
Abstract
Net ecosystem exchange of CO2 (NEE) measurement was carried out in tropical lowland paddy at ICAR-National Rice Research Institute, Cuttack, Odisha, India, in 2015 using eddy covariance technique with the objective to assess the variation of NEE of CO2 in lowland paddy and to find out the most suitable model for better partitioning of net ecosystem exchange of CO2 in tropical lowland paddy. Paddy is grown twice (dry and wet season) a year in this region in the lowland, and the field is kept fallow during the remainder of the year. Two different flux partitioning models (FPMs)—the rectangular hyperbola (RH) and the Q10, were evaluated to assess NEE of CO2, and its partitioning components—gross primary production (GPP) and ecosystem respiration (RE), and the resulting flux estimates were compared. The RH method assessed the effects of photosynthetically active radiation on the NEE, whereas the Q10 method utilized the relationship between ecosystem respiration and temperature in lowland paddy. The average NEE during the dry season and wet season was − 1.62 and − 1.83 g C m−2 d−1, respectively, whereas it varied from − 5.71 to 2.29 g C m−2 d−1 during the observation period covering both the cropping seasons and the fallow period. The mean difference between modeled GPP and RE from two FPMs was found significant in both the seasons. The maximum correlation for GPP estimation was found between two FPMs at the panicle initiation stage during both the dry season (R2 = 0.767) and wet season (R2 = 0.321). It was evident from the study that the Q10 method reliably produced the most realistic carbon flux estimates over the RH method, for the lowland paddy. The Q10 model which used nighttime flux and temperature data to estimate RE produced estimates that had lower prediction error (RMSE) as compared to the RH model. It can be concluded that in lowland paddy, the Q10 predicted better estimates of RE and GPP values than the RH method, suggesting that the Q10 model can be used for partitioning of NEE in tropical lowland paddy.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Net ecosystem exchange (NEE) measurements of carbon dioxide (CO2) are now playing a crucial role in the progress of climate change science through multiple scales with an increasing number of eddy covariance towers worldwide (Baldocchi 2008). Eddy covariance (EC) measurements of NEE are widely used globally to quantify the role of vegetation in controlling the scale and variability of the global carbon sink (Sarmiento and Wofsy, 1999; Baldocchi 2008).
Globally paddy is cultivated in about 160 M-ha, divulging a vital role in carbon cycle (Pathak et al. 2018; Chatterjee et al. 2019a, b). Paddy is grown in about 43.95 M-ha in India with an annual production of 106.54 Mt (GOI 2014). The NEE of CO2 of lowland paddy, a semiaquatic crop, can be measured over a large area using the EC technique (Chapin et al. 2006; Swain et al. 2018a, b). For assessment of the processes which control NEE in lowland paddy, EC data mostly depend on models that partition NEE into two components—gross primary production (GPP) and ecosystem respiration (RE). For understanding the partitioning of NEE with more accuracy to interpret its controls, this is imperative as the global budget for GPP is highly uncertain with the values ranging from 110 to 150 Pg-C yr−1 (Beer et al. 2010; Jung et al. 2011). Often estimates of GPP and RE data differ depending upon the choice of flux partitioning models. There are several measurements and modeling methodologies which are used for partitioning NEE, each of which has its own advantages and disadvantages (Desai et al. 2008). These flux partitioning models (FPMs) can be very useful for gap filling of missing data, thus allowing estimates of CO2 flux over long time periods (Falge et al. 2002). Hence, FPMs are important tools for inferring EC data, yet very few studies so far have critically examined how the choice of a given FPM affects the degree and variability of C-flux estimates in tropical lowland paddy.
Yearlong EC time series data from the lowland paddy in eastern India were used to generate NEE, GPP and RE estimates using two FPMs that differ significantly in the data source used (i.e., daytime versus nighttime data) and difficulty of the modeling procedure. These estimates from two models were then compared against each other to assess the magnitude and seasonal variability of NEE, RE and GPP. While doing the inter-comparison of data, it is imperative to note that model-based estimates are generally liable to various random and systematic errors and may not always denote the unknown “true” flux value. There is a substantial interest in partitioning the measured NEE of CO2 to find better perceptions into the process-level controls over NEE. During the nighttime, partitioning is simple, as RE becomes equal to NEE, whereas, during the daytime, the partitioning depends on the model used. Hence, there are considerable uncertainties related to the estimates of GPP and RE (Hagen et al. 2006; Richardson et al. 2006). The daytime RE can be extrapolated from the nighttime flux measurements using some temperature response function, but this approach does not account for daytime inhibition of foliar respiration, which is estimated to be 11–17% of GPP (Wohlfahrt et al. 2005). There is another method by which daytime RE is estimated from the y-axis intercept from a light response curve (Lasslop et al. 2010). Desai et al. (2008) reported that most of the partitioning methods varied by less than 10% in terms of yearly integrals and more variability among the methods was observed when extra gaps were included in the data. It was apparent that patterns of GPP across the locations tended to be consistent when a single partitioning algorithm was applied which indicates that choice of algorithm mostly results in asystematic bias of unknown magnitude since the “true” GPP is not known. In the case of the partitioning of RE, more variability within algorithms was observed at shorter timescales (e.g., with regard to diurnal cycles) (Lasslop et al. 2010). Soil also plays an important role in ecosystem respiration by sequestering C in labile and non-labile pools and subsequent release as CO2 which get altered with different agronomic management practices such as mulching, irrigation and fertilizer management (Chatterjee 2014; Chatterjee et al. 2016, 2017, 2018) and other organic input managements. Hence, estimates of the RE in large scale often contain errors in the data.
The current study focused on two different FPMs that have a strong basis in ecosystem physiology, particularly those that parameterize well-known relationships between respiration and temperature or between NEE and photosynthetically active radiation (PAR). The rectangular hyperbola (RH) is an established method to assess the effects of PAR on NEE. It has been reported that the flux data from the EC system at nighttime and temperature are significantly related to each other (Lee et al. 1999). On the other hand the Q10 method is very useful to model RE using temperature as a dominant factor. The Q10 method is also used for gap filling of missing flux data collected during nighttime conditions having sufficient turbulence (e.g., u* > 0.2 m s−1) by using air temperature as a key physical driver of RE (Stoy et al. 2005). These methodologies engage the simple equations used by many ecosystem models and, therefore, can produce transferable information for future studies of C-dynamics both across time and space at the lowland paddy ecosystem and other ecosystems relating with similar characteristics. Thus, it was hypothesized that NEE of CO2 varies with paddy-growing season, and there is a significant difference in estimates of GPP and RE partitioned by two different FPMs and choices of the model may influence the partitioning of NEE in tropical lowland paddy. To investigate this hypothesis, the current study was carried out with the following objectives (1) to study the variation of the net ecosystem exchange of CO2 in lowland paddy and (2) to find out the best-suited model for better partitioning of net ecosystem exchange of CO2 in lowland paddy.
Materials and methods
Study site and establishment of crop
The field experiment was conducted at the eddy covariance site of ICAR-National Rice Research Institute (20°26′ 60.0″N, 85°56′ 10.9″E, 24 m above average sea level) in Cuttack, Odisha, India, during the paddy-growing season of 2015. The climate of Cuttack is tropical humid, with wet hot summers (March to June) and brief mild winters (December to February). The annual average maximum and minimum temperatures in 2015 were 39.2 and 22.5 °C, respectively, whereas the annual average temperature was 27.7 °C. The PAR in this site varied from 112.70 to 1101.32 μmol m−2 s−1. Average annual rainfall was 1500 mm. The soil texture of the experimental site was sandy clay loam (25.9% clay, 21.6% silt and 52.5% sand) and categorized as Aeric Endoaquept in soil taxonomic classification system (Chatterjee et al. 2019a, b). The average bulk density of the study site was 1.42 Mg m−3. The measured pH (1:2.5 soil: water suspension) varied from 6.20 to 6.32, and a low average electrical conductivity (0.44 dS m−1) was recorded. Total carbon and total nitrogen of the study area varied from 11.2 to 11.4 g kg−1 and 0.78 to 0.9 g kg−1, respectively.
Paddy crop was grown in two seasons, i.e., dry season (DS) and wet seasons (WS) of 123 and 131 days, respectively, including two fallow periods, i.e., dry fallow (DF) and wet fallow (WF) of 64 and 47 days, respectively. Four crop growth stages were mostly identified which are vegetative (Veg), panicle initiation (PI), flowering (FL) and harvesting or maturity (H). Twenty-one-day-old seedlings of paddy (cultivar Naveen in DS cultivar Swarna Sub-1 in WS) were transplanted to a puddled soil with a spacing of 20 cm × 15 cm. Transplanting was done on the 2nd week of July in WS and 1st week of January in DS and harvested in November and last week of May, respectively. Nitrogen fertilizer was applied in three equal splits at basal, vegetative and panicle initiation stages. The rate of N application was 80 kg ha−1 in WS and 100 kg ha−1 in DS. Phosphorus (P) and potassium (K) were added at the rate 40 kg ha−1 each at basal during land preparation in both the seasons. During the fallow period, there was no crop left and the water was drained out before 15 days of the harvest. There was 8 cm standing water during the paddy-growing period, and before harvesting, it was drained out.
Eddy covariance instrumentation setup
The eddy covariance instrument was fitted in the lowland paddy field at its middle position covering a fetch area of 2.25 hectares. To assure a uniform height of paddy crop yearlong uniform breeder paddy variety was grown within the fetch area of the EC system. The components that comprised the eddy system were (1) a three-dimensional sonic anemometer (CSAT3, M/s Campbell Scientific Corp., Logan, Utah, USA) which measures wind speed along with three non-orthogonal sonic axes in the real time, (2) open path infrared gas analyzer (LI-7500A, M/s LICOR Inc., Canada) measuring fluctuations in CO2 and water vapor densities, (3) a temperature-humidity sensor (HMP45C, Campbell Scientific Corp., Logan, Utah, USA) measuring air temperature (Ta), relative humidity (RH), (4) a 4-component radiation sensor (CNR4, KIPP and ZONEN, Netherlands) measuring net radiation, (5) a soil temperature probe (107 B, Campbell Scientific Corp., Logan, Utah, USA) which measures soil temperature and (vi) a PAR sensor (LI190SB) which measures PAR. Both the sonic anemometer and infrared gas analyzer were mounted on a tripod aluminum mast at 1.5 m height. All the signals from different sensors were logged and stored in a data logger (CR3000, Campbell Scientific Corp., Logan, Utah, USA) at a sampling frequency of 10 Hz.
The NEE was calculated to sum up the half hourly daily CO2 flux and CO2 storage change. As paddy canopy height was relatively low, hence the storage term was neglected for the NEE calculation. The average vertical CO2 flux density (g C m−2 d−1) was calculated by the following formula (Webb et al. 1980; Baldocchi 2003):
where ρa is dry air density (kg m−3), C′ is CO2 mixing ratio, ω′ is the 30-min covariance between vertical fluctuations of wind speed (m s−1), time averaging was denoted by over bar, whereas the primes denote fluctuations of the average value. The flux symbol is negative when CO2 is assimilation by the vegetation from the atmosphere, and positive, otherwise. Average of NEE was used to compute daily and seasonal net exchange by the 30-min data representation (Massman and Lee 2002).
Processing, quality control and gap filling eddy covariance data
Raw eddy covariance flux data set has been processed for quality control and flux corrections (Mauder et al. 2006; Mauder and Foken 2011). The other corrections included are: the planar fit coordinate rotation correction (Wilczak et al. 2001), coordinate rotation (Kaimal and Finnigan 1994), translation of buoyancy into sensible heat flux (Liu et al. 2001); density fluctuations correction (“WPL” correction) (Tanner and Thurtell 1969; Webb et al. 1980). The u* filtering (Reichstein et al. 2005; Papale et al. 2006) and spike removal with linear interpolation for the detected spikes were performed (Vickers and Mahrt 1997). The threshold of frictional velocity (u*) for data during the nighttime was filtered as 0.1 m s−1 in this lowland paddy (Bhattacharyya et al. 2014). Gap filling of lost and discarded data was completed by the “look-up” table method (Falge et al. 2002).
Flux partitioning methods
Two different methods of varying complexity were explored for NEE partitioning into its components, i.e., GPP and RE. The study is mainly based on the methods that parameterize known relationships between driving meteorological parameters and NEE. One method, namely Q10 method, uses measured nighttime fluxes to predict RE as a function of air temperature. The second one is based on the rectangular hyperbolic fit (RH) which uses the intercept of the relationship between PAR and NEE of daytime to model RE. GPP was then calculated from the definition:
The units of NEE, GPP and RE is in µmol m−2 s−1
Rectangular hyperbola (RH)
The rectangular hyperbola is an established method to assess the effects of PAR on NEE. It has been reported that the flux data from the EC system at nighttime and temperature are significantly related to each other although it is largely scattered (Lee et al. 1999). They used an intercept parameter (γ) of the RH model (i.e., the Michaelis–Menten model) to examine the seasonal dynamics of RE in a way that was less bound by the limitations of nighttime EC data quality, and finally the NEE is written as (Ruimy et al. 1995):
where α denotes apparent ecosystem considerable yield, β is light saturation determined CO2 uptake rate (µmol m−2 s−1), γ signifies estimate of RE (µmol m−2 s−1), and Q denotes PAR (µmol m−2 s−1). In this study, the parameters of Eq. (3) were determined crop growth stage-wise and daily γ value was used as an estimate of RE (Table 1). Instantaneous data sets, β, γ and Q, are in µmol m−2 s−1.
Q10 approach
One of the most popular approaches to model RE using temperature as a dominant factor is the so-called Q10 equation:
where R10 denotes ecosystem base respiration at 10 °C, Ta is air temperature (°C), and Q10 describes the temperature sensitivity parameter, here delineating the amount of change in RE for a 10 °C change in temperature. This method is very useful for gap filling of missing flux data collected during nighttime conditions having sufficient turbulence (e.g., u* > 0.2 m s−1) by using air temperature as a key physical driver of RE (Stoy et al. 2005). The parameter values of R10 and Q10 were determined for each growth stage of paddy crop and season-wise for the whole year of study. It was noted that while parameterizing Eq. (4) seasonal variations in u* impact the eddy data set which is biased toward colder seasons (Gu et al. 2005; Reichstein et al. 2005).
For Q10 we built regression relationship of day-time NEE versus day-time air temperature. We considered that nighttime NEE is mostly contributed by RE in the absence of photosynthesis (GPP) during that period. Thus, we used this relationship to model day-time RE and day-time GPP as NEE = GPP − RE. The regression equations which were used for building the Q10 model are shown in Table 2.
For model validation and comparison we did an extensive literature survey for GPP and RE estimates in paddy in similar climatic conditions in different part of the world and assumed that our estimates of GPP and RE would follow similar trend. We did not have direct measurements of GPP and RE; hence, we compared with estimates of previous researchers (Ruimy et al. 1995; Bhattacharyya et al. 2013, 2014; Stoy et al. 2006; Desai et al. 2008; Wohlfahrt and Galvagno 2017; Oikawa et al. 2014, 2017; Wehr et al. 2016; Wohlfahrt and Gu 2015; Swain et al. 2016, 2018a; Vargas et al. 2011) for similar edaphoclimatic conditions.
Statistical analysis
Paired sample t test statistics for both the models of NEE partitioning was done by using statistical analyses software SPSS (version 20.0). Model root-mean-squared error (RMSE), standard deviation (SD) and coefficient of determination (R2) values were obtained from MS-Excel 2010 software.
Results
Daily, seasonal and annual variations in NEE
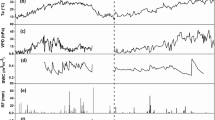
The average NEE of the lowland paddy field varied from − 5.71 to 2.29 g C m−2 d−1 during the observation period covering both the cropping seasons and the fallow period (Fig. 1). The average NEE during the DS and WS was − 1.62 and − 1.83 g C m−2 d−1, respectively. The average NEE during the fallow period after harvest of the dry season and wet season crop was − 1.49 and − 0.29 g C m−2 d−1, respectively. The average and cumulative NEE for the year 2015 was − 5.23 and − 542.47 g C m−2 d−1, respectively. Cumulative NEE for DF and WF was − 95.37 and − 14.81 g C m−2, whereas cumulative NEE values during DS and WS were − 199.73, − 232.55 g C m−2, respectively.

Variation of daily average net ecosystem exchange of CO2 during the dry and wet season (Veg vegetative, PI panicle initiation, FL flowering and H harvesting stages) and the fallow period of lowland paddy
Daily, seasonal and yearlong variations in GPP in two FPM
Throughout the entire cropping seasons (DS and WS) and fallow periods (DF and WF), the daily variation of average GPP varied from − 7.25 to − 20.80 µmol m−2 s−1 and from − 0.025 to − 22.68 µmol m−2 s−1, obtained from RH and Q10 method, respectively. The annual average values of GPP obtained from RH and Q10 method are − 16.02 and − 10.46 µmol m−2 s−1, respectively, and the statistical mean difference between GPP values estimated by these two different FPMs is significant (Table 3). During the DS the average GPP values (modeled by RH method) were − 8.20, − 10.28, − 15.81 and − 18.55 µmol m−2 s−1 in Veg, PI, FL and H stages, respectively, with a seasonal average value of − 13.37 µmol m−2 s−1. The average GPP values during DS (modeled by Q10 method) were − 1.76, − 5.12, − 10.77 and − 17.16 µmol m−2 s−1 estimated in Veg, PI, FL and H stages, respectively, with a seasonal average value of − 9.03 µmol m−2 s−1. During the DS it was observed that GPP values gradually increased as the cropping season progressed from Veg to H stage irrespective of any FPM. During the WS the average GPP values (modeled by RH method) were − 10.20, − 18.24, − 19.55 and − 18.41 µmol m−2 s−1 estimated in Veg, PI, FL and H stages, respectively, with a seasonal average value of − 17.30 µmol m−2 s−1, whereas the average GPP values in WS (modeled by Q10 method) were − 6.91, − 12.57, − 15.19 and − 10.08 µmol m−2 s−1 during Veg, PI, FL and H stages, respectively, with a seasonal average value of − 11.44 µmol m−2 s−1. The highest GPP was estimated during FL stage by both the FPM. It was observed that in both the seasons (DS and WS) the statistical mean difference between the GPP values estimated from RH and Q10 was significant although it was not found significant during Veg stages of both the cropping season in both the FPM. During the DF period the average GPP values, estimated from RH and Q10 method, were − 18.07 and − 12.36 µmol m−2 s−1, respectively, whereas during wet fallow (WF) the estimated average GPP values were − 16.58 and − 8.90 µmol m−2 s−1, respectively. The statistical mean difference between the GPP values estimated from both the FPM was also found to be significant in both the fallow period (Table 3). The seasonal and stage-wise variations of GPP in both the FPM are shown in Figs. 2 and 3, respectively.

Partitioning of net ecosystem exchange rectangular hyperbola into gross primary production and ecosystem respiration and diurnal variation (IST, Indian Standard Time) by method during the dry and wet seasons and fallow period of lowland paddy

Partitioning of net ecosystem exchange by rectangular hyperbola method into gross primary production and ecosystem respiration and diurnal variation (IST Indian Standard Time) in different crop growth stages of lowland paddy during dry and wet season
Daily, seasonal and yearlong variations in RE in two FPM
During the two cropping seasons (DS and WS) and fallow periods (DF and WF), the daily variation of average RE varied from 6.80 to 20.15 µmol m−2 s−1 and from 0.77 to 10.61 µmol m−2 s−1 obtained from RH and Q10 method, respectively. The annual average values of RE obtained from RH and Q10 method are 13.44 and 4.62 µmol m−2 s−1, respectively and the statistical mean difference between RE values estimated by the two FPM is significant (Table 4). During the DS the average RE values (modeled by RH method) were 7.78, 8.67, 12.65 and 15.17 µmol m−2 s−1 during Veg, PI, FL and H stages, respectively, with a seasonal average value of 11.24 µmol m−2 s−1. The average RE values (modeled by Q10 method) were 2.10, 2.63, 3.75 and 5.01 µmol m−2 s−1 during Veg, PI, FL and H stages, respectively, with a seasonal average value of 3.42 µmol m−2 s−1. During the DS it was observed that magnitude of RE gradually increased as the growing season progressed from Veg to H stage of paddy and estimated the highest at H stage. During the WS the average RE values (modeled by RH method) were 8.72, 15.39, 16.65 and 16.11 µmol m−2 s−1 during Veg, PI, FL and H stages, respectively, with a seasonal average value of 14.59 µmol m−2 s−1, whereas the average RE values (modeled by Q10 method) were 3.72, 4.53, 4.98 and 4.33 µmol m−2 s−1 during Veg, PI, FL and H stages of paddy, respectively, with a seasonal average value of 4.43 µmol m−2 s−1. The highest RE in the WS was estimated during FL stage in both the FPM (Table 4). It was observed that in both the season (DS and WS) the statistical mean difference between the RE values estimated from both the FPM was significant though it was not found significant during Veg stage of WS paddy (Table 3). During the DF period the average RE values, estimated from RH and Q10 method, were 15.85 and 6.55 µmol m−2 s−1, respectively, whereas during the WF the estimated average RE values were 12.67 and 5.70 µmol m−2 s−1, respectively. The statistical mean difference between the RE values estimated from both the FPM was also found to be significant in both the fallow period (Table 4). The seasonal and stage-wise variation of RE partitioned by both the FPM is shown in Figs. 4 and 5, respectively.

Partitioning of net ecosystem exchange by Q10 method into gross primary production and ecosystem respiration and diurnal variation (IST Indian Standard Time) in different crop growing seasons and fallow periods

Partitioning of net ecosystem exchange into gross primary production and ecosystem respiration by Q10 method in different crop growth stages and diurnal variation (IST Indian Standard Time) in lowland paddy during dry and wet season
Discussion
Variation of NEE in lowland paddy
Positive and negative sign of NEE denoted net CO2 emission into the atmosphere and net CO2 assimilation by lowland rice, respectively. The lowland paddy field acted as net CO2 sink throughout the entire growing season except for few days at the maturity and fallow period. The NEE in lowland paddy is mainly controlled by several factors and environmental variables such as latent heat, heat stress, vapor pressure deficit, canopy irradiance, stomatal response, circadian rhythm, growth stages of rice crop, leaf area index, biomass, high evaporative demand (Nair et al. 2011). Net ecosystem exchange also depends water management and the particular stages of the crop (Saito et al. 2005). It was observed that the amplitude of the daily variation in NEE increased with the progress of growing season and reached its maxima around the PI to FL stage and, then, decreased progressively till the maturity. It might be due to the reduction in leaf chlorophyll content or leaf senescence. These findings are in agreements with the findings of (Pakoktom et al. 2009) and (Bhattacharyya et al. 2013). Net ecosystem exchange becomes more negative during the day due to increasing CO2 assimilation by photosynthesis (increase in GPP) as air temperature and PAR increased (Figs. 6a, b, and 7). Similar results were reported by (Alberto et al. 2009) and (Nair et al. 2011). The presence of aquatic plants and algae in the floodwater may also affect the NEE (Miyata et al. 2000; Bhattacharyya et al. 2013).

a Mean daily air temperature (in Julian days). b Diurnal variation (IST Indian Standard Time) of average air temperature throughout the year

Variation of daily average photosynthetically active radiation in the year 2015
Variations in GPP in two FPM
It is evident from the results that the GPP estimated by the two different FPM (RH and Q10) significantly differed with each other and it is supported by their mean difference, RMSE and SD values (Tables 3 and 5). For DS, DF, WS and WF, the model RMSE values for RH method were 41.77, 21.40, 43.71 and 19.24% higher respectively, than Q10 method for the same following time frame, whereas the SD values were more in case of Q10 than RH method. Based on this observation it is obvious that RH method may be overestimating the GPP values as compared to the Q10 model in the lowland paddy field. The RH model used PAR data to partition NEE, and in wet season, PAR data (Fig. 7) are impacted due to cloud cover which may be attributed to error in the RH model while analyzing the data for WS period. The Q10 lacks this deficiency as it depends on temperature for partitioning NEE. While fitting the regression equation (for Q10) between nighttime air temperature and nighttime NEE we achieved a significant positive correlation (r2 ≈ 0.4–0.5) in both the season. Similar findings in this regard were reported by Stoy et al. (2006), Wohlfahrt and Galvagno (2017) and Oikawa et al. (2017).
Variations in RE in two FPM
The RE estimated by the two models significantly differed with each other which is supported by their mean difference, RMSE and SD values (Tables 4 and 5). For DS, DF, WS and WF, the model RMSE values for RH method were 86.81, 87.02, 84.22 and 77.30% higher, respectively, than Q10 method for the same time period. Similarly, the SD values were also more in case of RH as compared to Q10 method. From these observations it can be anticipated that the RH model was overestimating RE values much more than Q10 model; hence, Q10 was found best fit for this lowland paddy fields. However, Q10 approach may lead to systematic biases while partitioning as nighttime data were used to model daytime ecosystem respiration (RE), assuming that respiration processes did not cease in daytime and that the temperature responses of RE behaved similarly throughout the day and night. However, Reichstein et al. (2005) reported that these two assumptions are often violated. Nighttime plant respiration is often higher than daytime respiration which leads to an overestimation of daytime RE and GPP (Amthor 1995; Heskel et al. 2013; Kok 1949; Wehr et al. 2016; Wohlfahrt and Gu 2015). In the agricultural field, it is often difficult to predict daytime soil respiration from nighttime soil respiration rates due to the changing availability of substrate in soil (Oikawa et al. 2014; Tang et al. 2003; Vargas et al. 2011).
The model comparison and validation could have been more robust if we have had true ground measurements of the partitioning products (GPP and RE) in this region. Further research is needed for validating the FPM models more accurately and cohesively for a particular edaphoclimatic region. This study could be useful in data scarce region of the world.
Conclusions
In this study, the variation of the net ecosystem exchange of CO2 in lowland seasonal paddy cultivation is assessed and the best-suited model for partitioning of net ecosystem exchange of CO2 in lowland paddy is identified. The net ecosystem exchange of CO2 displayed a distinct daily and seasonal pattern during the paddy-growing season, and it was observed that lowland paddy has the capacity to sequester carbon from the atmosphere in the long run. It is evident from this study that the Q10 model unfailingly produced the most sensible C-flux estimates over rectangular hyperbolic (RH) method in lowland paddy field. The Q10 model that uses nighttime data to estimate RE, produced estimates which have lower RMSE values as compared to the other model. The RH model also requires many parameters, and accurate estimation of these parameters is important for better estimation of the partitioning products. The Q10 model predicted better estimates of RE and GPP values than the RH method that is seemingly realistic in nature, though further investigations are required to confirm these findings. The major drivers for these models are PAR and air temperature which varied temporally and controlled by many atmospheric and climatic factors. Again NEE is also controlled by many soil–plant-atmospheric factors. So understanding their interrelation is much needed. Partitioning of respiration from soil and biota is also not understood fully by scientific community. Considering these constraints, models are prone to bias and errors are normal. Long-term studies are needed to establish the real truth about efficiency and suitability of a model in a particular ecosystem. The present analysis might be useful for carbon sequestration and emission studies in wetland ecosystem and for partitioning of NEE in data scarce region where true estimates of GPP and RE are lacking
References
Alberto MC, Wassmann R, Hirano T, Miyata A, Kumar A, Padre A, Amante M (2009) CO2/heat fluxes in rice fields: comparative assessment of flooded and non-flooded fields in the Philippines. Agric For Meteorol 149:1737–1750
Amthor JS (1995) Higher plant respiration and its relationships to photosynthesis. In: Ecophysiology of photosynthesis. Springer, New York, pp 71–101
Baldocchi DD (2003) Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: past, present and future. Glob Change Biol 9:479–492. https://doi.org/10.1046/j.1365-2486.2003.00629.x
Baldocchi D (2008) TURNER REVIEW No: 15. ‘Breathing’ of the terrestrial biosphere: lessons learned from a global network of carbon dioxide flux measurement systems. Aust J Bot 56:1–26
Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Rödenbeck C, Arain MA, Baldocchi D, Bonan GB, Bondeau A (2010) Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science 834–838
Bhattacharyya P, Neogi S, Roy KS, Rao KS (2013) Gross primary production, ecosystem respiration and net ecosystem exchange in Asian rice paddy: an eddy covariance based approach. Curr Sci 104:67–75
Bhattacharyya P, Neogi S, Roy KS, Dash PK, Nayak AK, Mohapatra T (2014) Tropical low land rice ecosystem is a net carbon sink. Agric Ecosyst Environ 189:127–135
Chapin FS, Woodwell GM, Randerson JT, Rastetter EB, Lovett GM, Baldocchi DD, Clark DA, Harmon ME, Schimel DS, Valentini R, Wirth C (2006) Reconciling carbon-cycle concepts, terminology, and methods. Ecosyst 9(7):1041–1050
Chatterjee S (2014) Effects of irrigation, mulch and nitrogen on soil structure, carbon pools and input use efficiency in maize (Zea mays L.) (Doctoral dissertation, Division of Agricultural Physics, Indian Agricultural Research Institute, New Delhi). http://krishikosh.egranth.ac.in/handle/1/5810018929
Chatterjee S, Bandyopadhyay KK, Pradhan S, Singh R, Datta SP (2016) Influence of irrigation, crop residue mulch and nitrogen management practices on soil physical quality. J Indian Soc Soil Sci 64(4):351–367. https://doi.org/10.5958/0974-0228.2016.00048.7
Chatterjee S, Bandyopadhyay KK, Singh R, Pradhan S, Datta SP (2017) Yield and input use efficiency of maize (Zea mays L.) as influenced by crop residue mulch, irrigation and nitrogen management. J Indian Soc Soil Sci 65(2):199–209. https://doi.org/10.5958/0974-0228.2017.00023.8
Chatterjee S, Bandyopadhyay KK, Pradhan S, Singh R, Datta SP (2018) Effects of irrigation, crop residue mulch and nitrogen management in maize (Zea mays L.) on soil carbon pools in a sandy loam soil of Indo-gangetic plain region. CATENA 165:207–216. https://doi.org/10.1016/j.catena.2018.02.005
Chatterjee D, Tripathi R, Chatterjee S, Debnath M, Shahi M, Bhattacharyya P, Swain CK, Bhattacharya BK, Nayak AK (2019a) Characterization of land surface energy fluxes in a tropical lowland rice paddy. Theoret Appl Climatol 136(1–2):157–168. https://doi.org/10.1007/s00704-018-2472-y
Chatterjee D, Nayak AK, Vijayakumar S, Debnath M, Chatterjee S, Swain CK, Bihari P, Mohanty S, Tripathi R, Shahid M, Kumar A (2019b) Water vapor flux in tropical lowland rice. Environ Monit Assess 191(9):550. https://doi.org/10.1007/s10661-019-7709-4
Desai AR, Richardson AD, Moffat AM, Kattge J, Hollinger DY, Barr A, Falge E, Noormets A, Papale D, Reichstein M, Stauch VJ (2008) Cross-site evaluation of eddy covariance GPP and RE decomposition techniques. Agric For Meteorol 148:821–838
Falge E, Tenhunen J, Baldocchi D, Aubinet M, Bakwin P, Berbigier P, Bernhofer C, Bonnefond JM, Burba G, Clement R, Davis KJ (2002) Gap filling strategies for defensible annual sums of net ecosystem exchange. Agric For Meteorol 107:43–69
Government of India (GOI) (2014) All India report on number and area of operational holdings. Agriculture Census Division, Department of Agriculture &Co-Operation & Farmers Welfare. Ministry of Agriculture & Farmers Welfare, New Delhi
Gu L, Falge EM, Boden T, Baldocchi DD, Black TA, Saleska SR, Suni T, Verma SB, Vesala T, Wofsy SC, Xu L (2005) Objective threshold determination for night time eddy flux filtering. Agric For Meteorol 128:179–197
Hagen SC, Braswell BH, Linder E, Frolking S, Richardson AD, Hollinger DY (2006) Statistical uncertainty of eddy flux based estimates of gross ecosystem carbon exchange at Howland Forest. Maine J Geophys Res Atmos 111:D08S03. https://doi.org/10.1029/2005jd006154
Heskel MA, Atkin OK, Turnbull MH, Griffin KL (2013) Bringing the Kok effect to light: a review on the integration of daytime respiration and net ecosystem exchange. Ecosphere 4:1–14
Jung M, Reichstein M, Margolis HA, Cescatti A, Richardson AD, Arain MA, Arneth A, Bernhofer C, Bonal D, Chen J, Gianelle D (2011) Global patterns of land-atmosphere fluxes of carbon dioxide, latent heat, and sensible heat derived from eddy covariance, satellite, and meteorological observations. J Geophys Res 116
Kaimal JC, Finnigan JJ (1994) Atmospheric boundary layer flows. Oxford University Press, Oxford. https://doi.org/10.1016/0012-8252(94)90026-4
Kok B (1949) On the interrelation of respiration and photosynthesis in green plants. Biochim Biophys Acta 3:625–631
Lasslop G, Reichstein M, Papale D, Richardson AD, Arneth A, Barr A, Stoy P, Wohlfahrt G (2010) Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: critical issues and global evaluation. Glob Change Biol 16(1):187–208
Lee X, Fuentes JD, Staebler RM, Neumann HH (1999) Long term observation of the atmospheric exchange of CO2 with a temperate deciduous forest in southern Ontario, Canada. J Geophys Res Atmos 104:15975–15984
Liu H, Peters G, Foken T (2001) New equations for sonic temperature variance and buoyancy heat flux with an omnidirectional sonic anemometer (J). Bound-Layer Meteorol 100:459–468
Massman WJ, Lee X (2002) Eddy covariance flux corrections and uncertainties in long-term studies of carbon and energy exchanges. Agric For Meteorol 113(1–4):121–144
Mauder M, Foken T (2011) Eddy-covariance software TK3 [Data set], Documentation and instruction manual of the eddy-covariance software package TK3 (update), University of Bayreuth, Bayreuth, Germany, https://doi.org/10.5281/zenodo.20349, 2015.
Mauder M, Liebethal C, Göckede M, Leps JP, Beyrich F, Foken T (2006) Processing and quality control of flux data during LITFASS-2003. Boundary-Layer Meteoro 121:67–88. https://doi.org/10.1007/s10546-006-9094-0
Miyata A, Leuning R, Denmead OT, Kim J, Harazono Y (2000) Carbon dioxide and methane fluxes from an intermittently flooded paddy field. Agric For Meteorol 102:287–303. https://doi.org/10.1016/S0168-1923(00)00092-7
Nair R, Juwarkar AA, Wanjari T, Singh SK, Chakrabarti T (2011) Study of terrestrial carbon flux by eddy covariance method in revegetated manganese mine spoil dump at Gurgaon, India. Clim. Change 106:609–619. https://doi.org/10.1007/s10584-010-9953-z
Oikawa PY, Grantz DA, Chatterjee A, Eberwein JE, Allsman LA, Jenerette GD (2014) Unifying soil respiration pulses, inhibition, and temperature hysteresis through dynamics of labile soil carbon and O2. J Geophys Res. https://doi.org/10.1002/2013JG002434
Oikawa PY, Sturtevant C, Knox SH, Verfaillie J, Huang YW, Baldocchi DD (2017) Revisiting the partitioning of net ecosystem exchange of CO2 into photosynthesis and respiration with simultaneous flux measurements of 13CO2and CO2, soil respiration and a biophysical model, CANVEGP. Agric For Meteorol 234–235:149–163
Pakoktom T, Aoki M, Kasemsap P, Boonyawat S, Attarod P (2009) CO2 and H2O fluxes ratio in paddy fields of Thailand and Japan. Hydrol Res Lett 3:10–13
Papale D, Reichstein M, Aubinet M, Canfora E, Bernhofer C, Kutsch W, Longdoz B, Rambal S, Valentini R, Vesala T, Yakir D (2006) Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: algorithms and uncertainty estimation. Biogeosciences 3:571–583
Pathak H, Nayak AK, Jena M, Singh ON, Samal P, Sharma SG (eds) (2018) Rice research for enhancing productivity, profitability and climate resilience. ICAR-National Rice Research Institute, Cuttack, p 527
Reichstein M, Falge E, Baldocchi D, Papale D, Aubinet M, Berbigier P, Bernhofer C, Buchmann N, Gilmanov T, Granier A, Grünwald T (2005) On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Glob Change Biol 11:1424–1439. https://doi.org/10.1111/j.1365-2486.2005.001002.x
Richardson AD, Hollinger DY, Burba GG, Davis KJ, Flanagan LB, Katul GG, Munger JW, Ricciuto DM, Stoy PC, Suyker AE, Verma SB (2006) A multi-site analysis of uncertainty in tower-based measurements of carbon and energy fluxes. Agric For Meteorol 136:1–18
Ruimy A, Jarvis PG, Baldocchi DD, Saugier B (1995) CO2 fluxes over plant canopies and solar radiation: a review. Adv Ecol Res 26:1–68. https://doi.org/10.1016/S0065-2504(08)60063-X
Saito M, Miyata A, Nagai H, Yamada T (2005) Seasonal variation of carbon dioxide exchange in rice paddy field in Japan. Agric For Meteorol 135(1):93–109. https://doi.org/10.1016/j.agrformet.2005.10.007
Sarmiento JL, Wofsy SC (1999) A US carbon cycle science plan: Report of the Carbon and Climate Working Group. US Global Change Res. Program, Washington, DC
Stoy PC, Katul GG, Siqueira MB, Juang JY, McCarthy HR, Kim HS, Oishi AC, Oren R (2005) Variability in net ecosystem exchange from hourly to inter annual time scales at adjacent pine and hardwood forests: a wavelet analysis. Tree Physiol 25:887–902
Stoy PC, Katul GG, Siqueira MB, Juang JY, Novick KA, Uebelherr JM, Oren R (2006) An evaluation of models for partitioning eddy covariance-measured net ecosystem exchange into photosynthesis and respiration. Agric For Meteorol 141(1):2–18
Swain CK, Bhattacharyya P, Singh NR, Neogi S, Sahoo RK, Nayak AK, Zhang G, Leclerc MY (2016) Net ecosystem methane and carbon dioxide exchange in relation to heat and carbon balance in lowland tropical rice. Ecol Eng 95:364–374
Swain CK, Nayak AK, Bhattacharyya P, Chatterjee D, Chatterjee S, Tripathi R, Singh NR, Dhal B (2018a) Greenhouse gas emissions and energy exchange in wet and dry season rice: eddy covariance-based approach. Environ Monit Assess 190:423. https://doi.org/10.1007/s10661-018-6805-1
Swain CK, Bhattacharyya P, Nayak AK, Singh NR, Neogi S, Chatterjee D, Pathak H (2018b) Dynamics of net ecosystem methane exchanges on temporal scale in tropical lowland rice. Atmos Environ 191:291–301
Tang J, Baldocchi DD, Qi Y, Xu L (2003) Assessing soil CO2 efflux using continuous measurements of CO2 profiles in soils with small solid-state sensors. Agric For Meteorol 118:207–220
Tanner CB, Thurtell GW (1969) Anemoclinometer measurements of reynolds stress and heat transport in the atmospheric surface layer. The University of Wisconsin Tech, pp 82, Rep ECOM-66-G22-F. http://dx.doi.org/AD0689487
Vargas R, Baldocchi DD, Bahn M, Hanson PJ, Hosman KP, Kulmala L, Pumpanen J, Yang B (2011) On the multi-temporal correlation between photosynthesis and soil CO2 efflux: reconciling lags and observations. New Phytol 191:1006–1017
Vickers D, Mahrt L (1997) Quality control and flux sampling problems for tower and aircraft data. J Atmos Ocean Technol 14(3):512–526
Webb EK, Pearman GI, Leuning R (1980) Correction of flux measurements for density effects due to heat and water vapor transfer. Q J Roy Meteor Soc 106:85–100. https://doi.org/10.1002/qj.49710644707
Wehr R, Munger JW, McManus JB, Nelson DD, Zahniser MS, Davidson EA, Wofsy SC, Saleska SR (2016) Seasonality of temperate forest photosynthesis and daytime respiration. Nature 534:680–683
Wilczak JM, Oncley SP, Stage SA (2001) Sonic anemometer tilt correction algorithms. Boundary Layer Meteorol 99(1):127–150. https://doi.org/10.1023/A:1018966204465
Wohlfahrt G, Galvagno M (2017) Revisiting the choice of the driving temperature for eddy covariance CO2 flux partitioning. Agric For Meteorol 237–238:135–142
Wohlfahrt G, Gu L (2015) The many meanings of gross photosynthesis and their implication for photosynthesis research from leaf to globe. Plant, cell & environ 38(12):2500-2507
Wohlfahrt G, Bahn M, Haslwanter A, Newesely C, Cernusca A (2005) Estimation of daytime ecosystem respiration to determine gross primary production of a mountain meadow. Agric For Meteorol 130:13–25
Acknowledgements
The authors acknowledge the colleagues and research scholars at ICAR-National Rice Research Institute, Cuttack, India, who have assisted in this study. The first author sincerely acknowledges Dr. Jingyi Huang of University of Wisconsin-Madison, USA, for his suggestions to improve the manuscript. The first author also acknowledges the Indian Council of Agricultural Research for granting study leave and providing financial support.
Funding
This study has been supported by the Grant of National Innovations on Climate Resilient Agriculture, Indian Council of Agricultural Research, New Delhi, India.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors assert that there is no conflict of interest.
Rights and permissions
About this article

Cite this article
Chatterjee, S., Swain, C.K., Nayak, A.K. et al. Partitioning of eddy covariance-measured net ecosystem exchange of CO2 in tropical lowland paddy. Paddy Water Environ 18, 623–636 (2020). https://doi.org/10.1007/s10333-020-00806-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10333-020-00806-7