment in both members, with the principal member containing a large pale yellow, P-type droplet that cuts off at about 460 nm and the accessory member having low concentration of carotenoids that may or may not be contained in a small droplet [modified after Bowmaker (2008)]
The visible spectrum typically extends into the ultraviolet (UV) range (300–400 nm) in birds (Fig. 12.2), while this UV spectral part is invisible to humans (Burkhardt 1982; Bennett and Cuthill 1994). Within this range of vision, maximum sensitivity has been found at 470 nm for 15 bird species (Chen et al. 1984; Chen and Goldsmith 1986) and at 480 nm for the red-billed leiothrix (Leiothrix lutea; Burkhardt and Maier 1989; Maier 1992; Maier and Bowmaker 1993). In the UV range (<400 nm), birds have five times higher sensitivity compared to the human visual range (400–700 nm), and they can discriminate between nearby wavelengths differing by only 7–16 nm (Emmerton and Delius 1980). It is, thus, most likely that here we will find some of the most specialised adaptations and important functions within the most sensitive range of the avian visual system. However, so for experimentation studying, for example, the polarisation sensitivity in birds has mainly been concentrated outside this range of the visible spectrum (400–700 nm), since the polarisers used in the behavioural experiments absorb strongly or completely in the UV (<400 nm; Horváth and Varjú 2004, pp. 328–354).

Light intensity during the twilight period (measured as illuminance in lux on a horizontal surface) at different elevations of the sun. Given are sunset (highlighted by a vertical dotted line) and ends of civil, nautical and astronomical twilight periods [from Åkesson et al. (1998); modified after Dusenbery (1992), see also Rozenberg (1966)]
Birds have been indirectly shown to be able to perceive linear polarisation in behavioural experiments studying avian orientation (Able and Able 1990, 1993; Muheim et al. 2006a), but the photoreceptors and specific mechanism(s) involved in this process in the bird eyes are still not described (Horváth and Varjú 2004, pp. 328–354). Behavioural evidence has further provided conflicting results, which, in part, can be attributed to erroneous experimental design and lack of complete control of the manipulated cues (Horváth and Varjú 2004, pp. 328–354, and references therein). Differences may also be explained by species-specific adaptations in the avian visual system and differential use of cues for orientation between bird species. In this chapter I intend to review the most recent findings of polarisation vision and associated behaviours in birds, which have mainly been published after the in-depth review by Horváth and Varjú (2004, pp. 328–354), and put the avian polarisation vision in an ecological perspective.
2 Polarisation Vision in Birds
A proposed mechanism for avian polarisation vision has been suggested to involve a double cone (Young and Martin 1984; Cameron and Pugh 1991) with an associated transparent oil droplet, the optical functional characteristics of which have been extensively outlined by Horváth and Varjú (2004, pp. 328–354). In this mechanism the oil droplet significantly enhances the photon capture rate in the outer segment of the photoreceptor. The lack of melanin as a screening pigment in the outer segments of the double cones further facilitates that light may pass from one cell to the one nearby. The mechanism would allow light to scatter when passing the oil droplet located in the principal cone and then to pass the outer segment of the associated cone sideways. The polarisation characteristics of light passing the oil droplet will retain those of the incoming light, and since the outer segment of vertebrate photoreceptors is dichroic when illuminated from the side (Hárosi and MacNichol 1974), the accessory cone signals would have the inherited characteristics to vary with the E-vector orientation of scattered light, oriented parallel or perpendicular to the disc membranes as was pointed out by Horváth and Varjú (2004, pp. 328–354). If this promising mechanism is indeed used in the avian visual system for detecting light polarisation, however, still remains to be shown.
3 Celestial Orientation and Migration
Migratory birds have adapted their phenotype to cope with long migration flights, fuelling and orientation (Åkesson and Hedenström 2007). Three biological compasses have been described for birds, based on information from (1) the geomagnetic field (Wiltschko and Wiltschko 1972, 1995), (2) the stars (Emlen 1967a, b, 1970, 1975) and (3) the sun and the related pattern of skylight polarisation (Wiltschko 1980, 1981; Able 1982, 1989; Schmidt-Koenig 1990). The linearly polarised skylight is expected to be used for compass orientation as the E-vector of first-order-scattered skylight is always perpendicular to the scattering plane determined by the sun, the observer and the celestial point observed (Chaps. 17 and 18). The pattern of skylight polarisation is, thus in theory, providing birds with an additional cue to pinpoint the position of the sun when the sun itself may be occluded by clouds or landmarks or is below the horizon (Horváth and Varjú 2004). In insects and spiders the polarisation pattern of the sky may provide the most important compass used during navigation (von Frisch 1949; Wehner 1989), and polarised skylight may be used also to guide certain dung beetles during dusk orientation (Chap. 2).
The avian magnetic compass is based on the angle of inclination and not on the polarity of the Earth’s magnetic field (Wiltschko and Wiltschko 1972). The magnetic field is most likely perceived by specialised photoreceptors in the bird’s eyes, involving cryptochrome molecules and a radical-pair process (for review, see Mouritsen and Hore 2012). In a neuroanatomical study, Niessner et al. (2011) discovered bands of cryptochrome 1a molecules along the membrane discs of the outer segments of UV/violet cones in the retinas of the domestic chicken (Gallus gallus) and the European robin (Erithacus rubecula), a migratory songbird. The discovery is suggestive and supports the possible involvement of these visual receptor cells in avian magnetoreception. Furthermore, the sensitivity of the magnetic compass in birds is tuned to the ambient magnetic field (25,000–68,000 nT) expected to be met during migration, but may be modified also outside the range of intensities occurring at different regions of the Earth (15,000 and 150,000 nT, respectively; for review see Wiltschko and Wiltschko 2010). It will be interesting to find out if the mechanism for polarisation vision in birds, possibly involving the double cones (Young and Martin 1984), is somehow integrated in the retina with the magneto-sensitive photoreceptors.
A population-specific orientation relative to the geomagnetic field has been shown to be inherited in songbirds, in which experience from a combination of the geomagnetic field and the rotational centre of the night sky during development is crucial (Weindler et al. 1996). In addition, the necessary 180° shift in relation to the angle of inclination for two bird species, the garden warbler, Sylvia borin, and the bobolink, Dolichonyx oryzivorus, crossing the magnetic equator on migration has been demonstrated (Beason 1992; Wiltschko and Wiltschko 1992). The stellar compass is also inherited, but for functionality a rotating stellar sky needs to be experienced during development (Emlen 1970, 1975). The stellar compass provides birds with a direction relative to the geographic North, which is given by the rotation centre and may be memorised based on stellar patterns. This compass does not seem to be connected to an internal time sense in birds as the sun compass, which completely depends on this internal time-keeping mechanism in order to work (Schmidt-Koenig 1990). The sun compass in homing pigeons (Columba livia) further seems to be finely tuned to local azimuthal movement of the sun during the day (Wiltschko et al. 2000).
During long-distance flights the time-compensated sun compass has been suggested to be the major compass mechanism at work (Alerstam et al. 2001). At these long flights across longitudes, the bird’s internal time sense is thought not to be adjusted to local time during flights, but remains at the local time from the site where the bird departed (Alerstam et al. 2001). When applying such a mechanism, the flight trajectories will lead the birds along great circle routes, i.e. shortest-distance routes between two points on the globe. This hypothesis has rendered some support from the long flights by arctic waders flying along approximate great circle routes in autumn from the high Arctic to wintering sites in the southern hemisphere (Alerstam et al. 2001). However, not all migrating birds are following such routes between start and end positions during extensive migration flights (e.g. Gudmundsson et al. 1995; Åkesson et al. 2012), and it is still open how these deviating routes may be explained and what compass mechanism(s) may be used during these flights.
The majority of songbirds migrate at night, and these nocturnal migrations involve impressive numbers of birds crossing continents and seas. For instance, the autumn avian migration between Europe and Africa has been estimated to involve more than 2.6 billion birds (Haan et al. 2009). For successful migration, the birds rely on the functionality of their biological compasses in flight, but they need also to be able to pinpoint their species-specific and endogenously encoded geographical goal by additional cues. Nocturnal songbird migrants have been shown to predominantly initiate their nocturnal flights shortly after sunset (Fig. 12.3; e.g. Åkesson et al. 1996, 2001), but it has also been shown that the decision to start migration at night may be triggered by the visibility of a clear night sky (Åkesson et al. 1996). In addition to the visibility of celestial cues, also the wind situation will be important for departure decision (Åkesson and Hedenström 2000).

Time of departure flights where individual songbirds initiated migration during the twilight and night period in south Sweden. Given is the number of registrations of birds departing on nocturnal migration flights in relation to sun elevation at departure. Sun elevations at Sunset, Civil (CT), Nautical (NT) and Astronomical Twilights (AT) are given by vertical dotted lines. Light grey refers to flocks or single birds recorded by radar and dark grey to radio-tracked birds [from Åkesson et al. (1998)]
For songbirds the time of transition between day and night is critical for migration departure, and at this time migratory songbirds have been shown to climb into trees and bushes and to sit quietly observing the surroundings (Palmgren 1949), probably observing the sky conditions, compass cues as well as weather situations before migration departures. Thus, the orientation information available during the twilight period is most likely of great importance for the departing songbirds (Emlen 1975; Moore 1987; Åkesson et al. 1996), and one or several of the available cues will further provide guidance during the upcoming nocturnal migration flight.
4 Importance of the Twilight Period for Orientation
The twilight period involves the entire complex of atmospheric optical phenomena that takes place when the sun is near the horizon (from +6° to −18°) and when the transition between daylight illumination and darkness occurs (Rozenberg 1966; Fig. 12.4). At the twilight period, the celestial cues related to the sun, i.e. azimuthal position of the sun, horizon glow and colour gradients and the pattern of skylight polarisation, are clearly visible (Rozenberg 1966; Brines and Gould 1982; Cronin et al. 2006), and in the end of the civil twilight period (when the sun reaches below −6°) the first stars become visible. Furthermore, at sun elevations near the horizon the highest (up to 80 %) degrees of polarisation of skylight occur. The twilight provides the possibility to put different orientation cues into conformity by their coordinated presence, and based on this, the departing bird may decide on the particular course to select for the upcoming nocturnal flight (Åkesson et al. 1996). A similarly important period occurs at sunrise, when birds migrating predominantly at daytime initiate their migrations.

Schematic flight altitude profile based on a reflectivity profile collected from a Doppler weather radar located at De Bilt in The Netherlands and visualised for the night of 12 June 2009, showing twilight ascents of common swifts Apus apus. The altitude profile in grey shows that the swifts are engaged in both dusk and dawn ascent flights to similarly high altitudes of 2,500 and 2,000 m at both times. The displayed time series started at sunset and ended at sunrise. Solid vertical black lines indicate the transition between civil twilight and nautical twilight, when the sun has reached −6° below the horizon. Dotted vertical lines indicate the transition between night and nautical twilight [sun elevation −12°; modified after Dokter et al. (2013)]
It has recently been shown (Dokter et al. 2013) that the highly aerial common swift, Apus apus, makes ascent flights to high altitudes at times symmetrical to the time of local sunset and sunrise, when the sun shows the same elevation relative to the horizon at both times (Fig. 12.5). These high-altitude ascent flights have been demonstrated by weather Doppler radar for common swifts spending the night on the wings during the summer months and may extend up to 2,000–2,500 m altitudes (Dokter et al. 2013). The highest altitude was reached at both sunset and sunrise at solar elevations ranging between −6° and −8° relative to the horizon, when the maximum degree of polarisation of the twilight sky is visible, horizon glow (i.e. radiance and spectral gradients) is present and stars are also visible in the sky.

Maximum ascent flight altitude and solar elevation at the time when the common swifts reached the maximum altitude during (a) dusk ascent and (b) dawn ascent. Data are given for days with limited cloud cover. The solid lines show the linear least-square regressions [from Dokter et al. (2013)]
The occurrence of the two symmetric twilight high-altitude flights was unexpected and could not be explained by the presence of foraging insects, because the birds were aiming for altitudes above those where insects were present at night, and also the swifts reached higher altitudes than other bird species present in the same region at night (Dokter et al. 2013). The authors therefore concluded that the flights may instead be explained by other factors. One previously suggested possibility is that the common swifts may climb to high altitudes for nocturnal roosting (Weitnauer 1952; Bruderer and Weitnauer 1972; Tarburton and Kaiser 2001). However, the symmetrical climbing flights occurring in early morning could not be explained by this, since the swifts were shortly after reaching the highest altitude initiating slow descending flights to lower altitudes where they forage in daytime. Dokter et al. (2013) suggested that the ascent flights in early morning may be related to the probing for flight and foraging conditions during upcoming daytime, and/or facilitation to detect visual information for navigation such as distant landmarks and celestial information, in which the skylight polarisation pattern may be one cue of importance. Detection of the skylight polarisation pattern at nearby sunset and sunrise times has further been proposed to be important for compass calibration (Muheim et al. 2006a).
Symmetrical spike dives to deep waters in relation to sunset and sunrise times have further been observed for southern bluefin tunas (Thunnus maccoyii) in the ocean, at similar times as the swifts, and one possible explanation to these deep dives has been the relatively smooth temporal variations of the geomagnetic field at this time of the day, and thus facilitation to detect orientation cues such as the geomagnetic field (Willis et al. 2009). These two observations of the symmetrically timed twilight ascent in common swifts and spike dives in southern bluefin tunas call for further attention and suggest that the behaviour might be present also in other aerial species or long-distance migrating animals in the ocean, and may possibly be explained by the reading of orientation cues present in this time period, when in particular the pattern of skylight polarisation is especially pronounced due to the highest degrees of polarisation, and the temporal variations of the geomagnetic field are relatively smooth.
5 Importance of Skylight Polarisation Pattern in Compass Calibration
The functional characteristics of the biological compasses used by birds have been described in some detail, and the compasses based on celestial and geomagnetic information have been shown to be present in a number of songbird species. However, the integration between compass cues is much less understood, and cue-conflict experiments have rendered varying results (Åkesson 1994; Muheim et al. 2006a). Both the celestial and magnetic compasses may be recalibrated if their directional information is set into conflict (Able and Able 1990, 1993, 1995), and it is not completely clear why one or the other of the compasses is recalibrated (Åkesson 1994; Muheim et al. 2006a). In some studies performed with birds before migration, the birds seem to give precedence to celestial information and recalibrate the magnetic compass, while in other studies performed during migration the magnetic compass is used to instead recalibrate the celestial compass (Muheim et al. 2006a). Recalibration of compasses may be adaptive in songbirds, especially at high latitudes, where the magnetic declination (i.e. angular difference between geomagnetic and geographic North) may largely vary between nearby sites (Åkesson et al. 2002). Birds inhabiting these regions have to cope with variations in declination during their migrations. Free-flying migratory songbirds studied by radio-telemetry in North America have been shown to recalibrate their magnetic compass daily based on celestial twilight cues (Cochran et al. 2004). This suggests the importance of twilight sunset cues as a reference and possible involvement of the pattern of skylight polarisation in the recalibration process.
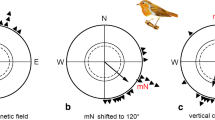
In a study (Muheim et al. 2006b) performed with Savannah sparrows (Passerculus sandwichensis) in late summer and during autumn migration, the skylight polarisation pattern near the horizon at sunset and sunrise has been shown to be crucial for recalibration of the magnetic compass (Fig. 12.6). The experiments were performed in circular cages with young and adult P. sandwichensis caught at the site of experiments in south-western Alaska. The experimental birds were held indoors without access to celestial cues, within the local geomagnetic field, and with possibility to experience the natural photoperiod. The orientation cage experiments were performed indoors in the local geomagnetic field at sunset and after, but without access to visual cues. Prior to the registration of the preferred orientation, the experimental birds had experienced a cue-conflict situation for one hour at sunrise or sunset, in which the celestial polarisation pattern was shifted ±90° relative to the pattern available at respective time of the day. Some birds were exposed to a cue conflict both at sunrise and sunset, and thereafter tested in circular Emlen cages. At exposure to the cue conflict, the Savannah sparrows were given access to the surrounding sky down to the horizon.

The magnetic orientation of Savannah sparrows exposed to a polarisation pattern shifted ±90° at sunrise (left) and sunset (right) has been measured in circular cages for birds migrating in Alaska in autumn. (a and c) 360° view of sky under natural and experimental conditions (gN: geographic north, mN: magnetic north). Black and grey bars indicate mean position of the band of maximum polarisation (BMP) at sunrise and sunset, respectively; hatched central zones indicate which areas of sky were not visible to the birds during exposure. (a) Outline of the natural relationship between sunrise/sunset celestial cues and the direction to magnetic North according to the local geomagnetic field where the experiments were performed. (b) Mean orientation of Savannah sparrows selected for exposure to cue conflict. (c) Alignment of ±90°-shifted polarisation axis using filters during exposure to cue conflict. (b and d) Magnetic orientation of birds tested indoors, plotted relative to mN and the individually preferred orientation prior to cue-conflict exposure (=0°). Savannah sparrows for which the disc of the Sun was visible during exposure are depicted by open symbols. Arrowheads show mean angle or axis of orientation. The length of the vector is a measure of concentration, being inversely related to scatter and drawn relative to the radius of the circle (r = 0 − 1). (d) Bird orientation relative to the magnetic field after cue-conflict exposure. (e) Angular deviations from each individual’s initial orientation before exposure [initial direction (b) of each individual set to =0°]. Arrows show mean axial distribution; dashed lines denote 95 % confidence intervals according to Batschelet (1981); triangles outside the circles give predicted responses for a ±90° shift in band of maximum polarisation relative to the natural sunrise (black) or sunset (grey) position [from Muheim et al. (2006b)]
The experimental birds that have previously expressed meaningful migratory directions towards south-east relative to the magnetic field were shown to recalibrate their magnetic course on the basis of the shifted celestial polarisation pattern (Muheim et al. 2006b). It became clear from these experiments that the Savannah sparrows had paid extra attention to the polarisation pattern near the horizon for these recalibrations, and that there were no differences in response between birds with or without previous experience from migration or tested at different times of the autumn period. Also in this part of the sky is the highest degree of polarisation under foggy and partly or completely cloudy conditions (Hegedüs et al. 2007a, b).
These cue-conflict experiments with Savannah sparrows were performed at high latitudes in the Arctic (Muheim et al. 2006b, 2007), but they have been proven difficult to repeat with other species of migratory birds and in other geographical areas (Muheim et al. 2008; Wiltschko et al. 2008; Gaggini et al. 2010; Schmaljohann et al. 2013), except in one species of songbird, the white-throated sparrow (Zonotrichia albicollis), studied during spring and autumn migration at a location in the transition zone between the breeding and wintering areas in North America (Muheim et al. 2009). Just like the Savannah sparrows described above, the white-throated sparrows were recalibrating their magnetic compass based on the pattern of skylight polarisation at both sunrise and sunset (Muheim et al. 2009). In the latter experiments, both the magnetic field and the polarised skylight pattern were manipulated to set the two cues into conflict, suggesting that the polarisation filters did not cause an unnatural situation to the birds (Muheim et al. 2009), which has previously been proposed. In other studies, however, the experimental birds that were given similar access to horizontal skylight cues have not been shown to recalibrate their magnetic compass on the basis of the celestial information from the pattern of skylight polarisation (Wiltschko et al. 2008; Gaggini et al. 2010; Schmaljohann et al. 2013). All of these non-responding cage experiments have been performed with migratory songbirds in Australia (Wiltschko et al. 2008) and Europe (Gaggini et al. 2010; Schmaljohann et al. 2013; Åkesson et al. unpublished), and one possible explanation for the discrepancies in the results may be local adaptation in migratory birds to the geometry of the geomagnetic and celestial compass information present in different geographical areas and to which area specific birds have evolved to migrate. However, this possibility needs to be investigated further.
6 Behavioural Evidence for Polarisation Vision in Birds
Conditioning experiments have been used to study the perception of polarisation in birds, and the first experiments were performed with homing pigeons, C. livia (Kreithen and Keeton 1974; Delius et al. 1976). Montgomery and Heinemann (1952) had previously investigated the response in homing pigeons to linearly polarised light by conditioning to a two-choice situation with a polarisation pattern orthogonally rotated relative to each other, and found no response to this manipulation, but a response to difference in brightness. Later, it has turned out to be very difficult to train birds to respond to artificial polarised stimuli (Coemans et al. 1990, 1994; Vos et al. 1995; Greenwood et al. 2003), and it has been pointed out that the difficulty may be related to optical artefacts in the experimental set-up and the difficulty to create an experimental situation where birds may be responding naturally to artificial manipulations (Horváth and Varjú 2004; Muheim 2011). The negative results of conditioning experiments designed to study polarisation sensitivity in birds have extensively been covered by Horváth and Varjú (2004, pp. 328–354).
7 Outlook
Despite the fact that birds are highly visual animals, we still lack crucial understanding of how their visual system is adapted to perceive light polarisation. A promising physiological mechanism in the bird’s eye involving a double cone has been proposed as the key mechanism in polarisation detection, but it still needs to be investigated if this is the case. It has further been hard to condition birds to polarisation detection tasks, and it might at least partly be explained by the difficulty to create a relevant artificial experimental situation where this can be tested by birds held in captivity. The twilight period involving the complete transition between day- and night-time, when the highest degree of polarisation is available in the sky, is central for migratory birds as it plays a key role in timing of migratory departures and selection of migration courses. Birds have access to several compasses and two celestial compasses based on information from the sun (and the related pattern of sky polarisation), and the stars are combined with a magnetic compass. The three compass mechanisms guide birds during flights across continents and seas, but still we lack understanding on which compass is used by migrants during the flights. Contradictory experiments with songbirds suggest a different compass hierarchy between bird species, and therefore this area needs further experimentation to fully understand the pattern observed. Because of several still unanswered questions, we should look forward to exciting discoveries on avian polarisation vision during the coming years.
References
Able KP (1982) Skylight polarization patterns at dusk influence migratory orientation in birds. Nature 299:550–551
Able KP (1989) Skylight polarization patterns and the orientation of migratory birds. J Exp Biol 141:241–256
Able KP, Able MA (1990) Ontogeny of migratory orientation in the Savannah sparrow, Passerculus sandwichensis: calibration of the magnetic compass. Anim Behav 39:905–913
Able KP, Able MA (1993) Daytime calibration of magnetic orientation in a migratory bird requires a view of skylight polarization. Nature 364:523–525
Able KP, Able MA (1995) Interactions in the flexible orientation system of a migratory bird. Nature 375:230–232
Åkesson S (1994) Comparative orientation experiments with different species of long-distance migrants: effect of magnetic field manipulation. Anim Behav 48:1379–1393
Åkesson S, Hedenström A (2000) Selective flight departure in passerine nocturnal migrants. Behav Ecol Sociobiol 47:140–144
Åkesson S, Hedenström A (2007) How migrants get there: migratory performance and orientation. Bioscience 57:123–133
Åkesson S, Alerstam T, Hedenström A (1996) Flight initiation of nocturnal passerine migrants in relation to celestial orientation conditions at twilight. J Avian Biol 27:95–102
Åkesson S, Walinder G, Karlsson L, Ehnbom S (2001) Reed warbler orientation: Initiation of nocturnal migratory flights in relation to visibility of celestial cues at dusk. Anim Behav 61:181–189
Åkesson S, Morin J, Muheim R, Ottosson U (2002) Avian orientation: effects of cue-conflict experiments with young migratory songbirds in the high Arctic. Anim Behav 64:469–475
Åkesson S, Klaassen R, Holmgren J, Fox JW, Hedenström A (2012) Migration routes and strategies in a highly aerial migrant, the common swift Apus apus, revealed by light-level geolocators. PLoS One 7(7):e41195
Alerstam T, Gudmundsson GA, Green M, Hedenström A (2001) Migration along orthodromic sun compass routes by arctic birds. Science 291:300–303
Andersson S, Amundsen T (1997) Ultraviolet colour vision and ornamentation in bluethroats. Proc R Soc Lond B 264:1587–1591
Batschelet E (1981) Circular statistics in biology. Academic, New York
Baylor ER, Fettiplace D (1975) Light path and photon capture in turtle photoreceptors. J Physiol 248:433–464
Beason RC (1992) You can get there from here: responses to simulated magnetic equator crossing by the Bobolink (Dolichonyx oryzivorus). Ethology 91:75–80
Beason RC, Loew ER (2008) Visual pigment and oil droplet characteristics of the bobolink (Dolichonyx oryzivorus), a new world migratory bird. Vis Res 48:1–8
Bennett ATD, Cuthill IC (1994) Ultraviolet vision in birds: what is its function? Vis Res 34:1471–1478
Bowmaker JK (1977) The visual pigments, oil droplets and spectral sensitivity of the pigeon. Vis Res 17:1129–1138
Bowmaker JK (1980) Colour vision in birds and the role of oil droplets. Trends Neurosci 3:196–199
Bowmaker JK (1991) Photoreceptors, photopigments and oil droplets. In: Gouras P (ed) Vision and visual dysfunction, vol 6, The perception of colour. Macmillan, London, pp 108–127
Bowmaker JK (2008) Evolution of vertebrate visual pigments. Vis Res 48:2022–2041
Bowmaker JK, Heath LA, Wilkie SE, Hunt DM (1997) Visual pigments and oil droplets from six classes of photoreceptor in the retinas of birds. Vis Res 37:2183–2194
Brines M, Gould J (1982) Skylight polarization patterns and animal orientation. J Exp Biol 96:69–91
Bruderer B, Weitnauer E (1972) Radarbeobachtungen über Zug und Nachtflüge des Mauerseglers (Apus apus). Rev Suisse Zool 79:1190–1200
Burkhardt D (1982) Birds, berries and UV. Naturwissenschaften 69:153–157
Burkhardt D, Maier EJ (1989) The spectral sensitivity of a Passerine bird is highest in the UV. Naturwissenschaften 76:82–83
Burton RF (2008) The scaling of eye size in adult birds: relationship to brain, head and body sizes. Vis Res 48:2345–2351
Cameron DA, Pugh EN (1991) Double cones as a basis for a new type of polarisation vision in vertebrates. Nature 353:161–164
Chen DM, Goldsmith TH (1986) Four spectral classes of cone in the retinas of birds. J Comp Physiol A 159:473–479
Chen DM, Collins JS, Goldsmith TH (1984) The ultraviolet receptor of bird retinas. Science 225:337–340
Church SC, Bennett AT, Cuthill IC, Partridge JC (1998) Ultraviolet cues affect the foraging behaviour of blue tits. Proc R Soc Lond B 265:1509–1514
Cochran WW, Mouritsen H, Wikelski M (2004) Migrating songbirds recalibrate their magnetic compass daily from twilight cues. Science 304:405–408
Coemans MAJM, Vos Hzn JJ, Nuboer JFW (1990) No evidence for polarization sensitivity in the pigeon. Naturwissenschaften 77:138–142
Coemans MAJM, Vos Hzn JJ, Nuboer JFW (1994) The orientation of the e-vector of linearly polarized light does not affect the behaviour of the pigeon, Columba livia. J Exp Biol 191:107–123
Cronin TW, Warrant EJ, Greiner B (2006) Celestial polarization patterns during twilight. Appl Opt 45:5582–5589
de Brooke ML, Hanley S, Laughlin SB (1999) The scaling of eye size with body mass in birds. Proc R Soc B 266:405–412
Delius JD, Perchard RJ, Emmerton J (1976) Polarized light discrimination by pigeons and an electroretinographic correlate. J Comp Physiol Psychol 90:560–571
Dokter AM, Åkesson S, Beekhuis H, Bouten W, Buurma L, van Gasteren H, Holleman I (2013) Twilight ascents by common swifts, Apus apus, at dawn and dusk: acquisition of orientation cues? Anim Behav 85:545–552
Emlen ST (1967a) Migratory orientation in the indigo bunting, Passerina cyanea. Part II: mechanism of celestial orientation. Auk 84:463–489
Emlen ST (1967b) Migratory orientation in the indigo bunting, Passerina cyanea. Part I: evidence for use of celestial cues. Auk 84:309–342
Emlen ST (1970) Celestial rotation: its importance in the development of migratory orientation. Science 170:1198–1201
Emlen ST (1975) Migration: orientation and navigation. Avian Biol 5:129–219
Emmerton J, Delius JD (1980) Wavelength discrimination in the “visible” and UV spectrum by pigeons. J Comp Physiol A 141:47–52
Finger E, Burkhardt D (1994) Biological aspects of bird colouration and avian colour vision including ultraviolet range. Vis Res 34:1509–1514
Gaggini V, Baldaccini N, Spina F, Giunchi D (2010) Orientation of the pied flycatcher Ficedula hypoleuca cue-conflict experiments during spring migration. Behav Ecol Sociobiol 64:1333–1342
Goldsmith TH (1991) Optimization, constraint and history in the evolution of eyes. Q Rev Biol 65:281–322
Goldsmith TH, Collins JS, Licht S (1984) The cone oil droplets of avian retinas. Vis Res 24:1661–1671
Greenwood VJ, Smith EL, Church SC, Partridge JC (2003) Behavioural investigation of polarisation sensitivity in the Japanese quail (Coturnix coturnix japonica) and the European starling (Sturnus vulgaris). J Exp Biol 206:3201–3210
Gudmundsson GA, Alerstam T, Benvenuti S, Papi F, Lilliendahl K, Åkesson S (1995) Examining the limits of flight and orientation performance: satellite tracking of brent geese migrating across the Greenland ice-cap. Proc R Soc Lond B 261:73–79
Haan S, Bauer S, Liechti F (2009) The natural link between Europe and Africa—2.1 billion birds on migration. Oikos 118:624–626
Hárosi FI, MacNichol EF Jr (1974) Visual pigments of goldfish cones: spectral properties and dichroism. J Gen Physiol 63:279–304
Hart NS, Partridge JC, Cuthill IC (1998) Visual pigments, oil droplets and cone photoreceptor distribution in the European starling (Sturnus vulgaris). J Exp Biol 201:1433–1446
Hart NS, Partridge JC, Cuthill IC (1999) Visual pigments, cone oil droplets, ocular media and predicted spectral sensitivity in the domestic turkey (Meleagris gallopavo). Vis Res 39:3321–3328
Hart NS, Partridge JC, Cuthill IC, Bennett AT (2000) Visual pigments, oil droplets, ocular media and cone photoreceptor distribution in two species of passerine bird: the blue tit (Parus caeruleus L.) and the blackbird (Turdus merula L.). J Comp Physiol A 186:375–387
Hegedüs R, Åkesson S, Wehner R, Horváth G (2007a) Could Vikings have navigated under foggy and cloudy conditions by skylight polarization? On the atmospheric optical prerequisites of polarimetric Viking navigation under foggy and cloudy skies. Proc R Soc A 463:1081–1095
Hegedüs R, Åkesson S, Horváth G (2007b) Polarization patterns of thick clouds: overcast skies have distribution of the angle of polarization similar to that of clear skies. J Opt Soc Am A 24:2347–2356
Horváth G, Varjú D (2004) Polarized light in animal vision: polarization patterns in nature. Springer, Heidelberg
Hunt S, Bennett AT, Cuthill IC, Griffiths R (1998) Blue tits are ultraviolet tits. Proc R Soc Lond B 265:451–455
Kreithen ML, Keeton WT (1974) Detection of polarized light by the homing pigeon, Columba livia. J Comp Physiol A 89:83–92
Lisney TJ, Rubene D, Rózsa J, Lovlie H, Håstad O, Ödeen A (2011) Behavioural assessment of flicker fusion frequency in chicken Gallus gallus domesticus. Vis Res 51:1324–1332
Lythgoe JN (1979) The ecology of vision. Oxford University Press, Oxford
Maier EJ (1992) Spectral sensitivities including the UV of the passeriform bird Leiothrix lutea. J Comp Physiol A 170:709–714
Maier EJ, Bowmaker JK (1993) Colour vision in the passeriform bird, Leiothrix lutea: correlation of visual pigment absorbance and oil droplet transmission with spectral sensitivity. J Comp Physiol A 172:295–301
Martin GR (1985) Eye. In: King AS, McLelland J (eds) Form and function in birds, vol 3. Academic, London, pp 311–373
Montgomery KC, Heinemann EG (1952) Concerning the ability of homing pigeons to discriminate patterns of polarized light. Science 116:454–456
Moore FR (1987) Sunset and the orientation behaviour of migrating birds. Biol Rev 62:65–86
Mouritsen H, Hore P (2012) The magnetic retina: light-dependent and trigeminal magnetoreception in migratory birds. Curr Opin Neurobiol 22:343–352
Muheim R (2011) Behavioural and physiological mechanisms of polarized light sensitivity in birds. Philos Trans R Soc B 366:763–771
Muheim R, Moore FR, Phillips JB (2006a) Calibration of magnetic and celestial compass cues in migratory birds: a review of cue-conflict experiments. J Exp Biol 209:2–17
Muheim R, Phillips JB, Åkesson S (2006b) Polarized light cues underlie compass calibration in migratory songbirds. Science 313:837–839
Muheim R, Åkesson S, Phillips JB (2007) Magnetic compass of migratory Savannah sparrows is calibrated by skylight polarization at sunrise and sunset. J Ornithol 148:485–494
Muheim R, Åkesson S, Phillips JB (2008) Response to R. Wiltschko et al. (Journal für Ornithologie): contradictory results on the role of polarized light in compass calibration in migratory songbirds. J Ornithol 149:659–662
Muheim R, Phillips JB, Deutschlander ME (2009) White-throated sparrows calibrate their magnetic compass by polarized light cues during both autumn and spring migration. J Exp Biol 212:3466–3472
Muntz WRA (1972) Inert absorbing and reflecting, pigments. In: Dartnall HJA (ed) The handbook of sensory physiology, vol WII/l. Springer, Berlin, pp 529–565
Niessner C, Denzau S, Gross JC, Peichl L, Bischof HJ, Fleissner G, Wiltschko W, Wiltschko R (2011) Avian ultraviolet/violet cones identified as probable magnetoreceptors. PLoS One 6:e20091
Palmgren P (1949) On the diurnal rhythm of activity and rest in birds. Ibis 91:561–576
Partridge JC (1989) The visual ecology of avian cone oil droplets. J Comp Physiol A 165:415–426
Rozenberg GV (1966) Twilight. A study in atmospheric optics. Plenum, New York
Schmaljohann H, Rautenberg T, Muheim R, Naef-Daenzer B, Bairlein F (2013) Response of a free-flying songbird to an experimental shift of the light polarization pattern around sunset. J Exp Biol 216:1381–1387
Schmidt-Koenig K (1990) The sun compass. Experientia 46:336–342
Tarburton MK, Kaiser E (2001) Do fledgling and pre-breeding common swifts Apus apus take part in aerial roosting? An answer from a radiotracking experiment. Ibis 143:255–264
von Frisch K (1949) Die Polarisation des Himmelslichtes als orienterender Faktor bei den Tänzen der Bienen. Experientia 5:142–148
Vos HJJ, Coemans M, Nuboer J (1995) No evidence for polarization sensitivity in the pigeon electroretinogram. J Exp Biol 198:325–335
Wald G, Zussman H (1937) Carotenoids of the chicken retina. Nature 140:197
Wehner R (1989) Neurobiology of polarization vision. Trends Neurosci 12:353–359
Weindler P, Wiltschko R, Wiltschko W (1996) Magnetic information affects the stellar orientation of young bird migrants. Nature 383:158–160
Weitnauer E (1952) Übernachtet der Mauersegler, Apus apus (L.), in der Luft? Ornithologische Beobachtungen 49:37–44
Willis J, Phillips J, Muheim R, Diego-Rasilla FJ, Hobday AJ (2009) Spike dives of juvenile southern bluefin tuna (Thunnus maccoyii): a navigational role? Behav Ecol Sociobiol 64:57–68
Wiltschko R (1980) Die Sonnenorientierung der Vögel. 1. Die Rolle der Sonne im Orientierungssystem und die Funktionsweise des Sonnenkompass. J Ornithol 121:121–143
Wiltschko R (1981) Die Sonnenorientierung der Vögel. 2. Entwicklung des Sonnenkompass und sein Stellenwert im Orientierungssystem. J Ornithol 122:1–22
Wiltschko W, Wiltschko R (1972) Magnetic compass of European robins. Science 176:62–64
Wiltschko W, Wiltschko R (1992) Migratory orientation: magnetic compass orientation of garden warblers, Sylvia borin, after a simulated crossing of the magnetic equator. Ethology 91:70–74
Wiltschko R, Wiltschko W (1995) Magnetic orientation in animals. Springer, Heidelberg
Wiltschko R, Wiltschko W (2010) Avian magnetic compass: its functional properties and physical basis. Curr Zool 56:265–276
Wiltschko R, Walker M, Wiltschko W (2000) Sun-compass orientation in homing pigeons: compensation for different rates of change in azimuth? J Exp Biol 203:889–894
Wiltschko R, Munro U, Ford H, Wiltschko W (2008) Contradictory results on the role of polarized light in compass calibration in migratory songbirds. J Ornithol 149:607–614
Young SR, Martin GR (1984) Optics of retinal oil droplets: a model of light collection and polarization detection in the avian retina. Vis Res 24:129–137
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2014 Springer-Verlag Berlin Heidelberg
About this chapter
Cite this chapter
Åkesson, S. (2014). The Ecology of Polarisation Vision in Birds. In: Horváth, G. (eds) Polarized Light and Polarization Vision in Animal Sciences. Springer Series in Vision Research, vol 2. Springer, Berlin, Heidelberg. https://doi.org/10.1007/978-3-642-54718-8_12
Download citation
DOI: https://doi.org/10.1007/978-3-642-54718-8_12
Published:
Publisher Name: Springer, Berlin, Heidelberg
Print ISBN: 978-3-642-54717-1
Online ISBN: 978-3-642-54718-8
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)