
Abstract
To improve our understanding of the complex genetic and ecological structure of bottlenose dolphin (Tursiops truncatus) populations, we examined the acoustic features of communication signals from two geographically contiguous areas: the Central–Eastern North Atlantic and the Mediterranean Sea. Variations in the whistles were evaluated for four locations. Ten signal parameters were measured and used to statistically differentiate between the areas. Over 79 % of sightings were correctly classified by discriminant function analysis, confirming an acoustic differentiation between the two basins. The results of cluster analysis using the mean values of the parameters for each sighting showed that the three easternmost sightings from the Mediterranean and one sighting from the Canary archipelago formed a separate cluster from the rest of the Atlantic. The two sightings from the Alboran Sea in the west Mediterranean were grouped with the Atlantic recordings. There was more variability in whistles from the Atlantic Ocean consistent with data from genetic and photo-identification studies that document resident and non-resident animals in the area. The results suggest that the Alboran area may be inhabited by animals differentiated from the rest of the Mediterranean basin as a result of habitat features.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The bottlenose dolphin is widely distributed in temperate and tropical waters worldwide. Morphologic differences have, in the past, led to the subdivision of the genus into different species (Hershkovitz 1966). Currently, three species, Tursiops truncatus, Tursiops aduncus and Tursiops australis (Charlton-Robb et al. 2006, 2011), are recognized with the occurrence of local subspecies (e.g. T. truncatus-ponticus in the Black Sea; Viaud-Martinez et al. 2008) and nearshore and offshore ecotypes for a number of geographic locations (Ross 1977, 1984; Walker 1981; Duffield et al. 1983; Ross and Cockcroft 1990; Van Waerebeek et al. 1990; Mead and Potter 1995). Pelagic forms of T. truncatus have been reported to range primarily between the 200 and 2,000-m isobaths (Wells et al. 1999). In the North-western Atlantic, this pelagic ecotype occurs mainly in waters beyond 34 km from shore and 34 m depth, while the coastal form occurs within 7.5 km from shore (Torres et al. 2003). In the Gulf of California, a distribution break was found around the 60-m isobath (Segura et al. 2006). In the Central–Eastern North Atlantic, no population structure was evident for either ecotype (Quérouil et al. 2007). Resident populations of T. truncatus exist around the Canary Islands and the Azores archipelago (Silva et al. 2008). Nevertheless, in the latter, photo-identification data suggest that resident individuals mix and interact with non-resident individuals rarely observed in the area (Silva et al. 2008). In the Mediterranean Sea, bottlenose dolphins (T. truncatus) are thought to belong to the coastal ecotype (Notarbartolo di Sciara G and Demma 2004; Gannier 2005) despite being regularly observed in deep waters near the continental slope (Forcada et al. 2004) or beyond the continental shelf (Bearzi et al. 2004; Ben Naceur et al. 2004).
The bottlenose dolphin (T. truncatus) is a highly vocal species that shows great plasticity in its communication signals (May-Collado and Wartzok 2008). In this study, we refer to whistle as unpulsed, narrow-band signals, lasting between 0.1 and 4 s. The acoustic frequency of whistles is usually modulated, showing distinct contours of the fundamental frequencies (Caldwell et al. 1990). The whistles of bottlenose dolphins (T. truncatus) have been classified by Caldwell et al. (1990) into signature whistles, stereotypic and individual-specific signals that are stable over time and are used for group cohesion and variant whistles produced in a variety of social contexts. Furthermore, Caldwell and Caldwell (1972) and Reiss and McCowan (1993) reported that bottlenose dolphins are able to spontaneously copy sounds from the environment, and Tyack (1986) showed that they can also copy the whistles of conspecifics. Since acoustic transmission and ambient noise conditions can be locally different, animals may change the frequency and temporal structure of signals in response to the acoustic environment to ensure the transfer of information (May-Collado and Wartzok 2008). Local conditions of the acoustic environments experienced by a population can be reflected in differing traits of the acoustic structure of whistles and contribute to their geographic variation (May-Collado and Wartzok 2008). Furthermore, geographic variation can be related to morphological differences in the structure of the vocal apparatus and in overall body size. The call parameter most affected by body size is minimum frequency (May-Collado et al. 2007).
Intra-specific variations in the acoustic parameters of whistles have been successfully used to distinguish populations of many odontocete species, particularly bottlenose dolphins (Wang et al. 1995; Jones and Sayigh 2002; Morisaka et al. 2005; Azzolin 2008; Baron et al. 2008; May-Collado and Wartzok 2008; Hawkins 2010). Here, we evaluated differences in the acoustic characteristics of the signals produced by bottlenose dolphins across the Central–Eastern North Atlantic and the Mediterranean Sea, and we tested population discriminability using whistles.
Methods
Study populations and data collection
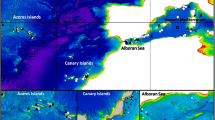
Recordings of bottlenose dolphin (T. truncatus) whistles were collected from four geographic locations: the Mediterranean Sea, the Azores archipelago, the Canary archipelago and the Bay of Biscay (Fig. 1 and Table 1).

Location of recordings analyzed for the Atlantic Ocean and Mediterranean Sea. Black dots represent the approximate position of sightings. Inserts show details of locations where sightings were closer. Depth contour of 200 m is shown in grey
The Mediterranean Sea is a semi-enclosed basin, located between 30° and 46° North and 6° West and 36° East, composed of two main sub-basins (eastern and western). The Mediterranean connects with the Atlantic Ocean through the Strait of Gibraltar. Data were only collected from the western sub-basin: in the Tyrrhenian, the Gulf of Lion (France), the Gulf of Vera (Spain; Eastern Almeria) and in the Alboran Sea (to the west of the Almerian-Oran barrier). Water depths can reach 3.8 km in the centre of the Tyrrhenian Sea. The Groupe de Recherche sur les Cétacés (GREC) provided the data from this location using either a mono towed hydrophone with Benthos AQ4 (in 1999) or a stereo towed hydrophone with the same elements (in 1998), with a linear flat response between 1 and 15 kHz ± 1 dB and between 15 and 30 kHz ± 3 dB (sensitivity of −156 dB re 1 V/μPa), a 29-dB pre-amplifier and 200 Hz high-pass filter. An external high-pass filter unit (Magrec Ltd.) set to 1 kHz was used on the hydrophone output to improve the quality of recordings.
The Azores archipelago is located between 36° and 40° North and 24° and 32° West and is composed of nine islands divided into three subgroups (western, central and eastern), extending about 600 km along a northwest–southeast axis. The islands are situated about 1,500 km from the Portuguese coast. The seabed around the islands is deep (around 1.50 km at 3 km off shore) with numerous scattered seamounts (Morato et al. 2008). The Department of Oceanography and Fisheries, Centre of IMAR of the University of the Azores (IMAR-DOP/UAç) and the International Fund for Animal Welfare-United Kingdom (IFAW) provided the recordings from the area using either an omnidirectional hydrophone (HTI-94-SSQ) with a linear flat response between 2 and 30 kHz ± 1 dB (sensitivity of −198 dB re 1 V/μPa) or a towed array with two hydrophones (Benthos AQ4).
The Canary Islands archipelago is located between 27° and 30° North and 13° and 19° West about 1,200 km from the Azores archipelago and 115 km west from the African coast. It is composed of seven main islands and extends 500 km. Water depth around the archipelago can reach more than 1 km at 1.8 km from the coast. The Society for the Study of Cetaceans in the Canary Archipelago (SECAC) obtained the recordings used in this study using a towed array with four elements: two hydrophones (Benthos AQ4) and two spherical ceramic hydrophone elements with a frequency response of ∼2–150 kHz (Seiche UK Ltd.) with a sensitivity for the front element of −161 dB re 1 V/μPa and the rear element of −158 dB re 1 V/μPa.
The Bay of Biscay is situated between 43° and 50° North and 1° and 10° West and is characterized by variable sea depths, ranging from the shallow continental shelf (less than 0.10 km) to the abyssal plain (greater than 4 km) with submarine canyons, seamounts and a steep continental slope. The width of the continental shelf varies from 110 to 185 km in the northern part of the bay (up to 45° N) to 46 km in the southern part and is as narrow as 5.5 km at the latitude of the Capbreton trough. The IFAW provided data from this location with the same instruments used in the Azores area. We only used recordings for which the species was confirmed visually and when it was visually certain that no other odontocetes were present in the area.
Sound analysis
We analyzed recordings by creating spectrograms in CoolEdit 2000 (Syntrillium Software, USA; Blackmann-Harris window; 256–512 band resolution; 2048 FFT size). We sampled all data at 48 kHz except for a few Mediterranean recordings collected at 44.1 kHz for which none of the maximum frequencies was over Nyquist nor frequency parameters or harmonics of the signals presented overturned contours. Each extracted sound was classified by assigning a signal quality index from zero (weak or overlapped with other sounds) to three (good signal-to-noise ratio and definition of the contour). When the gap between consecutive whistles was larger than 200 ms, these were analyzed as individual whistles (Bazua-Duran and Au 2002).
Only whistles classified as two or three (with the highest intensity) were used in the analysis in order to avoid using sounds of groups outside the visual range. From each whistle contour, ten parameters were measured manually following the method adopted by Oswald et al. (2003, 2007), Azzolin (2008) and Papale et al. (2013). These included duration, beginning frequency, end frequency, minimum frequency, maximum frequency, number of inflection points (mathematic definition in sine function of a change from positive to negative or negative to positive slope), steps (a rapid discontinuous change in frequency), number of minima in the contour and number of maxima in the contour (relative maximal and minimal points in the whistle contour) (Fig. 2). We also calculated the frequency range (maximum frequency–minimum frequency). Since we could not know the emitter, we considered on the whole dataset a mean of four sounds per animal per sighting. Furthermore, to avoid overestimation of the most repeated whistle structure due to the occurrence of possible signature whistles (Caldwell et al. 1990; Sayigh et al. 1990, 1998; Janik et al. 1994; Tyack 1997; Janik and Slater 1998; Janik 2000; Fripp et al. 2005) or mimicry between individuals, the contribution to the entire data set from signals with contour similar to another one was not allowed to exceed 14 %. To prevent any type of statistical bias due to this percentage, we randomized ten times the new dataset and compared 80 % of the data contained in each randomized dataset. Since we obtained different results only for the frequency range, parameter strictly related to the maximum and minimum frequencies, we decided not to consider it in the analysis.

Sample spectrogram representing a bottlenose dolphin whistle. Parameters manually measured for each whistle are shown: signal duration, beginning frequency, end frequency, minimum frequency, maximum frequency, the number of inflection points, the number of steps and the number of relative minima and relative maxima in the contour. Frequency range was calculated as maximum frequency minus minimum frequency
Data analysis
For each parameter, within- and between-basin coefficients of variation (CVs) were calculated as the ratio of the standard deviation to the mean (using all whistles for within-basin calculations and the mean value for each location for between-basin calculations) and expressed as a percentage (Lehner 1998). In order to evaluate which parameters are more likely to contribute to differences between whistles from different locations (the Mediterranean, the Azores, the Canary Islands and the Bay of Biscay), we compared inter-area CVs and intra-area CVs. The statistical software package PASW Statistics 18.0 (SPSS Institute Inc., Chicago, IL, USA) was used to create descriptive statistics (mean and standard deviation). Since the data were not normally distributed, we used the Mann–Whitney non-parametric test to determine whether and which whistle parameters varied between areas. We performed a discriminant function analysis (DFA) using the mean values for each sighting to determine whether whistles recorded could be correctly classified to the sampling areas. In this case, all the assumptions of the DFA were met. The leave-one-out procedure (Lachenbruch and Mickey 1968) was then used for cross-validation. Unfortunately, the sample from the Bay of Biscay was only represented by a single sighting, so, in view of the possibility of bias due to the homogeneity of signals in a short period and a single group, the recording was not considered in univariate and discriminant function analyses. Finally, we performed a hierarchical cluster analysis (using the within groups average linkage method) with the mean values for each sighting to classify them into the four study locations: Azores, Bay of Biscay, Canaries and Mediterranean Sea. For all of the multivariate statistics, we did not consider frequency range as a predictor variable due to its relationship with maximum and minimum frequency parameters.
Results
Study effort
For the Mediterranean Sea, we analyzed 3.06 h of recordings from which we extracted 577 whistles. For the statistical analysis, we considered 207 good quality sounds that originated from five sightings. For the Canary archipelago, we analyzed 2.25 h and extracted 186 whistles. We considered 94 sounds belonging to three sightings. For the Azores archipelago, 5.32 h from 20 sightings were investigated. We extracted 866 whistles and 352 of them were analyzed. In the Bay of Biscay, 0.18 h of recordings from one sighting were collected, from which 94 whistles were extracted and analyzed (Table 1).
Whistle variation between the Atlantic Ocean and the Mediterranean Sea
Parameters related to signal frequency were significantly higher in the Atlantic Ocean than in the Mediterranean, especially the beginning (Mann–Whitney test N = 747, Z = −6.03, P < 0.001), minimum (Z = −4.07, P < 0.001) and maximum (Z = −3.95, P < 0.001) frequencies. Mean values of signal modulation parameters, such as number of inflections and number of minima, were significantly lower in the Atlantic Ocean (Z = 5.20, P < 0.001; Z = 2.95, P < 0.001). The number of steps, maxima, end frequency and signal duration did not show significant differences between the basins (Z = −0.68, P = 0.49; Z = −0.63, P = 0.53; Z = −1.59, P = 0.11; Z = −1.49, P = 0.13; Table 2). The sightings could be correctly classified using DFA for 79.3 % of cross-validated cases (Table 3). The parameters that contributed to the classification were end frequency (coefficient = 0.89) and number of inflection points (coefficient = 0.68).
Whistle variation within and between areas
We performed a hierarchical cluster analysis using the mean values for the parameters for each sighting. The cluster analysis grouped three sightings from the Mediterranean Sea with one from the Canary archipelago. Three sightings from the Azores were also clustered separately from the other sightings from the region. The rest of the Atlantic sightings (2 from the Canaries, 17 from the Azores and 1 from the Bay of Biscay) were grouped together with the two sightings from the Alboran Sea (Fig. 3). The DFA performed using the mean values of the parameters for each sighting confirmed the results of the hierarchical cluster analysis. In this case, we excluded the Bay of Biscay because it contributed only one sighting. From the scatter plot of the analysis, it was possible to graphically identify one group encompassing the Azores, two sightings from the Alboran Sea (Mediterranean basin) and two from the Canaries and another group with the rest of the Mediterranean and Canarian sightings (Fig. 4).

Dendrogram of the hierarchic cluster analysis performed using mean values of each parameter for the sightings

Scatter plot of the discriminant function analysis performed using the mean values of each parameter for the sightings (Azores Islands, 20 sightings; Canary Islands, 3 sightings; Mediterranean Sea, 5 sightings)
Inter-area CVs of frequency parameters were generally lower, especially when compared with corresponding intra-area values (Table 2). Maximum frequency and range of frequency had the lowest inter-area CVs. The inter-area CVs for number of inflection points and steps (CV = 36.18, 42.25) were nearly double those for other modulation parameters and had only slightly higher intra-area CVs.
In order to evaluate the differences found between the Alboran Sea sightings and the rest of the Mediterranean (Eastern Almeria), we compared the mean values of the parameters. Signal duration in the Alboran Sea was significantly longer (Mann–Whitney test N = 207, Z = 3.55, P < 0.001), while beginning, end and minimum and maximum frequencies were lower than in the rest of the Mediterranean (Z = −3.64, P < 0.001; Z = −7.52, P < 0.001; Z = −6.22, P < 0.001; Z = −4.98, P < 0.001, respectively). In particular, the mean value of the end frequency parameter in the Alboran Sea was almost half the other Mediterranean sounds. Thus, this explains the differences among the parameter comparison for which there is no significant difference in end frequency and the DFA, where end frequency is the most important parameter to discriminate between Atlantic and Mediterranean. The number of inflection points and number of maxima were double or higher (Z = 2.52, P = 0.01; Z = 5.29, P < 0.001; Table 4). The number of steps and the number of minima (Z = −0.82, P = 0.41; Z = 1.73, P = 0.08) did not show any variation.
Although the Atlantic sightings could be grouped together, as evidenced by the DFA, the heterogeneity highlighted in the cluster analysis within the Central–Eastern North Atlantic led us to evaluate the variation of the parameters in particular between the Canary archipelago and the Azores islands. Significant differences were found in parameters related to frequency (Mann–Whitney test: N = 446, beginning frequency Z = −2.05, P < 0.04; end frequency Z = −6.09, P < 0.001; minimum frequency Z = −3.63, P < 0.001) and signal duration (Z = 4.23, P < 0.001). Other parameters that also showed significant variation between the locations were number of inflection points (Z = 3.64, P < 0.001), steps (Z = 6.37, P < 0.001), minima (Z = 2.83, P < 0.001) and maxima (Z = 2.64, P < 0.001). Maximum frequency did not show any differences (Z = −1.34, P = 0.18).
Discussion
Given the complexity of the genetic and ecological characteristics of bottlenose dolphins (Hoelzel et al. 1998), two scenarios can be put forward to interpret the variability observed in the communication sounds of the species in the Central–Eastern North Atlantic and the Mediterranean Sea:
-
1
No variation exists within the Central–Eastern North Atlantic, but a difference exists between the Atlantic Ocean and the Western Mediterranean Sea consistent with the partial isolation proposed by genetic studies, which have identified a single large population for each basin (Natoli et al. 2005), although with some gene flow between the Atlantic Ocean and the Western Mediterranean.
-
2
Significant differences exist between the characteristics of the signals of the locations of the same basin due to local conditions of the acoustic and social environments of resident individuals.
Unfortunately, data about the social (number of specimen per group, site fidelity, associated behaviour to whistles), ecological and physical environment (natural and anthropogenic noise, bathymetry, etc.) were not available for every site, and we could not assess the effect of these factors, but based on our acoustic results, we suggest that both scenarios coexist in the study area.
Our results confirm the existence of a significant divergence between the Central–Eastern North Atlantic and the Mediterranean Sea. About 79 % of the sightings were correctly assigned to one of the two basins based on frequency and modulation parameters of the whistles. Furthermore, within the Atlantic Ocean, the sightings can be grouped together (both with DFA and cluster analysis) with the exception of one sighting from the Canary Islands that clusters with the Mediterranean Sea recordings. Our acoustic results suggest that bottlenose dolphins occurring in North Atlantic pelagic waters belong to a large oceanic population consistent with the results reported by genetic studies. Quérouil et al. (2007) showed that bottlenose dolphins inhabiting the waters around the Azores and the island of Madeira have high gene flow, lack population structure within and between areas and are more similar to the pelagic populations of the Western North Atlantic than to dolphins from the Eastern Atlantic or the Mediterranean. Unlike coastal populations, oceanic bottlenose dolphins maintain high levels of gene flow and genetic diversity (Natoli et al. 2004, Quérouil et al. 2007). Furthermore, in the Azores archipelago, Silva et al. (2008) reported the absence of habitat partitioning between resident and non-resident dolphins. In the Canary Islands, bottlenose dolphins do not seem to be island associated, but moved between several islands of the archipelago (Castrillón et al. 2011; Tobeña et al. 2011). This suggests a situation similar to the Azores Islands. Nevertheless, although sightings could be grouped together, large heterogeneity was found in the Central–Eastern North Atlantic, where differences in acoustic parameters may represent local adaptations to the acoustic and social environments.
Furthermore, the coefficients of variation showed a high variability within each location, especially for the Canary Islands and the Mediterranean Sea. In these locations, the acoustic sample came from just a few sightings; nevertheless, results from the DFA and the hierarchical cluster analysis confirmed the variability highlighted, suggesting differences in the samples of both locations and the need for a more detailed study investigating micro-geographic variation.
Within the Mediterranean, sounds recorded from the Alboran Sea were significantly different from the recordings made in the Gulf of Vera, the Provencal and the Tyrrhenian Sea. The Alboran Sea whistles exhibited significantly lower frequency parameters and higher signal duration and modulation. Our acoustic results therefore suggest the presence of two different groups in the Mediterranean basin. Furthermore, although the mean values of the parameters from this area varied from those of Atlantic Ocean, the frequency parameters from the Alboran Sea were more similar to those from the Azores Archipelago. These results suggest that the Alboran Sea may be an area that is ecologically distinct from the rest of the Mediterranean and is perhaps a zone of transition between the Mediterranean and the Atlantic Ocean. Castellote et al. (2012) reported that fin whale (Balaenoptera physalus) calls detected in the Alboran basin and the Strait of Gibraltar were more similar to calls recorded in the Azores than to calls recorded elsewhere in the Western Mediterranean. These authors suggest that North Atlantic fin whales cross the Strait of Gibraltar and enter the Mediterranean Sea, but do not venture further than the Alboran Sea. Similarly, our recordings from the Alboran area are different to the rest of the Mediterranean, allowing us to assume that within their distribution range Mediterranean bottlenose dolphins have more than one evolutionary unit (considered as a distinct local population within a species that has different behavioural and phenological traits and thus harbours enough genetic uniqueness to warrant its own management and conservation). Furthermore, since the closest recording was collected in the Gulf of Vera (40 km east of the Alboran Sea), the possible limits to the distribution of Alboran animals may be at the Eastern end of the Alboran Sea, where an interchange zone could be present but not picked up by our sampling. The oceanographic features of the area, represented by the Almeria-Oran front, have already been suggested as a barrier to the movement of some species that leads to the creation of local populations of prey and their predators (Natoli et al. 2005). Bottlenose dolphins show genetic differentiation on either side of this front (Natoli et al. 2005), which is consistent with the acoustic results from our study.
In the Gibraltar area, the bottlenose dolphin population is considered strictly resident (Chico et al. 2011): in 2008, after 9 years of study, the re-sighting rate was found to be 90 %. A recent genetic study identifies individuals from the area as a pelagic population (Louis et al. 2013). Therefore, the Alboran basin may be inhabited by animals differentiated from the rest of the Mediterranean as a result of distinct habitat features, for example the presence of seamounts scattered through the whole area and currents coming from the Atlantic Ocean. The similarity between the oceanographic features of the Atlantic and the Almerian barrier suggests that an offshore population in the area could explain the acoustic relationship with the population inhabiting the Central–Eastern North Atlantic. This interpretation has important conservation implications since it suggests the presence of at least two different evolutionary units in the Mediterranean basin. Nevertheless, more data are needed to get new insights into the variability within the Mediterranean Sea especially where our relatively small sample size identified the possibility of a considerable acoustic difference.
The results reported here have value for the management of the species in the areas considered. Together with data from genetic studies, they provide a basis for defining bottlenose dolphin population ranges and give guidance to efforts aimed at defining conservation stocks. Despite common bottlenose dolphins' vocalizations being characterized by features under different selective forces and influenced by vocal production learning, their variation can be considered a proxy for the differentiation of evolutionary units that show genetic variation.
References
Azzolin M (2008) Acoustic identification of Mediterranean odontocetes as a prerequisite for their passive acoustic monitoring. Ph.D. Thesis, University of Torino, Italy
Baron SC, Marinez L, Garrison P, Keith KO (2008) Differences in acoustic signals from delphinids in the western North Atlantic and northern Gulf of Mexico. Mar Mamm Sci 24:42–56
Bazua-Duran C, Au WWL (2002) The whistles of Hawaiian spinner dolphins. J Acoust Soc Am 112:3064–3072
Bearzi G, Holcer D, di Notarbartolo Sciara G (2004) The role of historical dolphin takes and habitat degradation in shaping the present status of northern Adriatic cetaceans. Aquat Conserv Mar Freshw Ecosys 14:363–379
Ben Naceur L, Gannier A, Bradai MN, Drouot V, Bourreau S, Laran S, Khalfallah N, Mrabet R, Bdioui M (2004) Recensement du grand dauphin Tursiops truncatus dans les eaux tunisiennes. Bulletin de l'Institute National des Sciences et Technologies de la Mer de Salammbô 31:75–81
Caldwell MC, Caldwell DK (1972) Vocal mimicry in the whistle mode by an Atlantic bottlenosed dolphin. Cetology 9:1–8
Caldwell MC, Caldwell DK, Tyack PL (1990) Review of the signature-whistle hypothesis for the Atlantic bottlenose dolphin. In: Leatherwood S, Reeves RR (eds) The bottlenose dolphin. Academic, San Diego, pp 199–235
Castellote M, Clark CW, Lammers MO (2012) Fin whale (Balaenoptera physalus) population identity in the western Mediterranean Sea. Mar Mamm Sci 28:325–344
Castrillón J, Reyes M, Ruiz L, Pérez-Gil E, Brederlau B, Neves S, Pérez-Gil M, Tejedor M, Servidio A, Herrero R, Lucas T, Urquiola E, Martín V (2011) Simultaneous survey of bottlenose dolphin (Tursiops truncatus) in five special areas of conservation in Canary Islands. 25th Conference of the European Cetacean Society, Cadiz, Spain
Charlton-Robb K, Taylor AC, McKechnie SW (2006) A note on divergent mtDNA lineages of bottlenose dolphins from coastal waters of Southern Australia. J Cet Res Manag 8:173–179
Charlton-Robb K, Gershwin LA, Thompson R, Austin J, Owen K, McKechnie SW (2011) A New Dolphin Species, the Burrunan Dolphin Tursiops australis sp. nov., Endemic to Southern Australian Coastal Waters. PLoS One 6:E24047
Chico C, Jimenez-Torres C, Perez S, Verborgh P, Gauffier P, Esteban R, Gimenez J, Santos ME, De Stephanis R (2011) Survival rate, abundance and residency of bottlenose dolphin (Tursiops truncatus) in the strait of Gibraltar. 25th Conference of the European Cetacean Society, Cadiz, Spain
Duffield DA, Ridgeway SH, Cornell LH (1983) Hematology distinguishes coastal and offshore forms of dolphins (Tursiops). Can J Zool 61:930–933
Forcada J, Gazo M, Aguilar A, Gonzalvo J, Fernandez-Contreras M (2004) Bottlenose dolphin abundance in the NW Mediterranean: addressing heterogeneity in distribution. Mar Ecol Prog Ser 275:275–287
Fripp D, Owen C, Quintana-Rizzo E, Shapiro A, Buckstaff K, Jankowski K, Wells R, Tyack PL (2005) Bottlenose dolphin (Tursiops truncatus) calves appear to model their signature whistles on the signature whistles of community members. Anim Cogn 8:17–26
Gannier A (2005) Summer distribution and relative abundance of delphinids in the Mediterranean sea. Rev Ecol (Terre Vie) 60:223–238
Hawkins ER (2010) Geographic variations in the whistles of bottlenose dolphins (Tursiops aduncus) along the east and west coasts of Australia. J Acoust Soc Am 128:924–935
Hershkovitz P (1966) A catalogue of living whales. Bull US Nat Mus 246:1–259
Hoelzel AR, Potter CW, Best PB (1998) Genetic differentiation between parapatric ‘nearshore’ and ‘offshore’ populations of the bottlenose dolphin. Proc R Soc London B 265:1177–1183
Janik VM (2000) Whistle matching in wild bottlenose dolphin (Tursiops truncatus). Science 289:1355–1357
Janik VM, Slater PJB (1998) Context-specific use suggests that bottlenose dolphin signature whistles are cohesion calls. Anim Behav 56:829–838
Janik VM, Dehnhardt G, Todt D (1994) Signature whistle variations in a bottlenosed dolphin, Tursiops truncatus. Behav Ecol Sociobiol 35:243–248
Jones GJ, Sayigh LS (2002) Geographical variation in rates of vocal production of free-ranging bottlenose dolphins. Mar Mamm Sci 18:374–393
Lachenbruch PA, Mickey MR (1968) Estimation of error rates in discriminant analysis. Technometrics 10:1–11
Lehner PN (1998) Handbook of ethological methods, 2nd edn. Cambridge University, Cambridge, p 672
Louis M, Guinet C, Lucas T, Viricel A, Peltier H, Alfonsi E, Berrow S, Brownlow A, Covelo P, Dabin W, Deaville R, Gally F, Gauffier P, Penrose R, Silva M, Simon-Bouhet B (2013) Population genetics of bottlenose dolphins in the North East Atlantic: a pelagic versus coastal segregation. 27th Conference of the European Cetacean Society, Setubal, Portugal
May-Collado LJ, Wartzok D (2008) A comparison of bottlenose dolphin whistles in the Atlantic Ocean: factors promoting whistle variation. J Mammal 89:1229–1240
May-Collado LJ, Agnarsson I, Wartzok D (2007) Re-examining the relationship between body size and tonal signals frequency in whales: a comparative approach using a novel phylogeny. Mar Mamm Sci 23:524–552
Mead JG, Potter CW (1995) Recognizing two populations of the bottlenose dolphin (Tursiops truncatus) of the Atlantic coast of North America: morphologic and ecologic considerations. IBI Rep 5:31–44
Morato T, Machete M, Kitchingman A, Tempera F, Lai S, Menezes G, Pitcher TJ, Santos RS (2008) Abundance and distribution of seamounts in the Azores. Mar Ecol Prog Ser 357:17–21
Morisaka T, Shinohara M, Nakahara F, Akamatsu T (2005) Geographic variations in the whistles among three Indo-Pacific bottlenose dolphin Tursiops aduncus populations in Japan. Fish Sci 71:568–576
Natoli A, Peddemors VM, Hoelzel AR (2004) Population structure and speciation in the genus Tursiops based on microsatellite and mitochondrial DNA analyses. J Evol Biol 17:363–375
Natoli A, Birkun A, Aguilar A, Lopez A, Hoelzel AR (2005) Habitat structure and the dispersal of male and female bottlenose dolphins (Tursiops truncatus) based on microsatellite and mitochondrial DNA analyses. Proc R Soc London B 272:1217–1226
Notarbartolo di Sciara G, Demma M (2004) Guida dei mammiferi marini del Mediterraneo. In: Franco Muzzio (Ed) Scienze naturali Padova
Oswald JN, Barlow J, Norris TF (2003) Acoustic identification of nine delphinid species in the eastern tropical pacific ocean. Mar Mamm Sci 19:20–37
Oswald JN, Rankin S, Barlow J, Lammers MO (2007) A tool for real-time acoustic species identification of delphinid whistles. J Acoust Soc Am 122:587–595
Papale E, Azzolin M, Cascão I, Gannier A, Lammers MO, Martin VM, Oswald J, Perez-Gil M, Prieto R, Silva MA, Giacoma C (2013) Geographic variability in the acoustic parameters of striped dolphin’s (Stenella coeruleoalba) whistles. J Acous Soc Am 133(2):1126–1134
Quérouil S, Silva MA, Freitas L, Prieto R, Magalhaes S, Dinis A, Alves F, Matos JA, Mendonca D, Hammond PS, Santos RS (2007) High gene flow in oceanic bottlenose dolphins (Tursiops truncatus) of the North Atlantic. Conserv Genet. doi:10.1007/s10592-007-9291-5
Reiss D, McCowan B (1993) Spontaneous vocal mimicry and production by bottlenose dolphins (Tursiops truncatus): evidence for vocal learning. J Comp Psychol 107:301–312
Ross GJB (1977) The taxonomy of bottlenose dolphins, Tursiops species, in South African waters, with notes on their biology. Ann Cape Provincial Mus 11:135–194
Ross GJB (1984) The smaller cetaceans of the southeast coast of southern Africa. Ann Cape Provincial Mus 15:173–410
Ross GJB, Cockcroft VC (1990) Comments on Australian bottlenose dolphins and the taxonomic status of Tursiops aduncus (Ehrenberg, 1832). In: Leatherwood S, Reeves RR (eds) The bottlenose dolphin. Academic, New York, pp 101–128
Sayigh LS, Tyack PT, Wells RS, Scott MD (1990) Signature whistles of free-ranging bottlenose dolphins Tursiops truncatus: stability and mother–offspring comparisons. Behav Ecol Sociobiol 26:247–260
Sayigh LS, Tyack PL, Wells RS, Solows AR, Scott MD, Irvine AB (1998) Individual recognition in wild bottlenose dolphins: a field test using playback experiments. Anim Behav 57:41–50
Segura I, Rocha-Olivares A, Flores-Ramırez S, Rojas-Bracho L (2006) Conservation implications of the genetic and ecological distinction of Tursiops truncatus ecotypes in the Gulf of California. Biol Conserv 133:336–346
Silva MA, Prieto R, Magalhães S, Seabra MI, Santos RS, Hammond PS (2008) Ranging patterns of bottlenose dolphins living in oceanic waters: implications for population structure. Mar Biol 156:179–192
Tobeña M, Escanez A, Rodriguez Y, Lopez C, Aguilar N (2011) Inter-island movement of bottlenose dolphins (Tursiops truncatus) in the Canary islands: implications for conservation and SAC design. 25th Conference of the European Cetacean Society Cadiz, Spain
Torres LG, Rosel PE, D'Agrosa C, Read AJ (2003) Improving management of overlapping bottlenose dolphin ecotypes through spatial analysis and genetics. Mar Mamm Sci 19:502–514
Tyack PL (1986) Whistle repertoires of two bottlenosed dolphins, Tursiops truncatus: mimicry of signature whistles? Behav Ecol Sociobiol 18:251–257
Tyack PL (1997) Development and social functions of signature whistles in bottlenose dolphins Tursiops truncatus. Bioacoustics 8:21–46
Van Waerebeek K, Reyes JC, Read AJ, McKinnon J (1990) Preliminary observations of bottlenose dolphins from the Pacific coast of South America. In: Leatherwood S, Reeves RR (eds) The bottlenose dolphin. Academic, New York, pp 143–154
Viaud-Martinez KA, Brownell RL, Komnenou A, Bohonak AJ (2008) Genetic isolation and morphological divergence of Black Sea bottlenose dolphins. Biol Conserv 141:1600–1611
Walker WA (1981) Geographic variation in morphology and biology of bottlenose dolphins (Tursiops) in the eastern North Pacific. NOAA/NMFS Southwest Fisheries Science Centre Administrative Report no. LJ-81-3c
Wang D, Wursig B, Evans WE (1995) Whistles of bottlenose dolphins: comparisons among populations. Aquat Mamm 21:65–67
Wells RS, Rhinehart HL, Cunningham P, Whaley J, Baran M, Koberna C, Costa DP (1999) Long distance offshore movements of bottlenose dolphins. Mar Mamm Sci 15:1098–1114
Acknowledgments
The authors are grateful to IFAW for its contribution in the data collection. Without its support, this project could not have been accomplished. Data collection and processing in the Azores was funded by Fundação para a Ciência e a Tecnologia (FCT), Direcção Regional Para a Ciência, Tecnologia e Comunicações (DRCTC), FEDER, the Competitiveness Factors Operational (COMPETE), QREN European Social Fund and Proconvergência Açores Program through research projects PTDC/MAR/74071/2006 and M2.1.2/F/012/2011. The authors acknowledge funds provided by FCT to LARSyS Associated Laboratory and IMAR-University of the Azores/the Thematic Area E of the Strategic Project (OE and compete) and by the DRCTC–Government of the Azores pluriannual funding. MAS was supported by an FCT postdoctoral grant (SFRH/BPD/29841/2006), IC and RP were supported by FCT doctoral grants (respectively SFRH/BD/41192/2007 and SFRH/BD/32520/2006). RP received a research grant from the Azores Regional Fund for Science and Technology (M3.1.5/F/115/2012). Data collection by SECAC was funded by the EU LIFE programme—project LIFE INDEMARES (LIFE 07/NAT/E/000732)—and the Fundación Biodiversidad under the Spanish Ministry of Environment, Rural and Marine Affairs (project ZEC-TURSIOPS).
Conflict of interest
None of the authors has any conflict of interest with the contents of the manuscript.
Ethical standards
The work has been carried out without putting at risk endangered populations, species or habitats in agreements with the “Guidelines for the Use of Animals in Research.”
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Papale, E., Azzolin, M., Cascão, I. et al. Acoustic divergence between bottlenose dolphin whistles from the Central–Eastern North Atlantic and Mediterranean Sea. acta ethol 17, 155–165 (2014). https://doi.org/10.1007/s10211-013-0172-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10211-013-0172-2