
Abstract
Very little is known about the ecology of common bottlenose dolphins (Tursiops truncatus) living in oceanic waters. This study investigated the ranging and residence patterns of bottlenose dolphins occurring in the Azores (Portugal), the most isolated archipelago in the North Atlantic. Data were collected during standardized boat-based surveys conducted over a 6-year period in an area of approximately 5,400 km2 (main study area). To investigate the extent of movements of individual animals, non-systematic surveys were also conducted outside this area. Only 44 individuals out of 966 identified were frequently sighted within and between years. The remaining individuals were either temporary migrants from within or outside the archipelago, or transients. Resident dolphins showed strong geographic fidelity to the area. Long-distance movements (of almost 300 km), consistent with foraging or exploratory trips, were observed among non-resident dolphins. Home range size was estimated for 31 individuals sighted ≥10 times. Range areas of these dolphins varied in size and location, but considerable overlap was observed in the areas used, suggesting the absence of habitat partitioning between resident and non-resident dolphins. Estimates of home range size of bottlenose dolphins in the Azores were found to be considerably larger than those previously reported for this species. It is hypothesized that dolphins living in the Azores carry out extensive movements and have large home ranges in response to the lower density and patchy distribution of prey compared to other areas. The extensive ranging behaviour and the lack of territoriality provide an opportunity for interbreeding between dolphins associated with different islands, thus preventing genetic differentiation within the population of the Azores.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The analysis of ranging patterns is crucial to understanding several aspects of the ecology, dynamics, social structure and evolutionary trajectory of a population. Knowledge of individual patterns of space use may be used to identify residency and territoriality (Sandell 1989) and can provide important insights into the spatial and temporal distribution of resources (Damuth 1981). The movements of individuals also have fundamental effects on the genetic structure of populations (Wiens 1976) by providing the opportunities for gene flow to occur between different areas.
The concept of a home range to describe an individual animal’s area usage was first introduced by Burt (1943), who defined it as “the area traversed by an individual in its normal activities of food gathering, mating and caring for the young”. Inter- and intra-specific variation in home range size have been mainly explained as a function of body size/mass, diet, climate, competition, predation and reproductive strategies (McNab 1963; Damuth 1981; Swihart et al. 1988; Sandell 1989). Among these, variation in prey availability is considered one of the most important factors. In less productive habitats animals should maintain larger home ranges because they need to range further to find enough food (Sandell 1989). Similarly, animals preying on patchily distributed resources are predicted to have larger home ranges because they must travel further in order to find adequate food patches (Ford 1983). In practice, an increase in home range size with decreasing food availability/density seems to be a general result in mammals (Sandell 1989).
Although data on whole home ranges is scarce for most cetacean species, several studies reported a strong correlation between cetacean movement patterns and patterns of distribution and abundance of their prey (reviewed in Stevick et al. 2002). Differences in ranging patterns among populations of common bottlenose dolphins (hereafter called bottlenose dolphin) (Tursiops truncatus) occurring in different areas have also been related to the availability of food resources. For example, the Sarasota Bay dolphins are long-term, year-round residents with a home range of about 125 km2 and show strong site fidelity to the area (Scott et al. 1990). In South Carolina, resident dolphins have even smaller home ranges, show moderate levels of mobility and are never encountered outside estuarine areas (Gubbins 2002). These authors propose that the relatively abundant and predictable food resources in these areas may sustain a resident population year-round. On the other hand, certain habitats may provide only temporary, less abundant prey resources, and dolphins are forced to range over long distances in search for new food patches (Ballance 1992; Defran et al. 1999). These results are in agreement with theoretical predictions and studies on other taxonomic groups. However, nearly all the information available comes from inshore or coastal populations and the ranging behaviour of bottlenose dolphins living in oceanic waters remains largely unknown. The aim of this study is to fill in this gap, by studying the ranging behaviour of bottlenose dolphins living around the oceanic islands of the Azores.
The Azores archipelago is the most isolated archipelago in the North Atlantic, located about 1,500 km away from the nearest continental margin. The Gulf Stream and North Atlantic and Azores currents, and the dynamic Azores Front, are responsible for the seasonal and inter-annual dependent complex pattern of ocean circulation that characterizes the region, and results in the high salinity, high temperature and low-nutrient regime waters (Santos et al. 1995). Prominent topographic features in the ocean, such as islands and seamounts, are often associated with higher levels of biological productivity and diversity than open waters (Palacios 2002; Genin 2004). Oceanic islands are responsible for the development of localized upwellings, eddies and convergence zones, which in turn may cause enhanced primary productivities and promote biomass accumulation at specific sites (Caldeira et al. 2002; Palacios 2002). In addition, the islands may act as a barrier to the horizontal dispersal of zooplankton and larval/juvenile fish that tends to become entrapped in their vicinity (Palacios 2002), thereby increasing feeding opportunities for predators.
Although the influence of the islands on fine-scale oceanographic processes remains unknown, it has long been realized that amid this open ocean oligotrophic region, the waters in the vicinity of the Azores represent an “oasis” of biological productivity that attracts numerous pelagic organisms (Santos et al. 1995). If the waters around the islands provide suitable habitat and enough food resources, we expect bottlenose dolphins to show limited ranging behaviour and strong site fidelity to the area. If, on the other hand, food resources are scarce or available only sporadically, then dolphins are predicted to range over long distances in search of adequate food. Ultimately, the ranging behaviour of bottlenose dolphins may influence the structure of the population in the area, by providing the opportunity for dolphins from different islands to mix and genetic interchange to occur.
We used photo-identification data collected over a 6-year period to study the ranging patterns of bottlenose dolphins living around the Azores archipelago. Quantitative data on site fidelity, movements and home ranges of individually recognizable dolphins were examined to investigate whether bottlenose dolphin’s behavioural patterns are consistent with those of a coastal resident population, or of an oceanic transitory population.
Materials and methods
Study area
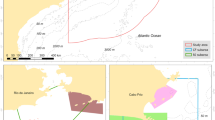
The Azores Archipelago (Portugal) is located in the middle of the Atlantic, between 37° and 41°N and 25° and 31°W, extending more than 600 km along a northwest-southeast trend and crossing the Mid-Atlantic Ridge (Santos et al. 1995). It consists of nine volcanic islands divided into three groups: eastern (comprising the islands of S. Miguel and Sta. Maria), central (islands of Faial, Pico, S. Jorge, Terceira and Graciosa) and western (islands of Flores and Corvo) (Fig. 1). The eastern and central groups of islands are approximately 230 km apart and the central and western groups are about 160 km apart. The bottom topography of the region is characterized by numerous shallow-water and emergent features (shoals, seamounts, islets and the islands) rising steeply from abyssal depths (>3,000 m), as well as deep-water ridges and submarine canyons (Fig. 2). Logistical reasons prevented equal survey effort within the central area and for the purpose of this study, the central group of islands was subdivided into two areas: main (comprising the islands of Faial, Pico and the channel between Pico and S. Jorge) and central (islands of Terceira, Graciosa and northern part of S. Jorge) (Fig. 1).

Map showing the location of the Azores in the Atlantic Ocean and the location of the four studied areas

Map of the study area showing the survey tracks (1999–2004) and the bathymetry
Boat surveys
Boat surveys were conducted in the main study area (approximately 5,400 km2) from March 1999 through October 2004. These surveys followed a pre-determined track, either alongshore at 1 km from the coast or in a zig-zag pattern up to 8 km from the islands, and these were designed to ensure as equal coverage as possible within the area. The survey route was selected based on the weather and sea conditions and time constraints on each day. An attempt was made to survey the main area at least twice a month between May and September and once a month during the remainder of the year. However, survey effort varied both within and among years. To investigate the extent of movements of individual animals, surveys were also conducted in the other three areas. Between 2002 and 2004, each of these secondary areas was visited twice for periods of 2–3 weeks. Surveys in the secondary areas were restricted to spring and summer months when better weather conditions were expected to occur. Searching effort was not equally distributed throughout these areas and was concentrated in regions where dolphins were more likely to be found or in the more sheltered locations.
Surveys were conducted from a 5.5 m rigid inflatable boat or from a 12 m fibreglass boat. During surveys, a steady speed of 16–22 km h−1 was maintained, while a minimum of three observers searched for dolphins and collected data on observation effort and weather and sea conditions. Surveys were only carried out in Beaufort sea-states ≤3. When dolphins were encountered, school size and composition and the initial time and location [determined by Global Positioning System (GPS)] were recorded. A ‘school’ was defined as all individuals within 100 m radius of each other (Irvine et al. 1981). An attempt was made to obtain several photographs of both sides of every individual present in the school. Photographs were taken with a Nikon F-90× autofocus camera equipped with a Nikkor AF 70–300 mm (f4-5.6) zoom lens, and using Kodak Elitechrome ISO 200 or Ektachrome Elite II ISO 200 colour slide film. Females were identified in the field by consistent association with a small calf during the course of a sighting (n = 21) (Mann and Smuts 1999), and in a few cases (n = 3) gender was assigned by visual inspection of the genital area. Sex was determined for other 64 dolphins through genetic analyses (see below). Dolphins were classified into broad categories—adults, subadults or calves—according to their size, colour, and behaviour (Mann and Smuts 1999). This classification was performed in the field while the individual was being photographed, and confirmed again through examination in the laboratory of the pictures taken. Calves were excluded from all the analyses performed because they usually do not possess enough marks to ensure their future recognition without error.
Beginning in April 2002, biopsy samples of adult and subadult dolphins were collected to investigate the genetic structure of the population. The biopsy sampling procedures and the methods and results of the genetic analyses were presented elsewhere (Quérouil et al. 2007) and in the present study we only used data on gender identification. For 64 dolphins that were simultaneously photographed and biopsied, sex was later determined through co-amplification of a short fragment of the male-specific SRY gene and a microsatellite fragment used as a PCR control for positive identification of females (see Quérouil et al. 2007). Once photographic data and biopsy samples had been collected, the dolphin school was abandoned and the survey resumed from that location.
Photo-identification procedures
Pictures obtained from each encounter were examined in the laboratory with an 8–20× binocular lens. Photographs were graded according to their focus, light and contrast, size of dorsal fin in relation to the frame and angle of dorsal fin. Only good quality photographs were used in this study. Individual animals were identified based primarily on the number and location of nicks and scars on their dorsal fins, but also on the scars and pigmentation pattern along the flanks (Würsig and Jefferson 1990). The best photographs of each dolphin were then compared with the best photograph of all previously identified individuals, and included in the catalogue as either a new identification or as a resighting of a known dolphin. Only individuals with sufficiently distinctive marks to allow future recognition were included in the dataset. If the number of individuals photo-identified in a given encounter was larger than the field estimate, the former value was used as the estimate of school size. Otherwise, the school size estimated in the field was used.
Residency and site fidelity in the main area
Sighting frequency, number of years observed, mean monthly sighting rate, and extent of movements were used to assess the degree of residency and fidelity of individual dolphins to the main area. For the remaining areas, the number of dolphin resightings and the temporal scale considered were judged insufficient to conduct these analyses. The dataset used to investigate residency and site fidelity included all dolphins seen at least once in the main area. The monthly sighting rate was calculated as the proportion of months a certain individual was seen in relation to the number of months surveyed during the years it was observed in the area. This value was then averaged across the years the individual was seen, resulting in a mean monthly sighting rate. This index therefore reflects the degree of fidelity during the periods when the individual frequented the area and is independent of the number of years it was seen. The mean monthly sighting rate varies between 0 and 1, the maximum value corresponding to an individual that was seen in all the months surveyed in the years it was observed in the area.
Movements
Linear distance between consecutive sightings of recognizable individuals was measured with the Animal Movement Analyst Extension of ArcView ® 3.2 (Hooge and Eichenlaub 1997) and used to assess the extent of movements and to evaluate differences in distance travelled by dolphins of different sex and age classes. There was a weak but significant correlation between distance and time elapsed between consecutive sightings of individuals (Pearson’s correlation, r = 0.206, P < 0.001). Thus, only sightings of individual dolphins made >31 days apart were analyzed, as we considered 1 month to be a time interval long enough to ensure independence between sightings.
Home ranges
Ranges of individual dolphins were calculated using Minimum Convex Polygons (MCP) (Mohr 1947) and the fixed kernel method (Worton 1989), available from the Animal Movement Analyst Extension of ArcView ® 3.2 (Hooge and Eichenlaub 1997). The MCP is the smallest convex polygon containing all the observed positions and the area within this polygon corresponds to the estimated home range size. The MCP is the oldest and most common home range estimator, and although it suffers from several biases (Kernohan et al. 2001) it was chosen for comparison purposes. The kernel is a probabilistic method that attempts to assess the animal’s utilization distribution (UD) within an area. Thus, instead of just reporting the size of the area used by the individual, kernel methods also assess the individual’s probability of occurrence at each point within its home range. Kernel methods have been found to be robust to a number of biases and generally to perform better than all the other estimators (Kernohan et al. 2001).
In the present study, the bandwidth value (which controls the width of individual kernels, determining the amount of smoothing applied to the data) used in the fixed kernel was calculated through the least squares cross validation, considered the most reliable and objective method for selecting the smoothing parameter (Seaman et al. 1999). Estimators may be critically affected by serial autocorrelation as the distance between consecutive positions decreases, leading to the underestimation of home range size (Kernohan et al. 2001). To attempt to ensure independence of sampling and decrease the bias from autocorrelation, multiple sightings from the same individual made on the same survey were eliminated from the dataset. In spite of this precaution, some degree of autocorrelation was still expected to occur. Therefore, Schoener’s ratio (ratio of the mean squared distance between successive observations and the mean squared distance from the centre of activity, Schoener 1981) was calculated for each individual and used to assess the amount of autocorrelation in the data and the potential effects on the estimates.
Before calculating home range size, extreme sightings of each individual were identified and removed using the harmonic mean outlier removal method (White and Garrott 1990). A bootstrap test was run to examine the increase in the MCP home range size with the increase in the number of locations used for each animal. For most of the animals analyzed, the area-observation curve approached the asymptote at ten sightings, indicating this value as the minimum number of sightings required to estimate the size of the home range. Location data from 31 individuals sighted ≥10 times were used to estimate the size of their home ranges, using both MCP and fixed kernel methods (after subtracting the area of landmasses from all the estimates).
Because MCP and kernel methods are known to behave differently when subject to the same source of bias, univariate general linear models were used to determine the effect of sample size and autocorrelation (indicated by Schoener’s ratio) on each of the estimators. The overall ranging area (MCP and kernel 95% UD) and core area (kernel 50% UD) calculated for adult and subadult dolphins were compared using the Mann–Whitney U test.
Spatial overlap between dolphin’s home ranges was estimated by measuring the size of the overlapping region of the kernel 95% UD of all possible pairs of dolphins using ArcView ® 3.2. Percentage of home range overlap between a pair of dolphins was calculated using the formula (R i,j/R i) × (R i,j/R j), where R i,j is the size of the area of overlap between dolphins i and j, and R i and R i are the total range sizes of dolphins i and j, respectively. To determine if the degree of space use sharing differed among dolphins of different sex, age and residence classes, the percentage of home range overlap between pairs of dolphins was compared using the Kruskal–Wallis test.
Results
Survey effort and sightings
In total, 353 surveys were conducted in the study area. Each survey lasted between 1 and 12 h, with an average of 4 h, and surveys in different areas were carried out with time intervals varying between 5 and 33 days. Most of the survey effort was concentrated in the main area, especially during spring and summer months (Table 1; Fig. 2). The western area was only surveyed 13 times in both years due to the poor weather and sea conditions in this region. Bottlenose dolphins were encountered during 42% of all surveys carried out in the main area, 57% in the eastern, 40% in the central and 31% in the western. Overall, 170 schools were photographed and 966 different individuals identified from the photographs. The number of dolphins identified in each year and area varied greatly but, as expected, largest numbers of dolphins were identified in the main study area (Table 1). Group size ranged from 1 to 110, with an average of 21.3 (±1.6 SE) animals. Fifty-one percent of the schools photographed ranged from 1 to 15 dolphins, 34% ranged from 16 to 40, and 15% of the schools had more than 40 animals. There were no significant differences in group size among areas (Kruskal–Wallis ANOVA, H = 3.689, P = 0.297, df = 3).
Residence and site fidelity in the main area
Of the 966 individuals identified in the Azores, 639 were observed at least once in the main area. Of these, 28 were calves and were excluded from all the analyses. The number of times each individual dolphin was observed in the main area varied considerably. Sighting frequencies ranged from 1 to 21 (median = 2.0), with most individuals being recorded only once (n = 215) or twice (n = 160). Only 5% of the dolphins were seen ≥10 times in the main area. The majority (57%) of the individuals was observed in a single year and several dolphins were seen repeatedly but in non-consecutive years. Only 22 dolphins were seen in the main area ≥5 years. These dolphins were seen throughout the year, without an evident seasonal pattern of occurrence.
Overall, the mean monthly sighting rate was low, ranging from 0.1 to 0.475 (0.160 ± 0.003 SE). On average, individuals were encountered in 16% of the months surveyed during the years they were recorded in the main area. There was no indication that males and females differed in their degree of residency or site fidelity (total sightings: U = 204.5, P = 0.387, n = 44; mean monthly sighting rate: U = 233.0, P = 0.842, n = 44). Similarly, there were no significant differences among dolphins from different age classes (total sightings: Z = 0.721, P = 0.471, n = 352; mean monthly sighting rate: Z = 1.637, P = 0.101, n = 352).
A strong association was observed between the number of years a dolphin was seen in the main area and its mean monthly sighting rate (weighted means ANOVA, F = 9.014, P < 0.0001, df = 5). This means that these dolphins not only showed between-year site fidelity but also used the area regularly throughout the year. Using the two variables together, dolphins were subsequently divided into two arbitrary groups: one group composed of 567 individuals sighted ≤3 years with mean monthly sighting rates averaging 0.16, and a second group comprising 44 dolphins observed at least 4 years and with an average mean monthly sighting rate of 0.23 (Fig. 3). The latter group was subsequently treated as the resident group in the main area, while the remaining dolphins were regarded as transients or occasional visitors. The two groups included dolphins of both sexes and age classes.

Relationship between the mean monthly sighting rate and the number of years dolphins were seen in the main area. Vertical bars correspond to 95% confidence intervals
There was a significant negative correlation between the number of years a given dolphin was observed in the main area and the maximum distance travelled by that animal (Pearson’s correlation, r = −0.165, P < 0.05) (Fig. 4). Large scale movements (>150 km) were only detected in dolphins observed in less than 2 years, and with the exception of two individuals, distances between consecutive sightings of dolphins sighted more than 2 years were less than 50 km.

Relationship between the maximum distance (km) between consecutive sightings and the number of years individual dolphins were seen in the main area
Movements
The average distance between consecutive sightings of recognizable individuals was 25.4 (±1.47 SE) km and approximately 72% of the movements recorded were less than 20 km. However, extensive movements (150–291 km) were also detected and these represented about 5% of the total. To investigate whether the wide-scale movements were carried out by a given class of individuals, distance between consecutive sightings was compared for dolphins of known sex and age class. There were no significant differences in the distribution of distances travelled by dolphins of different sex (D = 0.178, P > 0.1, n = 150) or age class (D = 0.143, P > 0.05, n = 595). Because surveys in the secondary areas were only conducted during spring and summer months, it was not possible to determine if these wide-scale movements occurred year-round or were restricted to a particular season. However, since the beginning of surveys in the secondary areas, long-distance movements were recorded every year.
Individuals were considered as belonging to the area where they were first seen. Approximately 7% of the 925 adult dolphins identified were encountered in more than one area during the study period (Table 2). However, the proportion of moving individuals (individuals first seen in one area and later photographed in a different area) was not independent of the area to which they belonged (χ 2 = 114.386, P < 0.0001, df = 3), with the central area recording the highest value (41%) (Table 2). The largest number of movements was recorded between the central and main areas, both within and between years, possibly because of the shortest distance separating them. No movements were ever detected between the eastern and western areas, the two most extreme groups of islands in the archipelago. Some dolphins were documented to repeatedly move back and forth between two areas, but there were no records of individuals moving to a third area.
In approximately 75% of the times only one moving individual was identified in the school and in 12.5% there were two individuals. However, on one occasion, a group of 11 moving individuals was reported, indicating some level of school cohesion. When moving to a different area, individuals did not remain alone or confined to their original groups but joined resident schools in the area. Except for two sightings involving pairs of individuals, dolphins from outside the area were always photographed together with dolphins already known in the area. Schools in which moving individuals were detected were significantly larger than those without moving individuals (Z = 2.510, P = 0.012, n = 113). However, there was no correlation between the number of moving individuals in a school and the school size (Spearman’s rank correlation, r = 0.118, P = 0.452, n = 43) or between the number of moving individuals and the number of individuals photo-identified in the school (Spearman’s rank correlation, r = 0.131, P = 0.401, n = 43). This means that moving individuals were consistently observed in larger schools, not because they comprised the majority of individuals in the school to which they moved, but because they tended to mix with already larger dolphin aggregations.
Home ranges
The mean MCP range size of the 31 dolphins was 182.0 km2, varying from 62.9 to 725.1 km2. The kernel method produced a mean 95% UD area of 437.2 km2 and a 50% UD core area of 86.4 km2 (Table 3). There was a significant correlation between the two estimators (Spearman’s rank correlation, r = 0.705, P < 0.0001, n = 31), but estimates produced by the 95% UD kernel were 44–243% higher from the ones generated by the MCP. For approximately 48% of the individuals used in the present study, values of Schoener’s ratio ranged from 1.4 to 2.0, indicating a moderate autocorrelation, which may result in a 5% negative bias in the home range estimates. For seven dolphins, location data showed Schoener’s ratios between 1.0 and 1.4, meaning a possible negative bias of 5–10%. Schoener’s ratio was above 2.0 for the remaining dolphins, indicating that the data were independent. The univariate GLM model was not significant for both estimators (MCP: R 2 = 0.04, F (3,27) = 0.386, P = 0.764; 95% UD: R 2 = 0.04, F (3,27) = 0.365, P = 0.779) and we found no significant effect of sample size or autocorrelation in the estimates of home range size produced by MCP or kernel methods.
Range areas varied in size and location for the 31 dolphins. For most of the dolphins, the kernel method produced multiple centres of activity for both the whole range and core areas (Figs. 5, 6). Estimated overall ranging areas and 50% core areas were generally larger for sub-adults (Table 3), though differences were not statistically significant (MCP: U = 80.0, P = 0.379, n = 30; 95% UD: U = 88.0, P = 0.598, n = 30; 50% UD: U = 90.0, P = 0.659, n = 30). Range sizes obtained for the few dolphins for which sex was known did not reveal any obvious differences, although small sample sizes prevented any statistical analysis (Table 3).

Ranging patterns of four resident dolphins in the main area, estimated by MCP and fixed kernel (overall ranging at 95% UD and core area at 50% UD)

Ranging patterns of two non-resident dolphins in the main area, estimated by MCP and fixed kernel (overall ranging at 95% UD and core area at 50% UD)
Overlap in home ranges between pairs of dolphins, varied from 2.2 to 94.3%, with an average of 39.9%. Spatial overlap was especially evident in the core areas. In all the dolphins studied, the 50% core area encompassed at least one of the two extremes of the channel between the islands of Faial and Pico. Percentage of home range overlap was higher among adult dolphins (45.3%), than among subadult (34.9%) and adult-subadult pairs (29.3%) (H = 89.426, P < 0.0001, df = 2). The overlap in the areas used by female–female pairs (48.4%) was higher than the overlap among female–male (41.6%) or male–male pairs (30.4%), but the sample size was too small for any statistical analysis.
Non-resident dolphins in the main area had larger home ranges than resident individuals, although sample sizes were small and differences were not significant (Table 3). Resident dolphins tended to share larger percentages of their home ranges with other resident animals (42.0%) (Fig. 5). Still, there was a high degree of overlap (21.4%) in the home ranges of resident and non-resident individuals (H = 142.408, P < 0.0001, df = 2) (Fig. 6).
Discussion
Residency and site fidelity in the main area
Despite the large number of dolphins identified in the main area, only a small number of individuals (44 animals) was frequently sighted and showed long-term and year-round site fidelity and could be classified as residents. The remaining 567 individuals, classified as non-resident animals, showed varying patterns of occurrence. A few dolphins sighted once in the main area were observed on several occasions in another group of islands. Data on sighting frequencies and inter-annual resightings suggests the existence of resident groups in secondary areas. The majority of dolphins seen once or twice in the main area, however, were never observed again and may have been just passing through the Azores. Other dolphins were seen a number of times but in non-consecutive years, suggesting there may be temporary emigration to other areas within or outside the archipelago. The classification of each dolphin as resident or non-resident is thus not definitive and requires further study.
The residence pattern found in the Azores—with a mixture of residents, transients, and temporary migrants—is in agreement with findings from coastal areas and seems to be a common trait among populations of bottlenose dolphins (Connor et al. 2000). Perhaps more unusual were the large number of individuals identified during this study and the relatively low encounter and resighting rates reported. These results can be partly explained by the large size of the study site. By extending the survey area far beyond the range of the resident group there was a higher chance of photographing transient dolphins or dolphins from different communities but also a lower probability of resighting the resident individuals. The latter point may also explain the relatively small resident group when compared to coastal communities, despite the much larger study site (Wells et al. 1987; Wilson et al. 1999; Ingram and Rogan 2002). However, the size of the study site alone cannot account for the large number of transient dolphins recognized during the surveys. Large populations characterized by a low number of individual resightings are typical of open water habitats (Defran et al. 1999) but also occur in coastal and inshore areas (Shane 2004). The greater productivity of the waters around the islands, compared to oceanic waters, seems responsible for attracting several cetacean species that use the area as a foraging post or migration stop (Silva et al. 2003). Thus, it seems reasonable to speculate that dolphins that occur in neighbouring oceanic regions may also be drawn to the Azores and use the area as a feeding ground on a temporary basis. The fact that these dolphins do not stay in the area also suggests that there are not enough food resources to sustain a larger population permanently. An alternative explanation could be that non-resident dolphins have different foraging strategies or prey preferences which cannot be sustained in the Azores. There is no information on the diet composition of dolphins in the Azores but analysis of fatty acids present in the blubber of resident and non-resident dolphins showed similar profiles (Walton et al. 2007).
In addition to using the area regularly throughout the year, resident dolphins also exhibited considerable geographic fidelity and were never encountered outside the main area. With a single exception, dolphins classified as residents showed very limited movements and, as expected, possessed smaller home ranges than non-resident animals. Within the resident group, there was considerable overlap in the ranging areas and core areas. Ranging behaviour is thought to partially shape the social structure by limiting the number of potential associates of each animal to those individuals that share similar ranges (Lusseau et al. 2006) and several studies have tried to distinguish different communities by examining the ranging and association patterns of individuals (Urian 2002; Lusseau et al. 2006). At present, however, it is not possible to conclude if the similarity in ranging patterns within the resident group means that these individuals comprise a distinct community (sensu Wells et al. 1987) and the hypothesis that the common ranges simply result from aggregative behaviour as a response of higher prey availability cannot be ruled out. In any case, it is clear that these dolphins do not constitute a closed and isolated unit since they often interacted with animals from outside the group and seemed to share extensive areas of their home ranges with non-resident dolphins.
Movements
The average distance between consecutive sightings of individual dolphins was about 20 km and most of the movements documented were less than 50 km. These restricted movements probably correspond to foraging bouts within the usual range of the individual, and may reflect shifts in prey distribution with tidal currents or due to daily horizontal or vertical migrations, as was documented for spinner dolphins (Stenella longirostris) in Hawaii (Benoit-Bird and Au 2003). Some of the movements recorded may also be related to social activities, such as resting or socializing.
More relevant to the question of how ranging behaviour affects the population structure is the finding that dolphins also performed long-distance movements of more than 100 km between the islands. In fact, there was a substantial degree of movement between the studied areas even though the survey effort in some of the secondary areas was very low and the time interval between surveys may have been insufficient to allow dolphins to move freely between areas. Thus, although long-distance movements here reported are likely underestimated, these results show that dolphins from different groups of islands are not geographically isolated from each other.
The fact that long-distance movements were not restricted to a single sex or age class and that at least in one instance a group of 11 individuals was involved, suggests that these movements are not related to reproductive strategies but to foraging. Dolphins were observed to repeatedly move back and forth between two areas, further suggesting that these movements were ranging rather than dispersal movements. We hypothesize that long-distance movements are related to the low abundance and patchy distribution of prey, forcing dolphins to either move between familiar and already established feeding areas or to venture outside their usual range in search for new feeding areas. Nevertheless, not all the individuals exhibited the same degree of mobility and these longer trips may represent an alternative individual or group foraging strategy. Further work will be needed to investigate if individual movement patterns remain consistent through time.
The long-distance movements here reported are not unexpected because individuals of this species are capable of much wider movements. Radiotracking data showed that dolphins are able to travel as much as 55 km in 12 h (Lynn 1995) and in the Southern California Bight (USA) dolphins showed high mobility, ranging up to 470 km along a 1-km stretch of open coastline (Defran et al. 1999). However, moving between oceanic islands is considerably different from moving along the coast, as it involves crossing large areas of deep, open waters, and could therefore entail greater risks to the animals. In Hawaii, bottlenose dolphins and pantropical spotted dolphins (S. attenuata) were found to be island-associated, spending most of their time in shallow waters and showing little or no movement between the islands (Baird et al. 2001, 2002). Similarly, spinner dolphins occurring in a remote Hawaiian atoll lived in stable societies and showed strong site fidelity and restricted movements (Karczmarski et al. 2005). These authors proposed that this type of behaviour may have evolved as a strategy to avoid predation. In this area, available habitats are separated by large geographic distances, forcing dolphins to travel across large stretches of deep open pelagic waters with potentially high risk of shark predation. Increased use of the shallow waters of the Sarasota area by bottlenose dolphins during the peak abundance of bull sharks (Carcharhinus leucas) may also be an adaptation for reducing the risk of predation by reducing the volume of water that must be kept under surveillance (Wells et al. 1980). It could be hypothesized that the higher mobility of dolphins in the Azores and the observed differences to studies conducted in Hawaii (Baird et al. 2001, 2002; Karczmarski et al. 2005) result from a lower abundance and greater temporal variability of food resources and a comparatively lower predation risk. The rarity of the most common dolphin predators and the lack of observations of animals bearing shark bite scars strongly suggest that the risk of predation is small in the Azores. However, the available information cannot be used to directly test any of these hypotheses. In the future, we will need to collect data on the distribution and abundance of potential prey and predator species and to relate those with dolphin movement patterns.
Performance of the home range estimators
Significant differences were found between estimates obtained by the MCP and fixed kernel methods, with the latter producing significantly larger home range sizes. This result is not unexpected as it was found that, at small sample sizes, kernel methods tend to overestimate home range sizes (Seaman et al. 1999), whereas MCP significantly underestimate them (Urian 2002). It has been recommended that home range estimates should be based on a minimum of 30 observations and preferably more than 50 (Seaman et al. 1999). However, Urian (2002) showed that approximately 150 sightings were necessary for obtaining accurate estimates of home ranges of the bottlenose dolphin population in Sarasota. This author also demonstrated that at more than 100 sightings MCP and kernel estimators produced similar results.
In the present study a minimum of 10 sightings was considered an adequate sample size, based on the asymptote of the area-observation curve. Yet, for an area-observation curve to be valid, each individual must display a constant centre of activity throughout the studied period (Gaustestad and Mysterud 1995). Although temporal changes in home range size and location could not be examined due to the small sample size, ranging patterns of individual animals are likely to have varied throughout the 6-year study period. Hence, estimates of home range size presented here could be affected by small sample sizes. Specifically, in the comparison of home range size between resident and non-resident dolphins, we would expect MCP estimates of home range size of non-resident animals to be underestimated, while kernel estimates may be overestimated. In addition, location data for 22 dolphins showed a moderate degree of autocorrelation, which could result in a 5–10% negative bias of home range size (Swihart and Slade 1997). Although this level of bias is negligible, in the case of the kernel method, bias arising from autocorrelated data would act in the opposite direction to the one caused by small sample sizes, while for MCP estimates the negative effect of both biases would add up. In spite of this, we found no significant effect of sample size and autocorrelation on the estimates of home range size obtained with each method. Thus, it is impossible to quantify the magnitude of the overall bias and to assess its effect in the results, and the estimates of home range size here provided should be viewed as approximations of actual ranges.
Home range characteristics
There was considerable overlap in the overall ranging area and core area of the 31 dolphins, independent of the sex or age class of the individuals. Even though the area regularly surveyed extended up to 8 km from the islands, the results presented here showed a clear pattern of preferential use for the areas very close to the islands. These results are entirely supported by the analysis of habitat preferences of bottlenose dolphins in the Azores using a different dataset (Silva 2007). According to this study, bottlenose dolphins preferentially used shallow areas (between 100 and 600 m) with high bottom relief. In the Azores, the absence of a continental shelf limits this kind of physiography to a narrow stretch around the islands. The areas close to the islands may provide a more suitable habitat compared to open waters. First, because the islands are responsible for creating small-scale upwellings and for trapping flow-driven nutrients that generate enhanced biological productivity and ultimately serve to concentrate food resources for the dolphins. Second, because by dwelling in shallower areas close to the islands the dolphins may also take advantage of bottom fishes in addition to schooling prey.
Examination of the 50% core area of these dolphins uncovered the location of two critical areas for the population within the study site. For all dolphins examined at least one of the entrances of the channel between the islands of Faial and Pico was part of their core area, and some individuals showed two separate core areas, located at both extremes of the channel. Being approximately 5–8 km wide and 12 km long, the channel is characterized by comparatively shallower waters (<200 m), irregular topography (depth varies from 8 to 200 m), strong tidal currents and a diversity of seabed types (Tempera et al. 2001). These features account for the significantly higher productivity of the channel relative to the surrounding areas, and are largely responsible for the observed diversity of habitats and species (Tempera et al. 2001). High usage of areas characterized by strong tidal currents and irregular bottom topography has been widely documented for several dolphin species (Norris and Dohl 1980). It has been suggested that these features may lead to increased foraging efficiency, either because the higher productivity or because the accumulation of fish in the area results in greater prey densities, or through facilitating prey capture (Norris and Dohl 1980; Wilson et al. 1997).
Dolphins in the Azores maintain large home ranges
Notwithstanding the scarcity of information available and the difficulty of comparing estimates obtained with different methods and/or sample sizes, estimates of home range size for bottlenose dolphins in the Azores (MCP: 62.9–721.1 km2; 95% UD: 171.4–1887.2 km2; 50% UD: 30.0–417.8 km2) were found to be 2–3 times greater than those previously reported for this species. MCP areas of dolphins sighted in the Shannon Estuary (Ireland) varied between 19.2 and 75.5 km2, with a mean of 47.7 km2 (Ingram and Rogan 2002). In Sarasota, Owen et al. (2002) estimated a mean 95% UD of 162.6 km2 and 72.1 km2, and a mean 50% UD of 28.7 km2 and 16.6 km2, for paired and unpaired males, respectively. Home range size (95% UD) calculated with the adaptive kernel of dolphins in South Carolina ranged from 17.2 to 98.9 km2 (mean = 51.3 km2). Using the MCP estimator, mean home range size of these dolphins was 40.8 km2 (Gubbins 2002). Although larger than previous estimates, mean area range (mean = 140.0 km2, SD = 90.7 km2) recorded for ten dolphins radiotracked in Matagorda Bay, Texas (Lynn 1995) is still substantially smaller than the estimates presented in this study.
The differences between the home range estimates in this study and others cannot be entirely explained by methodological distinctions. In the present study, the area regularly surveyed was over 5,400 km2 and a few surveys were conducted outside this area. Unless the study site encompasses the whole area generally used by the individuals, estimates of home range size will be negatively biased and will fail to represent actual home ranges. This seemed to be the case of the home ranges calculated for dolphins in the Shannon estuary, but does not explain the difference to the other sites.
As previously noted, patchy resources and/or lower habitat productivity are known to cause an increase in the overall ranging area as well as in the core area, as the intensity of use throughout the area becomes even (Ford 1983). Oceanic islands are themselves generators of biological patchiness (Barton et al. 2000), which coupled with the highly dynamic oceanic ecosystem of the Azores and the lower predation rate, in contrast to the generally more productive coastal and inshore study sites mentioned earlier, could explain the much larger home ranges documented in this study, as well as the extensive movements reported.
Dolphins from different sexes and age classes show similar ranging patterns
Female space use patterns are usually considered to be more directly affected by ecological parameters, such as availability of resources and predation risk. Male range patterns are primarily driven by female distribution because a male’s reproductive success depends on the number of mates he’s able to find and defend (Clutton-Brock 1989). Thus, in several mammalian species, males have larger home ranges than females, because females tend to minimize their ranges for reasons of energetic efficiency, whereas for males it is advantageous to range widely in search for potential mates (Sandell 1989). In species with pronounced sexual dimorphism, males may have larger home range sizes in response to their higher energetic demands as a consequence of their larger body size (Sandell 1989). Among coastal populations of bottlenose dolphins, males also seem to have much larger home ranges than females (Wells et al. 1987; Krützen et al. 2004). In the present study, however, ranging and core areas and the frequency distribution of distances travelled were similar in male and female dolphins. We suggest the lack of sexual differences in bottlenose dolphin’s home range size may be related to the lower productivity of this ecosystem compared to the coastal areas. Environmental productivity has been suggested as the main factor responsible for lack of differences in home range sizes among sexes in several marsupials (Fisher and Owens 2000). In the Azores, females need to maintain much larger home ranges to meet their energetic requirements. Consequently, males would have to increase their ranges much beyond the areas used by females, in order to maximize their contacts with many receptive females. Such increase in home range size may be to a point that it turns out to be energetically unfeasible for males. Also, the high degree of overlap in female home ranges and the absence of habitat partitioning among different groups of dolphins makes it unnecessary for males to range further in order to gain access to potential mates.
Other factors being equal, adults should have significantly larger home ranges due to their higher energetic demands (Sandell 1989). It is not clear why the same pattern was not observed in this study, especially because the less productive ecosystem of the Azores should make more obvious the tendency towards larger home ranges in response to the energetic constraints of the individuals (e.g. Herfindal et al. 2005). Typically, however, home range size is not determined by a single factor but results from the combination of several variables working simultaneously (McLoughlin and Ferguson 2000). Thus, it is possible that other factors also known to influence ranging patterns and not accounted for in this study (e.g. social affiliations and social learning of foraging techniques) could be obscuring the effect of body size.
Implications for population structure
In the Azores, the geographic distance between groups of islands is within the range ability of bottlenose dolphins and would hardly be an important factor in preventing interchange between areas. Furthermore, there is no evidence of any topographic feature or oceanographic phenomenon that could provide an ecological barrier to gene flow. Still, considering that dolphins would have to cross large areas of deep, open waters to move between different groups of islands, it would be reasonable to expect that the population would be divided into distinct geographic communities associated with each group of islands, with little or no movement between areas. The findings concerning residence and ranging patterns contradict this expectation and have strong implications regarding the structure of the population. The extensive ranging behaviour exhibited by some dolphins and the apparent lack of territoriality provide an opportunity for animals associated with different islands to mix. In addition, the islands are regularly visited by hundreds of dolphins that are not residents in any single group of islands. Whether residents at other islands or transients, dolphins from outside the main area interacted socially with resident animals. Although it is impossible to determine if breeding actually takes place during these encounters, it is evident that dolphins from different groups of islands are not isolated and genetic interchange is likely to occur, thus preventing genetic divergence of geographic-based communities.
Analysis of mitochondrial DNA and microsatellite DNA markers supports this hypothesis, indicating a lack of genetic differentiation between dolphins sampled in different groups of islands and the total absence of a population structure within the Azores (Quérouil et al. 2007). Fatty acid analysis carried out using the same samples used for genetic analyses also failed to reveal any clear distinctions between dolphins from different groups of islands (Walton et al. 2007). In addition, lack of isolation of the resident group is also supported by results of genetic analyses. Genetic diversity at the level of mtDNA shown by resident dolphins was similar to that found for the whole Azorean sample and microsatellites indicated that mean relatedness within the resident group was similar to that of the whole Azorean sample (Quérouil et al. 2007).
Conclusions
The high mobility of individuals and the varying patterns of residence in any single area suggest that bottlenose dolphins in the Azores constitute a single and open population, composed of several geographic communities that maintain social interactions with neighbouring communities and groups from within and outside the archipelago. These interactions are facilitated by the extensive ranging behaviour of some individuals and groups and by an apparent lack of habitat partitioning. This work also provides support for the hypothesis that dolphins in open waters carry out extensive movements and have large home ranges in response to the lower density and patchy distribution of food resources. Apart from the greater mobility shown by some individuals, the population of bottlenose dolphins living around the islands of the Azores shares several behavioural characteristics with resident populations in coastal habitats.
References
Baird RW, Ligon AD, Hooker SK, Gorgone AM (2001) Subsurface and nighttime behaviour of pantropical spotted dolphins in Hawai’i. Can J Zool 79:988–996. doi:https://doi.org/10.1139/cjz-79-6-988
Baird RW, Gorgone AM, Webster DL (2002) An examination of movements of bottlenose dolphins between islands in the Hawaiian island chain. Report prepared under contract #40JGNF110270 to the Southwest Fisheries Science Center, National Marine Fisheries Service, La Jolla
Ballance LT (1992) Habitat use patterns and ranges of the bottlenose dolphin in the Gulf of California, Mexico. Mar Mamm Sci 8:262–274. doi:https://doi.org/10.1111/j.1748-7692.1992.tb00408.x
Barton ED, Basterretxea G, Flament P, Mitchelson E, Jacob E, Jones B, Arlistegui J, Herrera F (2000) Lee region of Gran Canaria. J Geophys Res C 105:17173–17193. doi:https://doi.org/10.1029/2000JC900010
Benoit-Bird KJ, Au WWL (2003) Prey dynamics affect foraging by a pelagic predator (Stenella longirostris) over a range of spatial and temporal scales. Behav Ecol Sociobiol 53:364–373
Burt WH (1943) Territoriality and home range concepts as applied to mammals. J Mammal 30:25–27. doi:https://doi.org/10.2307/1375192
Caldeira RMA, Groom S, Miller P, Pilgrim D, Nezlin NP (2002) Sea-surface signatures of the island mass effect phenomena around Madeira Island, Northeast Atlantic. Remote Sens Environ 80:336–360. doi:https://doi.org/10.1016/S0034-4257(01)00316-9
Clutton-Brock TH (1989) Mammalian mating systems. Proc R Soc Lond B Biol Sci 236:1–39
Connor RC, Wells RS, Mann J, Read AJ (2000) The bottlenose dolphin: social relationships in a fission-fusion society. In: Mann J, Connor RC, Tyack PL, Whitehead H (eds) Cetacean societies: field studies of dolphins and whales. The University of Chicago Press, Chicago and London, pp 91–126
Damuth J (1981) Home range, home range overlap, and species energy use among herbivorous mammals. Biol J Linn Soc Lond 15:185–193. doi:https://doi.org/10.1111/j.1095-8312.1981.tb00758.x
Defran RH, Weller DW, Kelly DL, Espinosa MA (1999) Range characteristics of Pacific coast bottlenose dolphins (Tursiops truncatus) in the Southern California Bight. Mar Mamm Sci 15:381–393. doi:https://doi.org/10.1111/j.1748-7692.1999.tb00808.x
Fisher DO, Owens IPF (2000) Female home range size and the evolution of social organization in macropods marsupials. J Anim Ecol 69:1083–1098. doi:https://doi.org/10.1046/j.1365-2656.2000.00450.x
Ford RG (1983) Home range in a patchy environment: optimal foraging predictions. Am Zool 23:315–326
Gaustestad AO, Mysterud I (1995) The home range ghost. Oikos 74:195–204. doi:https://doi.org/10.2307/3545648
Genin A (2004) Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies. J Mar Syst 50:3–20. doi:https://doi.org/10.1016/j.jmarsys.2003.10.008
Gubbins C (2002) Use of home ranges by resident bottlenose dolphins (Tursiops truncatus) in a South Carolina estuary. Aquat Mamm 83:178–187
Herfindal I, Linnell JDC, Odden J, Birkeland Nilsen E, Andersen R (2005) Prey density, environmental productivity and home-range size in the Eurasian lynx (Lynx lynx). J Zool (Lond) 265:63–71. doi:https://doi.org/10.1017/S0952836904006053
Hooge PN, Eichenlaub B (1997) Animal movement extension to arcview. ver. 1.1. Alaska Science Center—Biological Science Office, U.S. Geological Survey, Anchorage
Ingram SN, Rogan E (2002) Identifying critical areas and habitat preferences of bottlenose dolphins Tursiops truncatus. Mar Ecol Prog Ser 244:247–255. doi:https://doi.org/10.3354/meps244247
Irvine AB, Scott MD, Wells RS, Kaufmann JH (1981) Movements and activities of the Atlantic bottlenose dolphin, Tursiops truncatus, near Sarasota, Florida. Fish Bull (Wash D C) 79:671–688
Karczmarski L, Würsig B, Gailey G, Larson KW, Vanderlip C (2005) Spinner dolphins in a remote Hawaiian atoll: social grouping and population structure. Behav Ecol 16:675–685. doi:https://doi.org/10.1093/beheco/ari028
Kernohan BJ, Gitzen RA, Millspaugh JJ (2001) Analysis of animal space use and movements. In: Millspaugh JJ, Marzluff JM (eds) Radio tracking and animal populations. Academic Press, Sand Diego, pp 125–166
Krützen M, Sherwin WB, Berggren P, Gales N (2004) Population structure in an inshore cetacean revealed by microsatellite and mtDNA analysis: bottlenose dolphins (Tursiops sp.) in Shark Bay, Western Australia. Mar Mamm Sci 20:28–47. doi:https://doi.org/10.1111/j.1748-7692.2004.tb01139.x
Lusseau D, Wilson B, Hammond PS, Grellier K, Durban JW, Parsons KM, Barton TR, Thompson PM (2006) Quantifying the influence of sociality on population structure in bottlenose dolphins. J Anim Ecol 75:14–24. doi:https://doi.org/10.1111/j.1365-2656.2005.01013.x
Lynn SK (1995) Movements, site fidelity, and surfacing patterns of bottlenose dolphins on the central Texas coast. MS Thesis, Texas A&M University
Mann J, Smuts B (1999) Behavioral development in wild bottlenose dolphin newborns (Tursiops sp.). Behaviour 136:529–566. doi:https://doi.org/10.1163/156853999501469
McLoughlin PD, Ferguson SH (2000) A hierarchical pattern of limiting factor helps explain variation in home range size. Ecoscience 7:123–130
McNab BK (1963) Bioenergetics and the determination of home range size. Am Nat 894:133–140. doi:https://doi.org/10.1086/282264
Mohr CO (1947) Table of equivalent populations of North American small mammals. Am Midl Nat 37:223–249. doi:https://doi.org/10.2307/2421652
Norris KS, Dohl TP (1980) The structure and functions of cetacean schools. In: Herman LM (ed) Cetacean behaviour: mechanisms and function. Wiley, USA, pp 211–261
Owen ECG, Wells RS, Hofmann S (2002) Ranging and association patterns of paired and unpaired adult male Atlantic bottlenose dolphins, Tursiops truncatus, in Sarasota, Florida, provide no evidence for alternative male strategies. Can J Zool 80:2072–2089. doi:https://doi.org/10.1139/z02-195
Palacios DM (2002) Factors influencing the island-mass effect of the Galápagos Archipelago. Geophys Res Lett 29:2134–2138. doi:https://doi.org/10.1029/2002GL016232
Quérouil S, Silva MA, Freitas L, Prieto R, Magalhães S, Dinis A, Alves F, Matos JA, Mendonça D, Hammond P, Santos RS (2007) High gene flow in oceanic bottlenose dolphins (Tursiops truncatus) of the North Atlantic. Conserv Genet 8:1405–1419. doi:https://doi.org/10.1007/s10592-007-9291-5
Sandell M (1989) The mating tactics and spacing behaviour of solitary carnivores. In: Gittleman JL (ed) Carnivore behaviour, ecology and evolution. New York, Cornell University Press, pp 164–182
Santos RS, Hawkins S, Monteiro LR, Alves M, Isidro E (1995) Marine research, resources and conservation in the Azores. Aquat Conserv Mar Freshw Ecosyst 5:311–354. doi:https://doi.org/10.1002/aqc.3270050406
Schoener TW (1981) An empirically based estimate of home range. Theor Popul Biol 20:281–325. doi:https://doi.org/10.1016/0040-5809(81)90049-6
Scott MD, Wells RS, Irvine AB (1990) A long-term study of bottlenose dolphins on the west coast of Florida. In: Leatherwood S, Reeves RR (eds) The bottlenose dolphin. Academic Press, San Diego, pp 235–244
Seaman DE, Millspaugh JJ, Kernohan BJ, Brundige GC, Raedeke KJ, Gitzen RA (1999) Effects of sample size on kernel home range estimates. J Wildl Manage 63:739–747. doi:https://doi.org/10.2307/3802664
Shane SH (2004) Residence patterns, group characteristics, and association patterns of bottlenose dolphins near Sanibel Island, Florida. Gulf Mex Sci 1:1–12
Silva MA (2007) Population biology of bottlenose dolphins in the Azores Archipelago. PhD thesis, University of St Andrews
Silva MA, Prieto R, Magalhães S, Cabecinhas R, Cruz A, Gonçalves JM, Santos RS (2003) Occurrence and distribution of cetaceans in the waters around the Azores (Portugal), Summer and Autumn 1999–2000. Aquat Mamm 29:77–83. doi:https://doi.org/10.1578/016754203101024095
Stevick PT, McConnell BJ, Hammond PS (2002) Patterns of movement. In: Hoelzel AR (ed) Marine mammal biology: an evolutionary approach. Blackwell Publishing, Oxford, pp 185–216
Swihart RK, Slade NA (1997) On testing for independence of animal movements. J Agric Biol Environ Stat 2:48–63. doi:https://doi.org/10.2307/1400640
Swihart RK, Slade NA, Bergstrom BJ (1988) Relating body size to the rate of home range use in mammals. Ecology 69:393–399. doi:https://doi.org/10.2307/1940437
Tempera F, Afonso P, Morato T, Prieto R, Silva M, Cruz A, Gonçalves J, Santos RS (2001) Comunidades biológicas marinhas dos Sítios de Interesse Comunitário do Canal Faial-Pico. Arquivos do DOP. Ser Relatorio Internos 5:1–95
Urian KW (2002) Community structure of bottlenose dolphins (Tursiops truncatus) in Tampa Bay, Florida, USA. MS thesis, University of North Carolina
Walton MJ, Silva MA, Magalhães S, Prieto R, Santos RS (2007) Using blubber biopsies to provide ecological information about bottlenose dolphins Tursiops truncatus around the Azores. J Mar Biol Assoc U K 87:223–230. doi:https://doi.org/10.1017/S0025315407054537
Wells RS, Irvine AB, Scott MD (1980) The social ecology of inshore odontocetes. In: Herman LM (ed) Cetacean behaviour: mechanisms and function. Wiley, New York
Wells RS, Scott MD, Irvine AB (1987) The social structure of free-ranging bottlenose dolphins. In: Genoways H (ed) Current mammalogy. Plenum Press, New York, pp 247–305
White GC, Garrott RA (eds) (1990) Analysis of wildlife radio-tracking data. Academic Press, San Diego
Wiens JA (1976) Population responses to patchy environments. Annu Rev Ecol Syst 7:81–120. doi:https://doi.org/10.1146/annurev.es.07.110176.000501
Wilson B, Thompson PM, Hammond PS (1997) Habitat use by bottlenose dolphins: seasonal distribution and stratified movement patterns in the Moray Firth, Scotland. J Appl Ecol 34:1365–1374. doi:https://doi.org/10.2307/2405254
Wilson B, Hammond PS, Thompson PM (1999) Estimating size and assessing trends in a coastal bottlenose dolphin population. Ecol Appl 9:288–300. doi:https://doi.org/10.1890/1051-0761(1999)009[0288:ESAATI]2.0.CO;2
Worton BJ (1989) Kernel methods for estimating the utilization distribution in home-range studies. Ecology 70:164–168. doi:https://doi.org/10.2307/1938423
Würsig B, Jefferson TA (1990) Methods of photo-identification for small cetaceans. Rep Int Whal Comm Spec Issue 12:43–52
Acknowledgments
This research was funded by the Portuguese Science and Technology Foundation (FCT), under the CETAMARH project (POCTI/BSE/38991/01), by an EU-LIFE program (B4-3200/98/509), and by an Interreg program (Interreg IIIBMAC/4.2/A2). We are also grateful to FCT for funding M. A. S. doctoral grant (SFRH/BD/8609/2002) and post-doctoral grants (SFRH/BPD/29841/2006), and S. M. M. and M. I. S. research grants through the CETAMARH project. We are thankful to the skippers (Filipe Goulart, P. Martins, V. Rosa, R. Bettencourt and N. Serpa), crew members, students and volunteers that helped us throughout this study. We also thank David Lusseau and three anonymous reviewers for their comments on an early draft of the manuscript. IMAR-DOP/UAç is the R&D Unit #531 funded through the pluri-annual and programmatic funding schemes of FCT-MCTES and DRCT-Azores. This research was conducted under license of the Environment Directorate of the Regional Government of the Azores.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Lewison.
Rights and permissions
About this article
Cite this article
Silva, M.A., Prieto, R., Magalhães, S. et al. Ranging patterns of bottlenose dolphins living in oceanic waters: implications for population structure. Mar Biol 156, 179–192 (2008). https://doi.org/10.1007/s00227-008-1075-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-008-1075-z