
Abstract
The supply of phosphorus, the essential element for plant growth and development, is often limited in natural environments. Plants employ multiple physiological strategies to minimize the impact of phosphate deficiency. In deciduous trees, phosphorus is remobilized from senescing leaves in autumn and stored in other tissues for reuse in the following spring. We previously monitored the annual changes in leaf phosphate content of white poplar (Populus alba) growing under natural conditions and found that about 75 % of inorganic and 60 % of organic leaf phosphates observed in May were remobilized by November. In order to analyze this process (such annual events), we have established a model system, in which an annual cycle of phosphate re-translocation in trees can be simulated under laboratory conditions by controlling temperature and photoperiod (=‘shortened annual cycle’). This system evidently allowed us to monitor the annual changes in leaf color, phosphate remobilization from senescent leaves, and bud break in the next spring within five months. This will greatly facilitate the analysis of cellular and molecular mechanisms of annual phosphate re-translocation in deciduous trees.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In woody plants, seasonal remobilization of nutrients has been studied for a long time since this process is important for plants to survive in nutrient-limited environments (Chapin and Kedrowsky 1983; Cherbuy et al. 2001; Keskitalo et al. 2005). Both deciduous (birch, alder, larch, poplar) and evergreen (spruce, oak) trees are known to remobilize various nutrients such as C, N, P, and S from senescing leaves before leaf fall. Remobilization of nutrients from senescing leaves enables plants to reuse them immediately. Residual nutrients, which are not remobilized, can be recycled through litterfall. However, decomposition and re-mineralization of the litter take several years before these nutrients become available to plants (Aerts 1996). It is an advantage for perennial plants to retain nutrients in core tissues that are least affected by alterations in the external environments.
Phosphorus is one of the precious nutrients, whose supply is often limited in natural and agricultural environments. Remobilization of phosphorus is therefore quite important for perennials such as deciduous woody plants. In Populus tremura, phosphate content in leaves decrease by about 80 % during autumn (Keskitalo et al. 2005). In deciduous taiga trees, it was revealed that P was stored in buds and stems during winter primarily as phospholipids, non-hydrolyzable esters (it was defined as the ester which was not hydrolyzed for 10 min in 1 M HCl at 95 °C, and phosphoryl choline is presumed as one of candidates), and nucleic acids, and they were converted to inorganic phosphpates (Pi) in spring (Chapin and Kedrowsky 1983). However, the molecular mechanisms that underlie the remobilization of phosphate in autumn are still unclear.
Recently, the annotated whole genome sequence of Populus trichocarpa Torr. & Groy, a model perennial plant, was released (Tuskan et al. 2006), and methods for genetic analysis in Populus have been established (Busov et al. 2010). Sequence and expression profiles of the Pht1 phosphate transporter gene family in Populus trichocarpa have been revealed. Some of these genes are thought to play roles in Pi remobilization from source to sink tissues (Loth-Pereda et al. 2011).
In herbaceous plants, it is known that Pi is re-translocated from mature lower leaves to young upper leaves, and that re-translocation is accelerated by phosphate starvation. In Arabidopsis thaliana, molecular mechanisms of phosphate allocation in plants have been well studied (Chiou et al. 2006; Fujii et al. 2005; Lin et al. 2009).
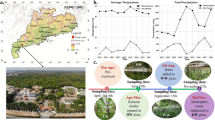
Phosphate remobilization has been often reported using field trees (Chapin and Kedrowsky 1983; Keskitalo et al. 2005). In the field condition, however, seasonal remobilization may be strongly affected by many factors such as the climate changes and nutritional conditions in soil. To simplify the experimental system, in the present study, we have established a model system for studying seasonal phosphate re-translocation in deciduous trees under laboratory conditions (=‘shortened annual cycle’) by controlling temperature and day length. This system eliminates environmental disturbances and allows us to analyze seasonal re-translocation of phosphates in a shorter period (about 4–5 months). Leaf-age dependent phosphate allocation and re-translocation from mature lower leaves to young upper leaves within a tree is investigated in this system.
Materials and methods
Plant materials
For the field analysis, leaves of a white poplar (Populus alba) were collected every month between 13:30 to 14:30 PM from April 2010 to November 2011, from a field at Uji campus of Kyoto University, Japan (34°91′N, 135°80′E, elevation 24 m). The tree was 6-year-old in 2010, and stood solitary. Leaves were randomly collected between 1 and 2.5 m from the ground.
For the laboratory analysis, leaves were collected from potted Populus alba clones on the last day of each culture stage (see culture conditions). Leaves were numbered from apex to the base. Younger leaves smaller than 3 cm in length were not counted.
In both analyses, the fresh weight of each sample comprising one leaf was measured, and then samples were frozen in liquid nitrogen and stored at −20 °C until extraction of phosphates.
Culture conditions
Poplar shoots with five leaves (about 10 cm) were cut from potted cuttings, which were maintained at stage 1 (see Table 1) in laboratory. After rooting with 1/5 MS medium containing 0.1 mg l−1 indole-3-butyric acid (IBA), cuttings were transferred to pots (diameter 10 cm; height 8 cm) containing vermiculite, which had been washed by tap water ten times and by distilled water five times. Potted cuttings were placed in a culture chamber: stage 1, LP-1, 5P (NK system Osaka, Japan); stage 2, LH-350S (NK system, Osaka, Japan); and stage 3, LH-220S (NK system, Osaka, Japan) under the conditions listed in Table 1. Light was supplied by fluorescence tubes: stage 1, FPR96EX-N/A (Panasonic, Osaka, Japan); stage 2, FL20SS ENW/18 (Panasonic, Osaka, Japan) and FL4SS EX-N/37-H (TOSHIBA, Tokyo, Japan); and stage 3, FL20SEX-N-HG, FL40SEX-N-HG, and FL15EX-N-X (NEC, Tokyo, Japan).
Plants were fertilized with 1/5 MS medium (4.12 mM NH4NO3, 3.76 mM KNO3, 0.02 mM H3BO3, 0.02 mM MnSO4, 7.4 μM ZnSO4, 0.2 μM KI, 0.2 μM Na2MoO4, 0.02 μM CuSO4, 0.02 μM CoCl2, 0.6 mM CaCl2, 0.25 mM KH2PO4, 0.3 mM MgSO4, and 0.02 mM FeNaEDTA) (in total 1 l for six potted cuttings) once a week at stage 1 and thereafter they were irrigated with distilled water only.
Extraction of Pi and organic phosphates (Po)
Frozen leaves were ground with a mortar and pestle in liquid nitrogen. A whole leaf including petiole was measured as one sample. Based on the fresh weight of each leaf, proper amount of water was added and the samples were homogenized. The homogenate was centrifuged at 20,400 g for 10 min at 4 °C (MX-301, TOMY, Tokyo, Japan). The supernatant was boiled for 7 min at 98 °C (DRY THERMO UNIT TAH-1G, TAITEC, Saitama, Japan), centrifuged again at 20,400g for 10 min at 4 °C. The supernatant was filtered through 0.5 μm Ultrafree centrifuge filter (Millipore, Massachusetts, USA) at 5,000g for 3 min at 4 °C. The extract was stored at −20 °C until measurement.
Pi and Po measurements
Pi was measured by a modified molybdenum blue method (Murphy and Riley 1962). Total phosphorus was measured by molybdenum blue method after degradation of Po by potassium persulfate oxidation. Po was then calculated by subtraction of the Pi value from the total phosphorus value.
Potassium persulfate oxidation
Samples were diluted as necessary so as to fit within the concentration range of the molybdenum blue assay (absorbance 0.05 ~ 0.2). To these diluted solutions, potassium persulfate solution (50 g l−1) was added in a volume ratio of 1:4, and heated in an autoclave at 120 °C for 90 min. The resulting solutions were diluted further twofold with H2O and total phosphate was measured by the molybdenum blue method.
The molybdenum blue assay method
For assay of the various phosphate fractions, the following stock solutions were prepared: ammonium molybdate (30 g l−1), 5 N sulfuric acid (245 g l−1), ascorbic acid (54 g l−1, prepared just before use), potassium antimonyl tartrate (1.36 g l−1). Just before the measurement, stock reagents of ammonium molybdate, sulfuric acid, ascorbic acid and potassium antimonyl tartrate, were mixed in a volume ratio of 2:5:2:1. The mixed reagent was added to inorganic or total phosphate solution in a volume ratio of 1:10. After 60 min, the absorbance was read on a spectrophotometer (DU 730, BECKMAN COULTER, Brea, CA, USA) at 855 nm. Potassium dihydrogen phosphate was used as a phosphate standard.
Results
Annual changes in leaf phosphate contents in the field
To confirm remobilization of phosphates from the autumn leaves, contents of Pi and Po in field poplar leaves were measured from April 2010 to November 2011 (Fig. 1a). In the poplar tree, bud break occurred in April followed by leaf expansion. From October to November, leaves turned yellow, and fell until December. In 2010, Pi contents in leaves were nearly constant from May to August (about 3 ~ 3.5 μmol leaf−1), whereas in 2011 they generally increased from April to August. Although the monthly Pi contents in 2010 and 2011 were different, the maximum Pi content in August 2011 was nearly equal to that in August 2010. The decrease in leaf Pi contents commenced in September, and from August to November, about 75 % of Pi in leaves was remobilized (71 % in 2010, 78 % in 2011). In 2010, leaf Po content decreased continuously from May to November, and about 57 % of Po content of that in May was remobilized. In 2011, Po contents fluctuated throughout the year. From August to November, about 57 % of Po content of that in August was remobilized. Po content in October 2011 was high, which have been due to an increase in the fresh weight of sample leaves in 2011 (Fig. 1d). Po concentrations based on fresh weight (mmol kg fresh weight (FW)−1) showed the similar tendency, and did not show such an increase in October 2011 (Fig. 1c). About 66 and 73 % of total phosphate in leaves were remobilized in 2010 and 2011, respectively (Fig. 1b).

Seasonal changes in leaf phosphates and leaf fresh weight in a field poplar. a Seasonal changes in inorganic and organic phosphate contents per leaf (Pi, Po). b Seasonal changes in total phosphate contents per leaf (Pt). c Seasonal changes in inorganic and organic phosphate levels expressed as per leaf fresh weight. d Seasonal changes in leaf fresh weight. Black squares and white squares indicate measured values in 2010 and 2011, respectively. Inorganic and organic phosphates are indicated by solid and broken lines, respectively (mean ± SE, n = 5 in 2010, n = 3 in 2011)
Stage-dependent changes of leaf phosphate contents in the shortened annual cycle
Popular trees were planted in pots and grown at 25 °C with 14 h light (c.a. 200 µmol m−2 s−1) for 1 month to represent spring and summer conditions (stage 1). In this system, phosphate contents in leaves depend on the amount of fertilized P. To approximate phosphate content in the natural leaves during growing season (3–4 µmol leaf−1), plants were fertilized with 1/5 MS medium once a week at only this stage (1 l per 6 potted cuttings). Then, to acclimatize to low temperature, trees were transferred to 15 °C with 8 h light (c.a. 30–40 µmol m−2 s−1) and cultured for 1 month to represent autumn conditions (stage 2). Finally, trees were cultured at 5 °C with 8 h light (c.a. 30–40 µmol m−2 s−1) for 2–3 months to represent winter conditions (stage 3). Table 1 summarizes these conditions.
During stages 1 and 2, trees were growing, and increased the number of countable leaves. Approximately 12 leaves developed during stage 1 and around four leaves during stage 2. The first leaf at the end of stage 1 corresponds to the third to fifth leaf at the end of stage 2. During stage 3, the number of leaves increased by no more than two. At that time, although the trees seemed to stop shoot apical growth, the leaf number increased only by expansion of uncounted small leaves in the preceding stage. Thus, the total number of leaves in the tree sapling before leaf fall became about 23, compared to around five at the beginning of culture. In stage 3, leaf coloration (beautiful yellow, see Fig. 2) and leaf fall occurred in the upper leaves, while the lower leaves withered and fell, but did not show the beautiful yellow color (Fig. 2b). When defoliated trees were again moved to stage 1 conditions, new leaves emerged within one or two weeks.

States of poplar trees in the shortened annual cycle system. a Outline of the shortened annual cycle system. After 2 months in stage 3, leaf coloration occurred in upper leaves. b Fourth leaf (left) and tenth leaf (right) in the same tree at the end of stage 3. Leaves were numbered from apical to basal. Immature leaves smaller than 3 cm were not counted
To measure phosphate distribution in poplar trees in this system, Pi and Po contents in leaves were measured along the shoot axis (Fig. 3a). In stage 1, except for the first leaf, Pi and Po contents per leaf were higher in young upper leaves than in lower leaves. The first leaf was not mature, and its fresh weight (approximately 137 mg) was smaller than lower leaves (the fifth leaf, 527 mg). However, phosphate levels of the first leaf (Pi 8.3 mmol kg FW−1, Po 7.4 mmol kg FW−1) was roughly the same value as the third leaf (Pi 8.9 mmol kg FW−1, Po 5.7 mmol kg FW−1), and higher than that of the fifth leaves (Pi 6.0 mmol kg FW−1, Po 2.6 mmol kg FW−1) and downward ones. In stage 2, Pi and Po contents were also higher in young upper leaves than in lower leaves. However, Pi contents in leaves were lower than in stage 1. In stage 3, Pi and Po contents decreased greatly in the fifth to the seventh leaves. The rate of phosphate remobilization during stages 1–3 was high in the fifth and the sixth leaves, and was low in the first, the third and lower than the sixth leaves (Fig. 3b). This shows that the efficiency of remobilization of phosphate varies according to the leaf position from apex.

Phosphate contents in poplar leaves in the shortened annual cycle system. a Leaf phosphate contents along shoot axis. Leaves were numbered from apical to basal. Immature leaves smaller than 3 cm were not counted. b The rate of phosphate remobilization along shoot axis. The rate was calculated from values at stage 1 and stage 3. c Changes in phosphate contents in sixth leaves during stage 1 to 3. Inorganic phosphate contents are indicated by white bars or solid lines. Organic phosphate contents are indicated by black bars or broken lines (mean ± SE, n = 3). At stage 1 of the second cycle, leaves were collected randomly from trees, because it was difficult to determine the leaf number
The first leaf at the end of stage 1 became the third to fifth leaf at the end of stage 2. Phosphate contents in this leaf increased during stage 2. Phosphate contents in lower leaves (sixth and seventh at stage 2 that correspond to about third leaf at stage 1) decreased during stage 2 (Fig. 3a). It was presumed that changes in phosphate contents in leaves were due to re-translocation of Pi and Po from lower leaves to younger upper leaves.
In a leaf, source and sink relationships change depending on the stage of development. When leaves are young, they act as sinks and are supplied with P from older tissues. As they become mature, they progressively change from sink to source, eventually exporting P to younger leaves. This decrease of leaf phosphate contents due to plant growth-dependent re-translocation needs to be distinguished from the remobilization of leaf phosphate, which occurs during autumn senescence. P contents of the sixth leaves were measured in each stage under the assumption the same position of the leaf has the same capacity as the source tissue for the sink tissues (Fig. 3c). It was considered that the influence of phosphate re-translocation would be suppressed by using the same leaf position. When compared to the autumn remobilization in the field (Fig. 1a), phosphate contents in the sixth leaves of the laboratory grown plants showed a similar decrease from stages 1–3 (Pi 88 %, Po 58 %). At stage 1 of the second cycle, Pi and Po contents in leaves increased again (Fig. 3c). Bud break occurred at the terminal bud and most axillary buds at the same time. Since it was difficult to determine the growth-dependent leaf number, leaves were collected randomly from trees at the second stage 1.
Discussion
In field measurements, remobilization of Pi and Po from senescing leaves occurred in late summer and autumn (Fig. 1). Phosphate remobilization from senescent leaves is an important strategy for perennials to conserve phosphorus. In nature, seasonal events like leaf fall and bud break occur only once a year and these events are strongly influenced by many factors such as temperature, rainfall, and length of sunshine. This complicates analysis by molecular means of the processes that contribute to re-translocation within the plant and remobilization during senescence. In an attempt to control the phenology, we have established and tested a controlled environmental system that mimics the developmental events that occur under field conditions. The main differences with this new system are that the phenology is more predictable, and the annual cycle can be achieved within approximately five months. This system enabled us to analyze phosphate re-translocation in a tree in a shorter period without disturbance from variable climatic influences.
In this shortened annual cycle system, by controlling temperature and day length, we succeeded in reproducing leaf coloration and phosphate remobilization from leaves similar to that which occurs under field conditions (Figs. 2, 3). In the field, phosphate remobilization started in September before leaf coloration. The precise trigger for this phosphate remobilization from senescent leaves is still unclear. In previous studies, it was revealed that Populus responded to a critically shortened day length with the cessation of elongation growth and the formation of terminal buds. These events are reported to be controlled by the CONSTANS(CO)/FLOWERING LOCUS T(FT) regulatory module in Populus (Böhlenius et al. 2006; van der Schoot and Rinne 2011). In the European aspen (Populus tremula), autumn senescence starts every year at around the same date, suggesting that the main trigger is the shortening of day length in the photoperiod (Keskitalo et al. 2005). In nitrogen recycling, the expression of bark storage protein genes during autumn is also regulated by the photoperiod, through the action of phytochrome (Zhu and Coleman 2001). It is seems likely that phosphate remobilization from senescing leaves and subsequent winter storage of phosphate are also regulated by the photoperiod. Further studies will be required to ensure whether the shortened senescence period and artificial climate control in the present study affects the in situ mechanisms of phosphate re-translocation.
In our shortened annual cycle system, trees that underwent the three developmental stages reverted back to stage 1 when the appropriate conditions were provided, and within two weeks, bud break occurred and phosphate contents in new leaves were much higher than in leaves at stage 3. It was considered that phosphate contents in these new leaves were mainly supplied from stem and/or buds because those trees were fertilized only at the previous stage 1. However, it is also possible that some residual fertilizer remained in the soil. From the results obtained, it was difficult to distinguish between phosphate remobilization from senescing leaves in autumn and phosphate re-translocation from old mature leaves to young upper leaves as the tree developed. Also, phosphate remobilization from leaf may be influenced by changes in physiological conditions of other sink tissues (shoot apex, root, bud, or stem). We now have a suitable system for investigation of a range of nutrient-related questions including phosphate fluxes between different tissues, and the molecular processes that underlie changes in phosphate movements within the plant during its annual cycle. This system will also be able to be applied to other seasonal events in deciduous trees, such as re-translocation of other nutrients, bud break, cambium reactivation, and abscission layer formation. The present system will be a valuable tool for analyses of interactive approaches using both field and laboratory analyses.
References
Aerts R (1996) Nutrient resorption from senescing leaves of perennials: are there general patterns? J Ecol 84:597–608
Böhlenius H, Huang T, Charbonnel-Cambaa L, Brunner A, Jansson S, Strauss S, Nilsson O (2006) CO/FT regulatory module controls timing of flowering and seasonal growth cessation in trees. Science 312:1040–1043
Busov VB, Strauss SH, Pilate G (2010) Transformation as a tool for Genetic analysis in Populus. In: Jansson S, Bhalerao RP, Groover AT (eds) Genetics and genomics of Populus, 1st edn. Springer, New York, pp 113–133
Chapin FSI, Kedrowsky RA (1983) Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocations in evergreen and deciduous Taiga trees. Ecology 64:376–391
Cherbuy B, Joffre R, Gillon D, Rambal S (2001) Internal remobilization of carbohydrates, lipids, nitrogen and phosphorus in the Mediterranean evergreen oak Quercus ilex. Tree Physiol 21:9–17
Chiou TJ, Aung K, Lin SI, Wu CC, Chiang SF, Su CL (2006) Regulation of phosphate homeostasis by microRNA in Arabidopsis. Plant Cell 18:412–421
Fujii H, Chiou TJ, Lin SI, Aung K, Zhu JK (2005) A miRNA involved in phosphate-starvation response in Arabidopsis. Curr Biol 15:2038–2043
Keskitalo J, Bergquist G, Gardeström P, Jansson S (2005) A cellular timetable of autumn senescence. Plant Physiol 139:1635–1648
Lin WY, Lin SI, Chiou TJ (2009) Molecular regulators of phosphate homeostasis in plants. J Exp Bot 60:1427–1438
Loth-Pereda V, Orsini E, Courty PE, Lota F, Kohler A, Diss L, Blaudez D, Chalot M, Nehls U, Bucher M, Martin F (2011) Structure and expression profile of the phosphate Pht1 transporter gene family in mycorrhizal Populus trichocarpa. Plant Physiol 156:2141–2154
Murphy J, Riley JP (1962) A modified single solution method for the determination of phosphate in natural waters. Anal Chim Acta 27:31–36
Tuskan GA, Difazio S, Jansson S, Bohlmann J, Grigoriev I, Hellsten U, Putnam N, Ralph S, Rombauts S, Salamov A, Schein J, Sterck L, Aerts A, Bhalerao RR, Bhalerao RP, Blaudez D, Boerjan W, Brun A, Brunner A, Busov V, Campbell M, Carlson J, Chalot M, Chapman J, Chen GL, Cooper D, Coutinho PM, Couturier J, Covert S, Cronk Q, Cunningham R, Davis J, Degroeve S, Déjardin A, dePpamphilis C, Detter J, Dirks B, Dubchak I, Duplessis S, Ehlting J, Ellis B, Gendler K, Goodstein D, Gribskov M, Grimwood J, Groover A, Gunter L, Hamberger B, Heinze B, Helariutta Y, Henrissat B, Holligan D, Holt R, Huang W, Islam-Faridi N, Jones S, Jones-Rhoades M, Jorgensen R, Joshi C, Kangasjärvi J, Karlsson J, Kelleher C, Kirkpatrick R, Kirst M, Kohler A, Kalluri U, Larimer F, Leebens-Mack J, Leplé JC, Locascio P, Lou Y, Lucas S, Martin F, Montanini B, Napoli C, Nelson DR, Nelson C, Nieminen K, Nilsson O, Pereda V, Peter G, Philippe R, Pilate G, Poliakov A, Razumovskaya J, Richardson P, Rinaldi C, Ritland K, Rouzé P, Ryaboy D, Schmutz J, Schrader J, Segerman B, Shin H, Siddiqui A, Sterky F, Terry A, Tsai CJ, Uberbacher E, Unneberg P, Vahala J, Wall K, Wessler S, Yang G, Yin T, Douglas C, Marra M, Sandberg G, Van de Peer Y, Rokhsar D (2006) The genome of black cottonwood, Populus trichocarpa (Torr & Gray). Science 313:1596–1604
van der Schoot C, Rinne PLH (2011) Dormancy cycling at the shoot apical meristem: transitioning between self-organization and self-arrest. Plant Sci 180:120–131
Zhu B, Coleman GD (2001) Phytochrome-mediated photoperiod perception, shoot growth, glutamine, calcium, and protein phosphorylation influence the activity of the poplar bark storage protein gene promoter (bspA). Plant Physiol 126:342–351
Acknowledgments
We would like to offer our heartfelt thanks to Prof. Shinobu Satoh for his advice about culture conditions for the establishment of a shortened annual cycle. We also greatly appreciate Dr. Rob Reid (University of Adelaide, Adelaide, Australia) and Dr. Shanti S. Sharma (Himachal Pradesh University, Shimla, India) for their kind discussion and correction of this manuscript. This work was supported by a Grant-in-Aid for Scientific Research of Innovative Areas from the Japanese Ministry of Education, Sports, Culture, Science, and Technology on “Perceptive plants (22120006)”. YK is grateful for a SUNBOR Scholarship.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Kurita, Y., Baba, K., Ohnishi, M. et al. Establishment of a shortened annual cycle system; a tool for the analysis of annual re-translocation of phosphorus in the deciduous woody plant (Populus alba L.). J Plant Res 127, 545–551 (2014). https://doi.org/10.1007/s10265-014-0634-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10265-014-0634-2